
The Effects of Shiga Toxin 1, 2 and Their Subunits on Cytokine and Chemokine Expression by Human Macrophage-Like THP-1 Cells

Abstract
:1. Introduction
2. Results
2.1. Effects of Stx1 and Stx2 Exposure on Cytokine/Chemokine Expression by Macrophage-Like THP-1 Cells

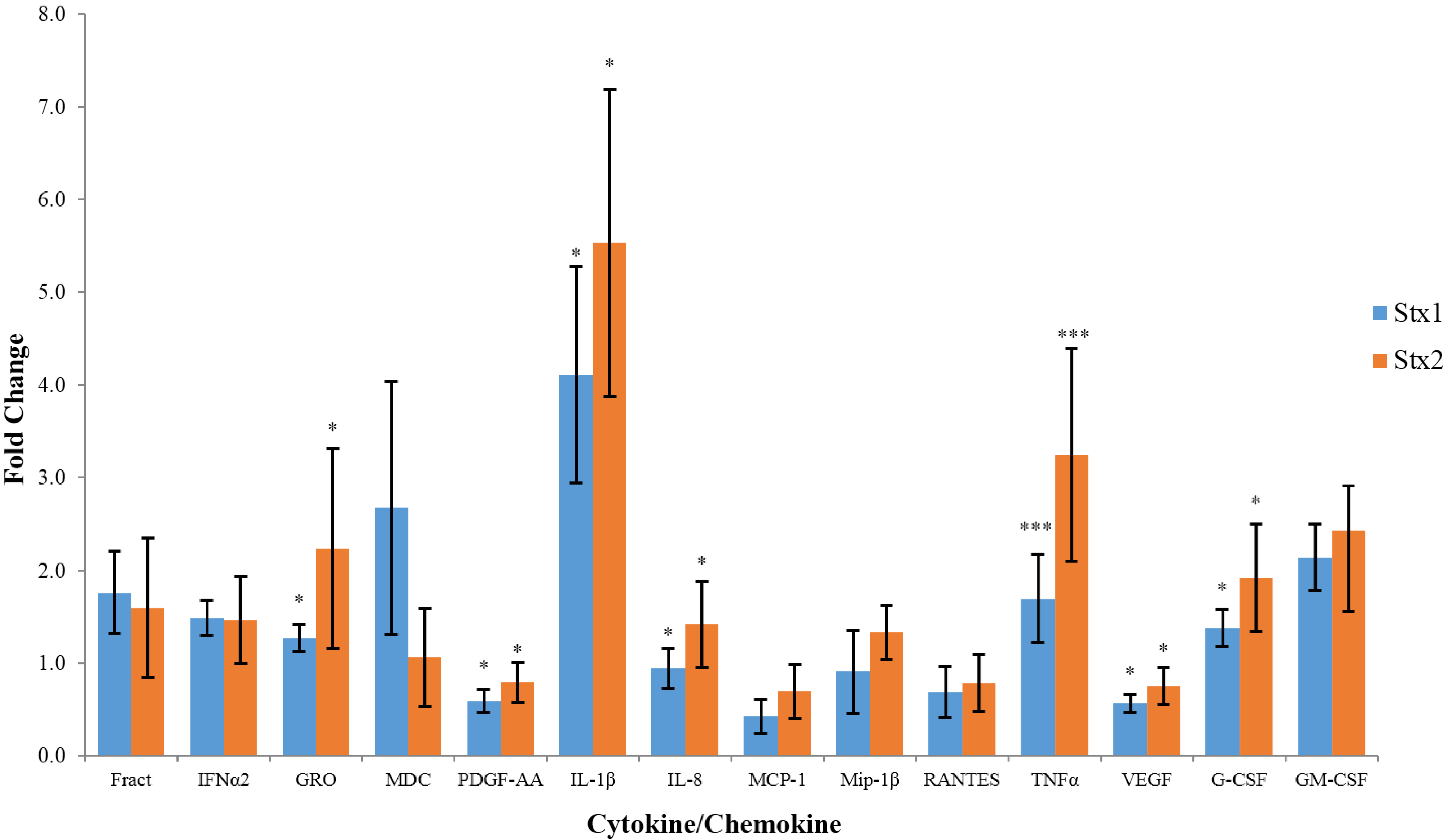{kind=link}
{kind=link}
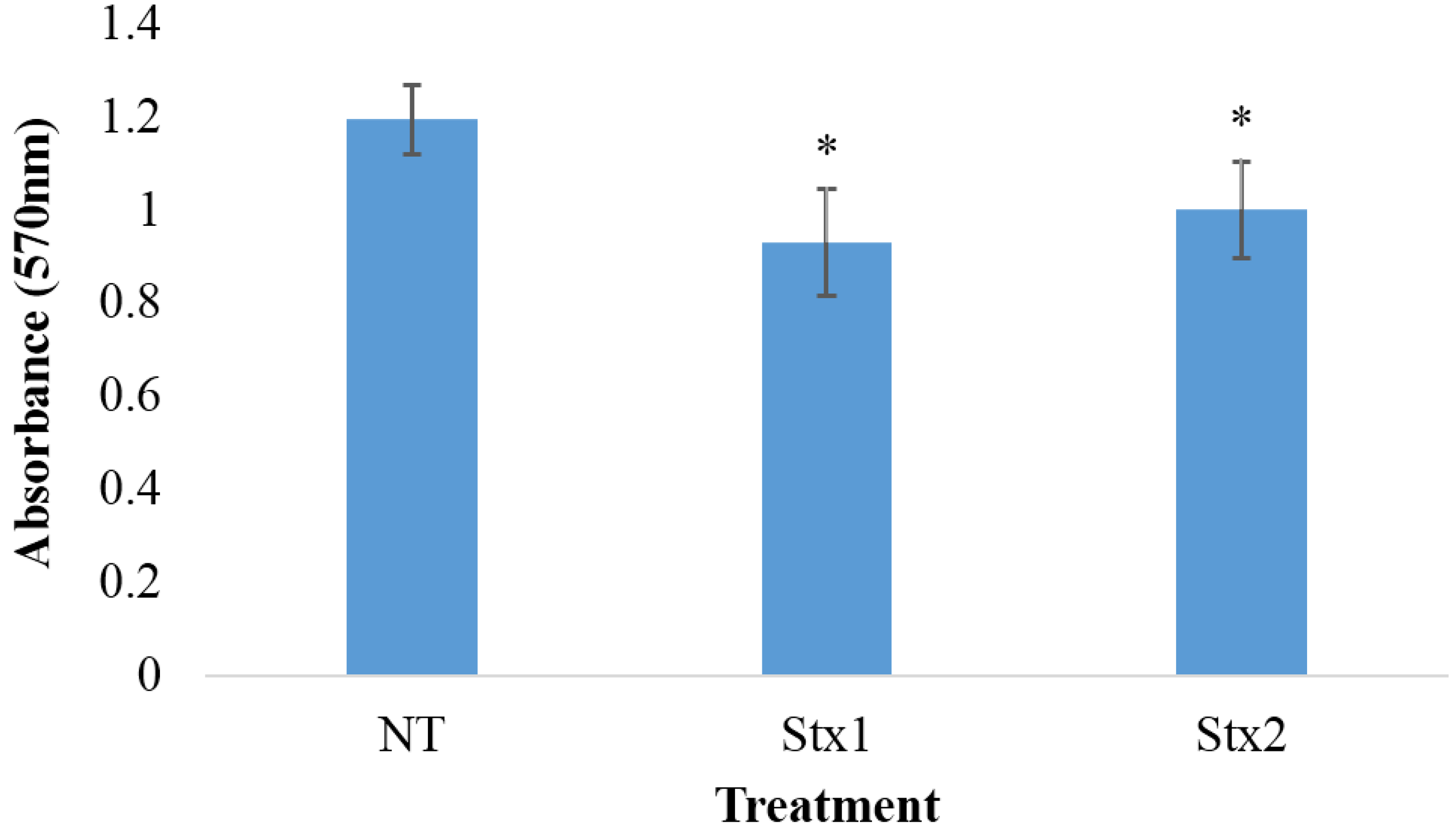{kind=link}
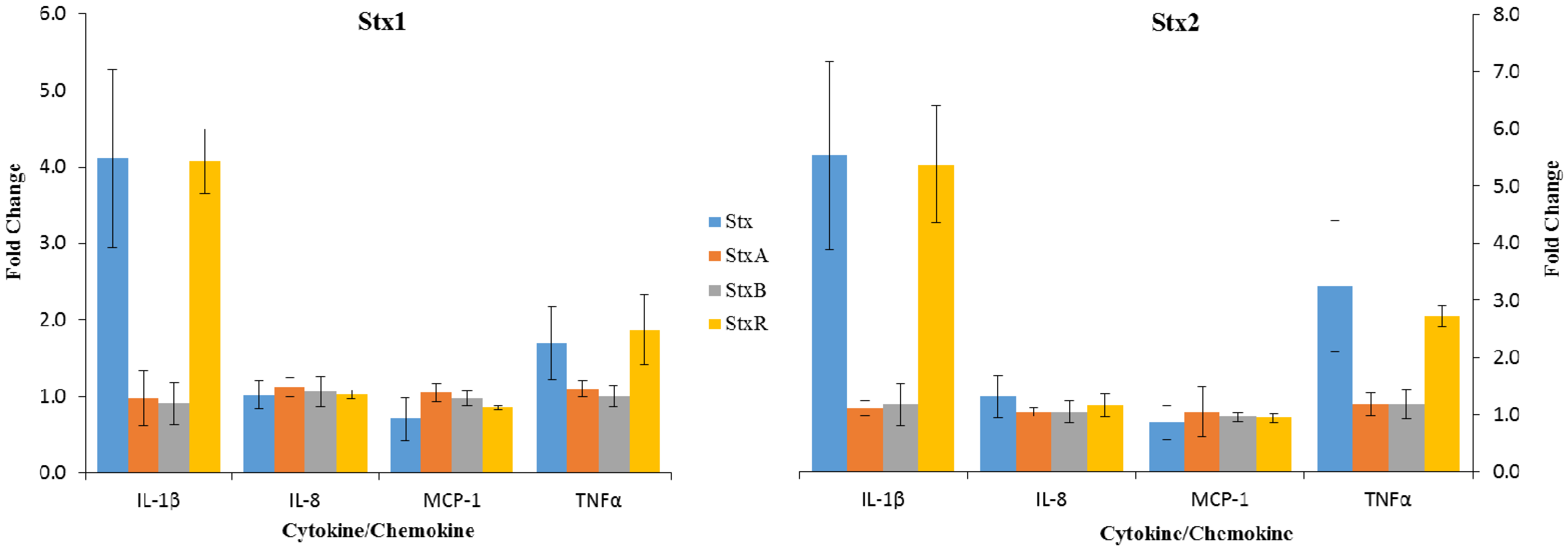{kind=link}
| Analytes | ||||||
|---|---|---|---|---|---|---|
| EGF | Eotaxin-1 | FGF-2 | Flt-3L | Fractalkine 1 | G-CSF | GM-CSF |
| GRO | IFNα2 | IFNγ | IL-1α | IL-1β | IL-1rα | IL-2 |
| IL-3 | IL-4 | IL-5 | IL-6 | IL-7 | IL-8 | IL-9 |
| IL-10 | IL-12(p40) | IL-12(p70) | IL-13 | IL-15 | IL-17A | IP-10 |
| MCP-1 | MCP-3 | MDC | Mip-1β | PDGF-AA | PDGF-AB/BB | RANTES |
| sCD40L | TGF-α | TNFα | TNFβ | VEGF-A | ||

| Analytes | |||||
|---|---|---|---|---|---|
| GM-CSF 1 | IFNγ | IL-1β | IL-2 | IL-4 | IL-6 |
| IL-8 | IL-10 | IL-12(p70) | MCP-1 | TNFα | |

2.2. Effects of Stx1 and Stx2 Exposure on the Viability of Macrophage-Like THP-1 Cells

2.3. Effects of StxA and StxB Subunits on Cytokine/Chemokine Expression by Macrophage-Like THP-1 Cells

3. Discussion
4. Experimental Section
4.1. THP-1 Cell Culture and Differentiation
4.2. Shiga Toxins
4.3. Macrophage-Like THP-1 Cell Cytokine/Chemokine Analysis
4.4. MTT Assay
4.5. Shiga Toxin Subunit Separation and Evaluation
4.6. Macrophage-Like THP-1 Cell Cytokine/Chemokine Analysis for the Effects of Shiga Toxin A and B Subunits
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jandhyala, D.M.; Ahluwalia, A.; Obrig, T.G.; Thorpe, C.M. ZAK: A MAP3Kinase that transduces Shiga toxin-and ricin-induced proinflammatory cytokine expression. Cell. Microbiol. 2008, 10, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.S.; Cleary, T.G.; Obrig, T.G. Recent advances in understanding the pathogenesis of the hemolytic uremic syndrome. Pediatr. Nephrol. 1990, 4, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Y.; Sperandio, V. Enterohemorrhagic E. coli (EHEC) pathogenesis. Front. Cell. Infect. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.D.; Holmes, R.K. Shiga and Shiga-like toxins. Microbiol. Rev. 1987, 51, 206–220. [Google Scholar] [PubMed]
- Zoja, C.; Buelli, S.; Morigi, M. Shiga toxin-associated hemolytic uremic syndrome: Pathophysiology of endothelial dysfunction. Pediatr. Nephrol. 2010, 25, 2231–2240. [Google Scholar] [CrossRef] [PubMed]
- Boerlin, P.; McEwen, S.A.; Boerlin-Petzold, F.; Wilson, J.B.; Johnson, R.P.; Gyles, C.L. Associations between virulence factors of Shiga toxin-producing Escherichia coli and disease in humans. J. Clin. Microbiol. 1999, 37, 497–503. [Google Scholar] [PubMed]
- Fraser, M.E.; Chernaia, M.M.; Kozlov, Y.V.; James, M.N.G. Crystal structure of the holotoxin from Shigella dysenteriae at 2.5 Å resolution. Nat. Struct. Biol. 1994, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, C.A.; Binnington, B.; Manis, A.; Branch, D.R. Globotriaosyl ceramide receptor function—Where membrane structure and pathology intersect. FEBS Lett. 2010, 584, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Garred, Ø.; Prydz, K.; Kozlov, J.V.; Hansen, S.H.; van Deurs, B. Retrograde transport of endocytosed Shiga toxin to the endoplasmic reticulum. Nature 1992, 358, 510–512. [Google Scholar] [CrossRef] [PubMed]
- Kurmanova, A.; Llorente, A.; Polesskaya, A.; Garred, Ø.; Olsnes, S.; Kozlov, J.; Sandvig, K. Structural requirements for furin-induced cleavage and activation of Shiga toxin. Biochem. Biophys. Res. Commun. 2007, 357, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Ogasawara, T.; Ito, K.; Yutsudo, T.; Takeda, Y. Inhibition of elongation factor 1-dependent aminoacyl-tRNA binding to ribosomes by Shiga-like toxin I (VT1) from Escherichia coli O157:H7 and by Shiga toxin. FEMS Microbiol. Lett. 1987, 44, 91–94. [Google Scholar] [CrossRef]
- Westerholt, S.; Pieper, A.K.; Griebel, M.; Volk, H.D.; Hartung, T.; Oberhoffer, R. Characterization of the cytokine immune response in children who have experienced an episode of typical hemolytic-uremic syndrome. Clin. Diagn. Lab. Immunol. 2003, 10, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Litalien, C.; Proulx, F.; Mariscalco, M.M.; Robitaille, P.; Turgeon, J.P.; Orrbine, E.; Rowe, P.C.; McLaine, P.N.; Seidman, E. Circulating inflammatory cytokine levels in hemolytic uremic syndrome. Pediatr. Nephrol. 1999, 13, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Van der Kar, N.C.; Monnens, L.A.; Karmali, M.A.; van Hinsbergh, V.W. Tumour necrosis factor and interleukin-1 induce expression of the verocytotoxin receptor globotriaosylceramide on human endothelial cells: Implications for the pathogensis of the haemolytic uremic syndrome. Blood 1992, 80, 2755–2764. [Google Scholar]
- Proulx, F.; Toledano, B.; Phan, V.; Clermont, M.J.; Mariscalco, M.M.; Seidman, E.G. Circulating Granulocyte Colony-Stimulating Factor, C–X–C, and C–C Chemokines in Children with Escherichia Coli O157:H7 Associated Hemolytic Uremic Syndrome. Pediatr. Res. 2002, 52, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Keepers, T.R.; Gross, L.K.; Obrig, T.G. Monocyte Chemoattractant Protein 1, Macrophage Inflammatory Protein 1, and CCL5 Recruit Macrophages to the Kidney in a Mouse Model of Hemolytic-Uremic Syndrome. Infect. Immun. 2007, 75, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Harrison, L.M.; van Haaften, W.C.E.; Tesh, V.L. Regulation of Proinflammatory Cytokine Expression by Shiga Toxin 1 and/or Lipopolysaccharides in the Human Monocytic Cell Line THP-1. Infect. Immun. 2004, 72, 2618–2627. [Google Scholar] [CrossRef] [PubMed]
- Levya-Illades, D.; Cherla, R.P.; Lee, M.S.; Tesh, V.L. Regulation of cytokine and chemokine expression by the ribotoxic stress response elicited by Shiga toxin type 1 in human macrophage-like THP-1 cells. Infect. Immun. 2012, 80, 2109–2120. [Google Scholar] [CrossRef] [PubMed]
- Van Setten, P.A.; van Hinsbergh, V.W.; van den Heuvel, L.P.; Prefers, F.; Dijkman, H.B.; van der Velden, T.J.; Monnens, L.A. Monocyte Chemoattractant Protein-1 and Interleukin-8 Levels in Urine and Serum of Patients with Hemolytic Uremic Syndrome. Pediatr. Res. 1998, 43, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth, S.R.; Tipping, P.G. Macrophage-induced glomerular fibrin deposition in experimental glomerulonephritis in the rabbit. J. Clin. Investig. 1985, 76, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Karpman, D.; Andreasson, A.; Thysell, H.; Kaplan, B.S.; Svanborg, C. Cytokines in childhood hemolytic uremic syndrome and thrombotic thrombocytopenic purpura. Pediatr. Nephrol. 1995, 9, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Sean, P.; Roth, R.J.; Wu, J.J.; Jones, E.A.; Bennett, A.M.; Flavell, R.A. Dynamic regulation of pro- and anti-inflammatory cytokines by MAPK phosphatase 1 (MKP-1) in innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 2274–2279. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Tani, S.; Matsumoto, Y.; Takeda, T. Serum amyloid P component is the Shiga toxin 2-neutralizing factor in human blood. J. Biol. Chem. 2001, 276, 41576–41579. [Google Scholar] [CrossRef] [PubMed]
- Griener, T.P.; Strecker, J.G.; Humphries, R.M.; Mulvey, G.L.; Fuentealba, C.; Hancock, R.E.W.; Armstrong, G.D. Lipopolysaccharide renders transgenic mice expressing human serum amyloid P component sensitive to Shiga toxin 2. PLoS ONE 2011, 6, e21457. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, G.; Vanmaele, R.; Mrazek, M.; Cahill, M.; Armstrong, G.D. Affinity purification of Shiga-like toxin I and Shiga-like toxin II. J. Microbiol. Methods 1998, 32, 247–252. [Google Scholar] [CrossRef]
- Mulvey, G.M.; Rafter, D.J.; Armstrong, G.D. Potential for using antibiotics combined with a Shiga toxin-absorbing agent for treating 0157:H7 Escherichia coli infections. Can. J. Chem. 2002, 80, 871–874. [Google Scholar] [CrossRef]
- Head, S.C.; Karmali, M.A.; Lingwood, C.A. Preparation of VT1 and VT2 hybrid toxins from their purified dissociated subunits. Evidence for B subunit modulation of a subunit function. J. Biol. Chem. 1991, 266, 3617–3621. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandelli, J.R.; Griener, T.P.; Laing, A.; Mulvey, G.; Armstrong, G.D. The Effects of Shiga Toxin 1, 2 and Their Subunits on Cytokine and Chemokine Expression by Human Macrophage-Like THP-1 Cells. Toxins 2015, 7, 4054-4066. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7104054
Brandelli JR, Griener TP, Laing A, Mulvey G, Armstrong GD. The Effects of Shiga Toxin 1, 2 and Their Subunits on Cytokine and Chemokine Expression by Human Macrophage-Like THP-1 Cells. Toxins. 2015; 7(10):4054-4066. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7104054
Chicago/Turabian StyleBrandelli, Jeremy R., Thomas P. Griener, Austin Laing, George Mulvey, and Glen D. Armstrong. 2015. "The Effects of Shiga Toxin 1, 2 and Their Subunits on Cytokine and Chemokine Expression by Human Macrophage-Like THP-1 Cells" Toxins 7, no. 10: 4054-4066. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7104054