
Comparative Ochratoxin Toxicity: A Review of the Available Data

Abstract
:1. Ochratoxin A—An Important Mycotoxin

| Name | OTA | OTB | OTC |
|---|---|---|---|
| CAS number | 303-47-9 | 4825-86-9 | 4865-85-4 |
| Molecular formula | C20H18ClNO6 | C20H19NO6 | C22H22ClNO6 |
| Molar mass (g·mol−1) | 403.8 | 369.4 | 431.9 |
2. Occurrence of Ochratoxins
2.1. Biosynthesis

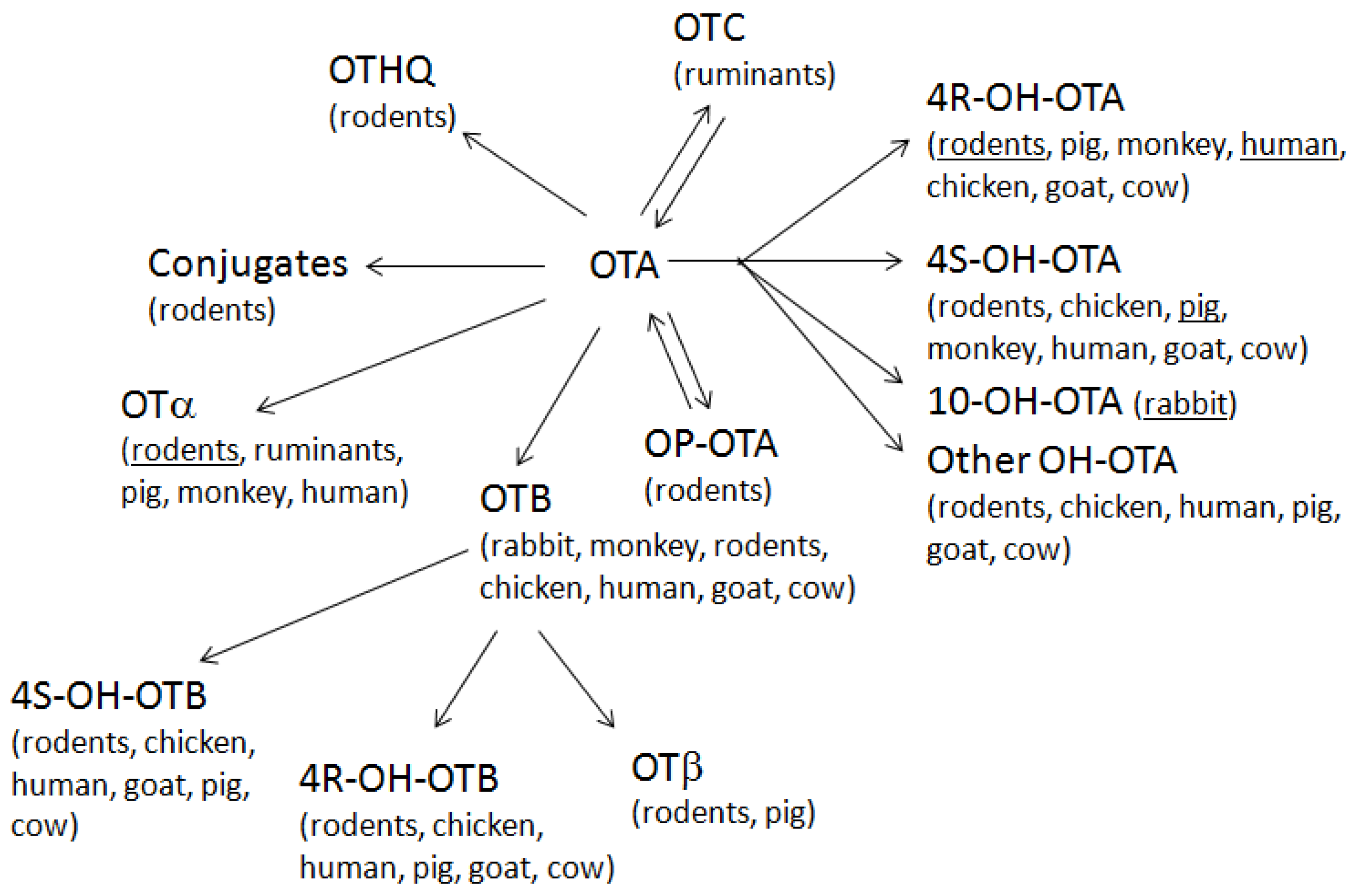{kind=link}
{kind=link}
| Name | Abbrev. | R1 and Others | R2 | R3 | R4 | R5 |
|---|---|---|---|---|---|---|
| Naturally occurring ochratoxins, metabolites and conjugates | ||||||
| Ochratoxin A | OTA | Phenylalanyl | Cl | H | H | H |
| Ochratoxin B | OTB | Phenylalanyl | H | H | H | H |
| Ochratoxin C | OTC | Phenylalanyl, ethyl ester | Cl | H | H | H |
| Ochratoxin A methyl ester | -- | Phenylalanyl, methyl ester | Cl | H | H | H |
| Ochratoxin B methyl ester | -- | Phenylalanyl, methyl ester | H | H | H | H |
| Ochratoxin B ethyl ester | -- | Phenylalanyl, ethyl ester | H | H | H | H |
| Ochratoxin α | OTα | OH | Cl | H | H | H |
| Ochratoxin β | OTβ | OH | H | H | H | H |
| 5′-Hydroxyochratoxin A | 5′-OH-OTA | Phenylalanyl, OH at C-5′ | Cl | H | H | H |
| 7′-Hydroxyochratoxin A | 7′-OH-OTA | Phenylalanyl, OH at C-7′ | Cl | H | H | H |
| 9′-Hydroxyochratoxin A | 9′-OH-OTA | Phenylalanyl, OH at C-9′ | Cl | H | H | H |
| 4R-Hydroxyochratoxin A | 4R-OH-OTA | Phenylalanyl | Cl | H | OH | H |
| 4S-Hydroxyochratoxin A | 4S-OH-OTA | Phenylalanyl | Cl | OH | H | H |
| 4R-Hydroxyochratoxin B | 4R-OH-OTB | Phenylalanyl | H | H | OH | H |
| 4S-Hydroxyochratoxin B | 4S-OH-OTB | Phenylalanyl | H | OH | H | H |
| 10-Hydroxyochratoxin A | 10-OH-OTA | Phenylalanyl | Cl | H | H | OH |
| Open lactone of ochratoxin A | OP-OTA | Phenylalanyl | Cl | H | H | H |
| Ochratoxin hydroquinone | OTHQ | Phenylalanyl | OH | H | H | H |
| Ochratoxin quinone | OTQ | Phenylalanyl | O | H | H | H |
| Ochratoxin α glucuronide | -- | Glucuronide | Cl | H | H | H |
| -- | OH; glucuronide at C-8 | Cl | H | H | H | |
| Ochratoxin A phenol-glucuronide | -- | Phenylalanyl; glucuronide at C-8 | Cl | H | H | H |
| Ochratoxin A amino-glucuronide | -- | Phenylalanyl; glucuronide at N | Cl | H | H | H |
| Ochratoxin A acyl-glucuronide | -- | Phenylalanyl; acyl-glucuronide instead of carboxyl | Cl | H | H | H |
| Ochratoxin A acyl-hexose | -- | Phenylalanyl; acyl-hexose instead of carboxyl | Cl | H | H | H |
| Ochratoxin A acyl-pentose | -- | Phenylalanyl; acyl-pentose instead of carboxyl | Cl | H | H | H |
| Synthetic ochratoxins | ||||||
| Ochratoxin A, tyrosine analogue | -- | Tyrosine | Cl | H | H | H |
| Ochratoxin A, serine analogue | -- | Serine | Cl | H | H | H |
| Ochratoxin A, hydroxyproline analogue | -- | Hydroxyproline | Cl | H | H | H |
| Ochratoxin A, lysine analogue | -- | Lysine | Cl | H | H | H |
| Ochratoxin A, alanine analogue | -- | Alanine | Cl | H | H | H |
| Ochratoxin A, leucine analogue | -- | Leucine | Cl | H | H | H |
| d-Ochratoxin A | d-OTA | d-Phenylalanyl | Cl | H | H | H |
| Ochratoxin A, ethylamide | OE-OTA | Phenylalanyl, ethylamide | Cl | H | H | H |
| O-methylated ochratoxin A | OM-OTA | Phenylalanyl, OCH3 at C-8 | Cl | H | H | H |
| Methyl ester of ochratoxin α | M-OTα | Methoxy | Cl | H | H | H |
| Ochratoxin A, decarboxylated | DC-OTA | Phenylethylamine | Cl | H | H | H |
| Other related compounds | ||||||
| Ochracin (=mellein) | -- | H at C-7 | H | H | H | H |
2.2. Contamination Levels and Human Exposure
2.3. Toxicokinetics of Ochratoxins

2.4. Detection Methods for Ochratoxins
3. Differential Toxicity of Ochratoxin Group Members
3.1. Comparative Toxicity of Ochratoxin Group Members
3.2. Ochratoxin B (OTB)
| Cell Type (Gender) | 96 h EC50 | 24–96 h EC50 | 48 h EC50 | ||||
|---|---|---|---|---|---|---|---|
| NR | MTT | CN | NR | MTT | CN | CV | |
| HKC (f) | OTA ↑ | OTA ↑ | OTA ↑ | n.d. | n.d. | n.d. | OTA ↑ |
| HKC (m) | Equal | Equal | Equal | Equal | Equal | Equal | OTA ↑ |
| PKC (f) | Equal | Equal | Equal | n.d. | n.d. | n.d. | Equal |
| PKC (m) | Equal | Equal | Equal | Equal | Equal | Equal | Equal |
| RPTC (f) | OTA ↑ | OTA ↑ | n.d. | n.d. | n.d. | n.d. | n.d. |
| RPTC (m) | OTA ↑ | OTA ↑ | n.d. | n.d. | n.d. | n.d. | n.d. |
| LLC-PK1 | OTA ↑ | OTA ↑ | Equal | OTA ↑ | OTA ↑ | Equal | OTA ↑ |
| NRK-52E | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | OTA ↑ |
| IHKE | n.d. | n.d. | n.d. | Equal | Equal | Equal | n.d. |

| Species (Gender) | [3H]OTA | [3H]OTB | p | ||||
|---|---|---|---|---|---|---|---|
| SB (pmol mg−1 Protein) | SB (%) | n | SB (pmol mg−1 Protein) | SB (%) | n | ||
| Human (m) | 21.4 ± 4.5 | 20.5 ± 4.1 | 6 | 1.25 ± 0.15 | 52.9 ± 10.7 | 3 | 0.0002 |
| Human (f) | 23.1 ± 4.7 | 22.0 ± 4.3 | 4 | 3.79 ± 2.04 | 73.6 ± 4.0 | 3 | <0.0001 |
| Rat (m) | 10.2 ± 3.7 | 10.1 ± 3.4 | 14 | 0.44 ± 0.42 | 31.2 ± 15.0 | 8 | <0.0001 |
| Rat (f) | 14.2 ± 6.3 | 11.5 ± 4.7 | 10 | 0.73 ± 0.39 | 43.3 ± 11.4 | 6 | <0.0001 |
| Pig (m) | 3.4 ± 1.0 | 3.3 ± 0.9 | 8 | 0.19 ± 011 | 27.2 ± 13.0 | 5 | 0.0002 |
| Pig (f) | 2.1 ± 0.7 | 2.0 ± 0.6 | 8 | <LOD | <LOD | 2 | n.c. |
| Mouse (m) | 0.9 ± 02 | 0.9 ± 0.2 | 6 | 0.16 | 14.3 | 1 | n.c. |
| Mouse (f) | 1.5 ± 0.6 | 1.5 ± 0.5 | 9 | <LOD | <LOD | 2 | n.c. |
3.3. Ochratoxin C (OTC) and Other Ethyl or Methyl Esters of Ochratoxins
3.4. Ochratoxin α (OTα) and Ochratoxin β (OTβ)
3.5. Hydroxyochratoxins
3.6. Ochratoxin Quinone (OTQ)/Hydroquinone (OTHQ)
3.7. Lactone-Opened OTA (OP-OTA)
3.8. Conjugated Ochratoxin Forms
3.9. Synthetic OTA Analogues
3.10. Structure–Activity Relationship
4. Toxicity of Mixtures of Mycotoxins
5. Summary and Final Remarks
Acknowledgments
Authors Contributions
Conflicts of Interest
Abbreviations
References
- Van der Merwe, K.; Steyn, P.; Fourie, L.; Scott, D.; Theron, J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.; Dietrich, D. Mycotoxins affecting the kidney. In Toxicology of Kidney; Hook, J., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 895–936. [Google Scholar]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed-focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Bellver Soto, J.; Fernández-Franzón, M.; Ruiz, M.; Juan-García, A. Presence of ochratoxin A (OTA) mycotoxin in alcoholic drinks from southern European countries: Wine and beer. J. Agric. Food Chem. 2014, 62, 7643–7651. [Google Scholar] [CrossRef] [PubMed]
- EC. Assessment of Dietary Intake of Ochratoxin A by the Population of EU Member States; Report of the Scientific Cooperation, Task 3.2.7; Directorate-General Health and Consumer Protection, European Commission: Rome, Italy, 2002. [Google Scholar]
- Haighton, L.A.; Lynch, B.S.; Magnuson, B.A.; Nestmann, E.R. A reassessment of risk associated with dietary intake of ochratoxin A based on a lifetime exposure model. Crit. Rev. Toxicol. 2012, 42, 147–168. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.; Dietrich, D.R. Ochratoxin A: The continuing enigma. Crit. Rev. Toxicol. 2005, 35, 33–60. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Opinion of the scientific panel on contaminants in the food chain on a request from the commission related to ochratoxin A in food. EFSA J. 2006, 365, 1–56. [Google Scholar]
- Duarte, S.; Lino, C.; Pena, A. Food safety implications of ochratoxin A in animal-derived food products. Vet. J. 2012, 192, 286–292. [Google Scholar] [CrossRef] [PubMed]
- FAO. Recommended practices for the prevention of mycotoxins in food, feed and their products. FAO Food Nutr. Pap. 1979, 10, 1–71. [Google Scholar]
- Visconti, A.; Bottalico, A. High levels of ochratoxins A and B in moldy bread responsible for mycotoxicosis in farm animals. J. Agric. Food Chem. 1983, 31, 1122–1123. [Google Scholar] [CrossRef] [PubMed]
- Huff, W.E.; Wyatt, R.D.; Tucker, T.L.; Hamilton, P.B. Ochratoxicosis in the broiler chicken. Poult. Sci. 1974, 53, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, P.B.; Huff, W.E.; Harris, J.R.; Wyatt, R.D. Natural occurrences of ochratoxicosis in poultry. Poult. Sci. 1982, 61, 1832–1841. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, P.; Burns, R.B.; Maxwell, M.H. Ultrastructural study of the liver and kidney in ochratoxicosis A in young broiler chicks. Res. Vet. Sci. 1984, 36, 104–116. [Google Scholar] [PubMed]
- Dwivedi, P.; Burns, R.B. Pathology of ochratoxicosis A in young broiler chicks. Res. Vet. Sci. 1984, 36, 92–103. [Google Scholar] [PubMed]
- Kozaczynski, W. Experimental ochratoxicosis A in chickens. Histopathological and histochemical study. Arch. Vet. Pol./Pol. Acad. Sci. Comm. Vet. Sci. 1994, 34, 205–219. [Google Scholar]
- Elaroussi, M.A.; Mohamed, F.R.; el Barkouky, E.M.; Atta, A.M.; Abdou, A.M.; Hatab, M.H. Experimental ochratoxicosis in broiler chickens. Avian Pathol. J. WVPA 2006, 35, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.; Dutton, M.; Njobeh, P.; Mosonik, J.; Steenkamp, P. Mycotoxic nephropathy in Bulgarian pigs and chickens: Complex aetiology and similarity to Balkan endemic nephropathy. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2010, 27, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Elling, F.; Moller, T. Mycotoxic nephropathy in pigs. Bull. World Health Organ. 1973, 49, 411–418. [Google Scholar] [PubMed]
- Stoev, S.; Denev, S. Porcine/chicken or human nephropathy as the result of joint mycotoxins interaction. Toxins 2013, 5, 1503–1530. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.; Denev, S.; Dutton, M.; Njobeh, P.; Mosonik, J.; Steenkamp, P.; Petkov, I. Complex etiology and pathology of mycotoxic nephropathy in South African pigs. Mycotoxin Res. 2010, 26, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Krogh, P. Epidemiology of mycotoxic porcine nephropathy. Nord. Vet. Med. 1976, 28, 452–458. [Google Scholar] [PubMed]
- Stoev, S.; Gundasheva, D.; Zarkov, I.; Mircheva, T.; Zapryanova, D.; Denev, S.; Mitev, Y.; Daskalov, H.; Dutton, M.; Mwanza, M.; et al. Experimental mycotoxic nephropathy in pigs provoked by a mouldy diet containing ochratoxin A and fumonisin B1. Exp. Toxicol. Pathol. 2012, 64, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.; Vitanov, S.; Anguelov, G.; Petkova-Bocharova, T.; Creppy, E. Experimental mycotoxic nephropathy in pigs provoked by a diet containing ochratoxin A and penicillic acid. Vet. Res. Commun. 2001, 25, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Goliński, P.; Hult, K.; Grabarkiewicz-Szczesna, J.; Chełkowski, J.; Kneblewski, P.; Szebiotko, K. Mycotoxic porcine nephropathy and spontaneous occurrence of ochratoxin A residues in kidneys and blood of Polish swine. Appl. Environ. Microbiol. 1984, 47, 1210–1212. [Google Scholar] [PubMed]
- Stoev, S. The role of ochratoxin A as a possible cause of Balkan endemic nephropathy and its risk evaluation. Vet. Hum. Toxicol. 1998, 40, 352–360. [Google Scholar] [PubMed]
- Abid, S.; Hassen, W.; Achour, A.; Skhiri, H.; Maaroufi, K.; Ellouz, F.; Creppy, E.; Bacha, H. Ochratoxin A and human chronic nephropathy in Tunisia: Is the situation endemic? Hum. Exp. Toxicol. 2003, 22, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Hmaissia Khlifa, K.; Ghali, R.; Mazigh, C.; Aouni, Z.; Machgoul, S.; Hedhili, A. Ochratoxin A levels in human serum and foods from nephropathy patients in Tunisia: Where are you now? Exp. Toxicol. Pathol. 2012, 64, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Grollman, A.P.; Jelaković, B. Role of environmental toxins in Endemic (Balkan) Nephropathy. J. Am. Soc. Nephrol. 2007, 18, 2817–2823. [Google Scholar] [CrossRef] [PubMed]
- Maaroufi, K.; Pfohl-Leszkowicz, A.; Achour, A.; el May, M.; Grosse, Y.; Hammami, M.; Ellouz, F.; Creppy, E.; Bacha, H. Ochratoxin A genotoxicity, relation to renal tumors. Arch. Inst. Pasteur Tunis 1994, 71, 21–31. [Google Scholar] [PubMed]
- Pepeljnjak, S.; Klarić, M. «Suspects» in etiology of endemic nephropathy: Aristolochic acid versus mycotoxins. Toxins 2010, 2, 1414–1427. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Knox, B.P.; Bruno, K.S.; Solfrizzo, M.; Baker, S.E.; Perrone, G. Identification and characterization of the polyketide synthase involved in ochratoxin A biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2014, 179, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Medina, Á.; Córdoba, J.J.; Magan, N. The influence of salt (NaCl) on ochratoxin A biosynthetic genes, growth and ochratoxin A production by three strains of Penicillium nordicum on a dry-cured ham-based medium. Int. J. Food Microbiol. 2014, 178, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Wang, Q.; Liu, F.; Selvaraj, J.N.; Liu, L.; Xing, F.; Zhao, Y.; Zhou, L.; Liu, Y. Functional characterization of new polyketide synthase genes involved in ochratoxin A biosynthesis in Aspergillus ochraceus fc-1. Toxins 2015, 7, 2723–2738. [Google Scholar] [CrossRef] [PubMed]
- Färber, P.; Geisen, R. Analysis of differentially-expressed ochratoxin A biosynthesis genes of Penicillium nordicum. Eur. J. Plant Pathol. 2004, 110, 661–669. [Google Scholar] [CrossRef]
- Geisen, R.; Schmidt-Heydt, M.; Karolewiez, A. A gene cluster of the ochratoxin A biosynthetic genes inPenicillium. Mycotox Res. 2006, 22, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Sartori, D.; Massi, F.P.; Ferranti, L.S.; Fungaro, M.H. Identification of genes differentially expressed between ochratoxin-producing and non-producing strains of Aspergillus westerdijkiae. Indian J. Microbiol. 2014, 54, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Huffman, J.; Gerber, R.; Du, L. Recent advancements in the biosynthetic mechanisms for polyketide-derived mycotoxins. Biopolymers 2010, 93, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Ringot, D.; Chango, A.; Schneider, Y.J.; Larondelle, Y. Toxicokinetics and toxicodynamics of ochratoxin A, an update. Chem. Biol. Interact. 2006, 159, 18–46. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, J.; Caddick, M.X.; Dobson, A.D. A polyketide synthase gene required for ochratoxin A biosynthesis in Aspergillus ochraceus. Microbiology 2003, 149, 3485–3491. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, J.; Stapleton, P.C.; Dobson, A.D. Ochratoxin A biosynthetic genes in Aspergillus ochraceus are differentially regulated by pH and nutritional stimuli. Fungal Genet. Biol. 2006, 43, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Bacha, N.; Atoui, A.; Mathieu, F.; Liboz, T.; Lebrihi, A. Aspergillus westerdijkiae polyketide synthase gene “aoks1” is involved in the biosynthesis of ochratoxin A. Fungal Genet. Biol. 2009, 46, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karolewiez, A.; Geisen, R. Cloning a part of the ochratoxin A biosynthetic gene cluster of Penicillium nordicum and characterization of the ochratoxin polyketide synthase gene. Syst. Appl. Microbiol. 2005, 28, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Valez, H.; Dobson, A.D. Analysis of the effect of nutritional factors on OTA and OTB biosynthesis and polyketide synthase gene expression in Aspergillus ochraceus. Int. J. Food Microbiol. 2009, 135, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Kevei, E.; Rinyu, E.; Téren, J.; Kozakiewicz, Z. Ochratoxin production by Aspergillus species. Appl. Environ. Microbiol. 1996, 62, 4461–4464. [Google Scholar] [PubMed]
- Harris, J.; Mantle, P. Biosynthesis of ochratoxins by Aspergillus ochraceus. Phytochemistry 2001, 58, 709–716. [Google Scholar] [CrossRef]
- Davis, N.; Searcy, J.; Diener, U. Production of ochratoxin A by Aspergillus ochraceus in a semisynthetic medium. Appl. Microbiol. 1969, 17, 742–744. [Google Scholar] [PubMed]
- Trenk, H.; Butz, M.; Chu, F. Production of ochratoxins in different cereal products by Aspergillus ochraceus. Appl. Microbiol. 1971, 21, 1032–1035. [Google Scholar] [PubMed]
- Mantle, P.G.; Chow, A.M. Ochratoxin formation in Aspergillus ochraceus with particular reference to spoilage of coffee. Int. J. Food Microbiol. 2000, 56, 105–109. [Google Scholar] [CrossRef]
- Hesseltine, C.W. Solid State Fermentations. Biotechnol. Bioeng. 1972, 9, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Azpilicueta, C.A.; Arbeloa, M.I.; de Maquirriain, P.F.J.; Moreno, N.J. Study of ochratoxin A in wines from sun-dired grapes and in commercial wines from different geographical origins. In Food Chemistry Research Developments; Papadopoulos, K., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2008; pp. 113–139. [Google Scholar]
- Yang, S.; Zhang, H.; de Saeger, S.; de Boevre, M.; Sun, F.; Zhang, S.; Cao, X.; Wang, Z. In vitro and in vivo metabolism of ochratoxin A: A comparative study using ultra-performance liquid chromatography-quadrupole/time-of-flight hybrid mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 3579–3589. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Dohnal, V.; Huang, L.; Kuča, K.; Wang, X.; Chen, G.; Yuan, Z. Metabolic pathways of ochratoxin A. Curr. Drug Metab. 2011, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Bruinink, A.; Rasonyi, T.; Sidler, C. Differences in neurotoxic effects of ochratoxin A, ochracin and ochratoxin-α in vitro. Nat. Toxins 1998, 6, 173–177. [Google Scholar] [CrossRef]
- Tozlovanu, M.; Faucet-Marquis, V.; Pfohl-Leszkowicz, A.; Manderville, R.A. Genotoxicity of the hydroquinone metabolite of ochratoxin A: Structure-activity relationships for covalent DNA adduction. Chem. Res. Toxicol. 2006, 19, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Tangni, E.K.; di Mavungu, J.D.; Vanhaecke, L.; de Saeger, S.; Wu, A.; Callebaut, A. In vitro glucuronidation of ochratoxin A by rat liver microsomes. Toxins 2013, 5, 2671–2685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mally, A.; Keim-Heusler, H.; Amberg, A.; Kurz, M.; Zepnik, H.; Mantle, P.; Völkel, W.; Hard, G.C.; Dekant, W. Biotransformation and nephrotoxicity of ochratoxin B in rats. Toxicol. Appl. Pharmacol. 2005, 206, 43–53. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Selected Mycotoxins: Ochratoxins, Trichothecenes, Ergot; WHO-IPCS: Geneva, Switzerland, 1990. [Google Scholar]
- Tabata, S.; Iida, K.; Kimura, K.; Iwasaki, Y.; Nakazato, M.; Kamata, K.; Hirokado, M. Investigation of ochratoxin A, B and citrinin contamination in various commercial foods. Shokuhin Eiseigaku Zasshi 2008, 49, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Scheuer, R.; Gareis, M. Occurrence of ochratoxin A and B in spices. Mycotoxin Res. 2002, 18, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, V.; Avellone, G.; Pitonzo, R.; Capocchiano, V.G.; Mazza, A.; Cicero, N.; Dugo, G. Natural co-occurrence of ochratoxin A, ochratoxin B and aflatoxins in Sicilian red wines. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Mally, A. Ochratoxin a and mitotic disruption: Mode of action analysis of renal tumor formation by ochratoxin A. Toxicol. Sci. 2012, 127, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Heyndrickx, E.; Sioen, I.; Huybrechts, B.; Callebaut, A.; de Henauw, S.; de Saeger, S. Human biomonitoring of multiple mycotoxins in the Belgian population: Results of the BIOMYCO study. Environ. Int. 2015, 84, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Dixit, R.; Riviere, J.; Krishnan, K.; Andersen, M.E. Toxicokinetics and physiologically based toxicokinetics in toxicology and risk assessment. J. Toxicol. Environ. Health B Crit. Rev. 2003, 6, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, R.; Hult, K. Ochratoxin A in blood and its pharmacokinetic properties. Food Chem. Toxicol. 1992, 30, 201–204. [Google Scholar] [CrossRef]
- Galtier, P. Pharmacokinetics of ochratoxin A in animals. IARC Sci. Publ. 1991, 115, 187–200. [Google Scholar] [PubMed]
- Coronel, M.; Sanchis, V.; Ramos, A.; Marin, S. Ochratoxin A: Presence in human plasma and intake estimation. Food Sci. Technol. Int. 2010, 16, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Vettorazzi, A.; González-Peñas, E.; de Cerain, A. Ochratoxin A kinetics: A review of analytical methods and studies in rat model. Food Chem. Toxicol. 2014, 72, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Mally, A.; Dekant, W. Mycotoxins and the kidney: Modes of action for renal tumor formation by ochratoxin A in rodents. Mol. Nutr. Food Res. 2009, 53, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, D.R.; Heussner, A.H.; O’Brien, E. Ochratoxin A: Comparative pharmacokinetics and toxicological implications (experimental and domestic animals and humans). Food Addit. Contam. 2005, 22, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Schlatter, C.; Studer-Rohr, J.; Rásonyi, T. Carcinogenicity and kinetic aspects of ochratoxin A. Food Addit. Contam. 1996, 13 (Suppl.), 43–44. [Google Scholar] [PubMed]
- Marquardt, R.R.; Frohlich, A.A. A review of recent advances in understanding ochratoxicosis. J. Anim. Sci. 1992, 70, 3968–3988. [Google Scholar] [PubMed]
- Breitholtz Emanuelsson, A.; Fuchs, R.; Hult, K.; Appelgren, L. Distribution of 14C-ochratoxin A and 14C-ochratoxin B in rats: A comparison based on whole-body autoradiography. IARC Sci. Publ. 1991, 115, 201–203. [Google Scholar] [PubMed]
- Breitholtz-Emanuelsson, A.; Fuchs, R.; Hult, K.; Appelgren, L. Synthesis of 14C-ochratoxin A and 14C-ochratoxin B and a comparative study of their distribution in rats using whole body autoradiography. Pharmacol. Toxicol. 1992, 70, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Hagelberg, S.; Hult, K.; Fuchs, R. Toxicokinetics of ochratoxin A in several species and its plasma-binding properties. J. Appl. Toxicol. 1989, 9, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Galtier, P.; Charpenteau, J.-L.; Alvinerie, M.; Labouche, C. The pharmacokinetic profile of ochratoxin A in the rat after oral and intravenous administration. Drug Metab. Dispos. 1979, 7, 429–434. [Google Scholar] [PubMed]
- Abrunhosa, L.; Santos, L.; Venâncio, A. Degradation of ochratoxin A by proteases and by a crude enzyme of Aspergillus niger. Food Biotechnol. 2006, 20, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Støren, O.; Holm, H.; Størmer, F. Metabolism of ochratoxin A by rats. Appl. Environ. Microbiol. 1982, 44, 785–789. [Google Scholar] [PubMed]
- World Health Organization (WHO). Safety Evaluation of Certain Mycotoxins in Food; WHO-IPCS: Geneva, Switzerland, 2001. [Google Scholar]
- Pinelli, E.; el Adlouni, C.; Pipy, B.; Quartulli, F.; Pfohl-Leszkowicz, A. Roles of cyclooxygenase and lipoxygenases in ochratoxin A genotoxicity in human epithelial lung cells. Environ. Toxicol. Pharmacol. 1999, 7, 95–107. [Google Scholar] [CrossRef]
- Størmer, F.; Hansen, C.; Pedersen, J.; Hvistendahl, G.; Aasen, A. Formation of (4R)- and (4S)-4-hydroxyochratoxin A from ochratoxin A by liver microsomes from various species. Appl. Environ. Microbiol. 1981, 42, 1051–1056. [Google Scholar] [PubMed]
- Stein, A.; Phillips, T.; Kubena, L.; Harvey, R. Renal tubular secretion and reabsorption as factors inochratoxicosis: Effects of probenecid on nephrotoxicity. J. Toxicol. Environ. Health 1985, 16, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Størmer, F.; Støren, O.; Hansen, C.; Pedersen, J.; Aasen, A. Formation of (4R)- and (4S)-4-hydroxyochratoxin A and 10-hydroxyochratoxin A from Ochratoxin A by rabbit liver microsomes. Appl. Environ. Microbiol. 1983, 45, 1183–1187. [Google Scholar] [PubMed]
- Creppy, E.E.; Størmer, F.C.; Röschenthaler, R.; Dirheimer, G. Effects of two metabolites of ochratoxin A, (4R)-4-hydroxyochratoxin A and ochratoxin alpha, on immune response in mice. Infect. Immun. 1983, 39, 1015–1018. [Google Scholar] [PubMed]
- Xiao, H.; Madhyastha, S.; Marquardt, R.R.; Li, S.; Vodela, J.K.; Frohlich, A.A.; Kemppainen, B.W. Toxicity of ochratoxin A, its opened lactone form and several of its analogs: Structure-activity relationships. Toxicol. Appl. Pharmacol. 1996, 137, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Zepnik, H.; Paehler, A.; Schauer, U.; Dekant, W. Ochratoxin A-induced tumor formation: Is there a role of reactive ochratoxin A metabolites? Toxicol. Sci. 2001, 59, 59–67. [Google Scholar] [CrossRef] [PubMed]
- El Adlouni, C.; Pinelli, E.; Azémar, B.; Zaoui, D.; Beaune, P.; Pfohl-Leszkowicz, A. Phenobarbital increases DNA adduct and metabolites formed by ochratoxin A: Role of CYP 2C9 and microsomal glutathione-S-transferase. Environ. Mol. Mutagen. 2000, 35, 123–131. [Google Scholar] [CrossRef]
- Roth, A.; Chakor, K.; Creppy, E.E.; Kane, A.; Roschenthaler, R.; Dirheimer, G. Evidence for an enterohepatic circulation of ochratoxin A in mice. Toxicology 1988, 48, 293–308. [Google Scholar] [CrossRef]
- Kühn, I.; Valenta, H.; Rohr, K. Determination of ochratoxin A in bile of swine by high-performance liquid chromatography. J. Chromatogr. B Biomed. Appl. 1995, 668, 333–337. [Google Scholar] [CrossRef]
- Gross-Steinmeyer, K.; Weymann, J.; Hege, H.; Metzler, M. Metabolism and lack of DNA reactivity of the mycotoxin ochratoxin a in cultured rat and human primary hepatocytes. J. Agric. Food Chem. 2002, 50, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Zepnik, H.; Völkel, W.; Dekant, W. Toxicokinetics of the mycotoxin ochratoxin A in F344 rats after oral administration. Toxicol. Appl. Pharmacol. 2003, 192, 36–44. [Google Scholar] [CrossRef]
- Malaveille, C.; Brun, G.; Bartsch, H. Structure-activity studies in E. coli strains on ochratoxin A (OTA) and its analogues implicate a genotoxic free radical and a cytotoxic thiol derivative as reactive metabolites. Mutat. Res. 1994, 307, 141–147. [Google Scholar] [CrossRef]
- Dai, J.; Park, G.; Wright, M.W.; Adams, M.; Akman, S.A.; Manderville, R.A. Detection and characterization of a glutathione conjugate of ochratoxin A. Chem. Res. Toxicol. 2002, 15, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J.; Jahn, A.; Blom, M.J. Toxicity and metabolism of ochratoxin A. Nat. Toxins 1995, 3, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Marquardt, R.R.; Frohlich, A.A. Identification of ochratoxins and some of their metabolites in bile and urine of rats. Food Chem. Toxicol. 2000, 38, 141–152. [Google Scholar] [CrossRef]
- Turner, N.W.; Subrahmanyam, S.; Piletsky, S.A. Analytical methods for determination of mycotoxins: A review. Anal. Chim. Acta 2009, 632, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.M. Methods of analysis for ochratoxin A. Adv. Exp. Med. Biol. 2002, 504, 117–134. [Google Scholar]
- Visconti, A.; de Girolamo, A. Fitness for purpose—Ochratoxin A analytical developments. Food Addit. Contam. 2005, 22, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Meulenberg, E.P. Immunochemical methods for ochratoxin A detection: A review. Toxins 2012, 4, 244–266. [Google Scholar] [CrossRef] [PubMed]
- Moeller, I.; O’Brien, E.; Heussner, A.; Day, B.; Dietrich, D. Monoclonal Antibodies Against Ochratoxin B: Production and Characterisation. In Proceedings of the Joint Speciality Symposium on Renal Toxicology and Toxicologic Pathology, Lindau, Germany, 27 September–1 October 2004.
- Heussner, A.H.; Moeller, I.; Day, B.W.; Dietrich, D.R.; O’Brien, E. Production and characterization of monoclonal antibodies against ochratoxin B. Food Chem. Toxicol. 2007, 45, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Ausländer, S.; Dietrich, D.R. Development and characterization of a monoclonal antibody against ochratoxin B and its application in ELISA. Toxins 2010, 2, 1582–1594. [Google Scholar] [CrossRef] [PubMed]
- Glaze, T.; Heussner, A.; Dietrich, D.; Rubio, F. ELISA and sample clean-up methods for determination of ochratoxin B. In Proceedings of the 5th International Symposium on recent advances in food analyses (RAFA), Prague, Czech Recpublic, 1–4 November 2011.
- Auslaender, S.T.; Heussner, A.H.; Dietrich, D.R. Monoclonal antibodies against OTB: Potential future applications. Toxicol. Sci. (Toxicologist) 2008, 102, 100. Available online: http://www.toxicology.org/pubs/docs/Tox/2008Tox.pdf (accessed on 20 October 2015). [Google Scholar]
- Doi, K.; Uetsuka, K. Mechanisms of mycotoxin-induced dermal toxicity and tumorigenesis through oxidative stress-related pathways. J. Toxicol. Pathol. 2014, 27, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Limonciel, A.; Jennings, P. A Review of the evidence that ochratoxin A is an Nrf2 inhibitor: Implications for nephrotoxicity and renal carcinogenicity. Toxins 2014, 6, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Malir, F.; Ostry, V.; Novotna, E. Toxicity of the mycotoxin ochratoxin A in the light of recent data. Toxin Rev. 2013, 32, 19–33. [Google Scholar] [CrossRef]
- Klarić, M.S.; Rašić, D.; Peraica, M. Deleterious effects of mycotoxin combinations involving ochratoxin A. Toxins 2013, 5, 1965–1987. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Bognanno, M.; Galvano, F. Toxicity of ochratoxin A and its modulation by antioxidants: A review. Toxins (Basel) 2013, 5, 1742–1766. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Vettorazzi, A.; van Delft, J.; López de Cerain, A. A review on ochratoxin A transcriptomic studies. Food Chem. Toxicol. 2013, 59, 766–783. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Manderville, R.A. An update on direct genotoxicity as a molecular mechanism of ochratoxin A carcinogenicity. Chem. Res. Toxicol. 2012, 25, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Uetsuka, K. Mechanisms of mycotoxin-induced neurotoxicity through oxidative stress-associated pathways. Int. J. Mol. Sci. 2011, 12, 5213–5237. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R.; Lima, N. Toxicology of mycotoxins. EXS 2010, 100, 31–63. [Google Scholar] [PubMed]
- Pfohl-Leszkowicz, A. Ochratoxin A and aristolochic acid involvement in nephropathies and associated urothelial tract tumours. Arh. Hig. Rada Toksikol. 2009, 60, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Marin-Kuan, M.; Cavin, C.; Delatour, T.; Schilter, B. Ochratoxin A carcinogenicity involves a complex network of epigenetic mechanisms. Toxicon 2008, 52, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An overview on toxicity and carcinogenicity in animals and humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar] [CrossRef] [PubMed]
- Clark, H.A.; Snedeker, S.M. Ochratoxin a: Its cancer risk and potential for exposure. J. Toxicol. Environ. Health B Crit. Rev. 2006, 9, 265–296. [Google Scholar] [CrossRef] [PubMed]
- Al-Anati, L.; Petzinger, E. Immunotoxic activity of ochratoxin A. J. Vet. Pharmacol. Ther. 2006, 29, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Schilter, B.; Marin-Kuan, M.; Delatour, T.; Nestler, S.; Mantle, P.; Cavin, C. Ochratoxin A: Potential epigenetic mechanisms of toxicity and carcinogenicity. Food Addit. Contam. 2005, 22, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Mally, A.; Dekant, W. DNA adduct formation by ochratoxin A: Review of the available evidence. Food Addit. Contam. 2005, 22, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Turesky, R. Perspective: Ochratoxin A is not a genotoxic carcinogen. Chem. Res. Toxicol. 2005, 18, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Manderville, R.A. A case for the genotoxicity of ochratoxin A by bioactivation and covalent DNA adduction. Chem. Res. Toxicol. 2005, 18, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Gekle, M.; Sauvant, C.; Schwerdt, G. Ochratoxin A at nanomolar concentrations: A signal modulator in renal cells. Mol. Nutr. Food Res. 2005, 49, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Walker, R. Risk assessment of ochratoxin: Current views of the European Scientific Committee on Food, the JECFA and the Codex Committee on Food Additives and Contaminants. Adv. Exp. Med. Biol. 2002, 504, 249–255. [Google Scholar] [PubMed]
- Gekle, M.; Sauvant, C.; Schwerdt, G.; Silbernagl, S. Tubulotoxic mechanisms of ochratoxin A. Kidney Blood Press. Res. 1998, 21, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Fillastre, J.P. Experimental and human nephrotoxicity induced by ochratoxins. Bull. Acad. Natl. Med. 1997, 181, 1447–1460. [Google Scholar] [PubMed]
- Godin, M.; Fillastre, J.P.; Simon, P.; Francois, A.; Roy, F.L.; Morin, J.P. Is ochratoxin A nephrotoxic in human beings? Adv. Nephrol. Necker Hosp. 1997, 26, 181–206. [Google Scholar] [PubMed]
- Plestina, R. Nephrotoxicity of ochratoxin A. Food Addit. Contam. 1996, 13, 49–50. [Google Scholar] [PubMed]
- Dirheimer, G. Mechanistic approaches to ochratoxin toxicity. Food Addit. Contam. 1996, 13, 45–48. [Google Scholar] [PubMed]
- Gekle, M.; Silbernagl, S. Renal toxicodynamics of ochratoxin A: A pathophysiological approach. Kidney Blood Press. Res. 1996, 19, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.P. Immunotoxicity of mycotoxins. J. Dairy Sci. 1993, 76, 892–897. [Google Scholar] [CrossRef]
- Huff, J.E. Carcinogenicity of ochratoxin A in experimental animals. IARC Sci. Publ. 1991, 115, 229–244. [Google Scholar] [PubMed]
- Dirheimer, G.; Creppy, E.E. Mechanism of action of ochratoxin A. IARC Sci. Publ. 1991, 115, 171–186. [Google Scholar] [PubMed]
- Munro, C.; Scott, P.M.; Moodie, C.A.; Willes, R.F. Ochratoxin A—Occurrence and toxicity. J. Am. Vet. Med. Assoc. 1973, 163, 1269–1273. [Google Scholar] [PubMed]
- Rumora, L.; Grubisić, T.Z. A journey through mitogen-activated protein kinase and ochratoxin A interactions. Arh. Hig. Rada Toksikol. 2009, 60, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Marin-Kuan, M.; Ehrlich, V.; Delatour, T.; Cavin, C.; Schilter, B. Evidence for a role of oxidative stress in the carcinogenicity of ochratoxin A. J. Toxicol. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- IARC. Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1993. [Google Scholar]
- EFSA. Statement on recent scientific information on the toxicity of ochratoxin A. EFSA J. 2010, 8, 1–7. [Google Scholar]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Evaluation of Certain Food Additives and Contaminants; Technical Report Series No. 806 for WHO: Geneva, Switzerland, 1991. [Google Scholar]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Evaluation of Certain Food Additives and Contaminants; Technical Report Series No. 855 for WHO: Geneva, Switzerland, 1995. [Google Scholar]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Evaluation of Certain Mycotoxins in Food; Technical Report for Series No. 896 for WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Food Additives and Contaminants: Ochratoxin A (addendum); WHO: Geneva, Switzerland, 2007. [Google Scholar]
- European Commission. Maximum Levels for Certain Contaminants in Foodstuffs. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1445355561269&uri=CELEX:32006R1881 (accessed on 20 October 2015).
- NTP (National Toxicology Program). Toxicology and carcinogenesis studies of ochratoxin A (CAS No. 303-47-9) in F344/N rats (gavage studies); U.S. Department of Health and Human Services, Public Health Service, National Institutes of Health: Bethesda, MD, USA, 1989.
- NTP (National Toxicology Program). Report on Carcinogens (Ochratoxin A); U.S. Department of Health and Human Services, Public Health Service, National Institutes of Health: Bethesda, MD, USA, 2011.
- NTP (National Toxicology Program). Report on Carcinogens (Ochratoxin A); U.S. Department of Health and Human Services, Public Health Service, National Institutes of Health: Bethesda, MD, USA, 2014. Available online: https://ntp.niehs.nih.gov/ntp/roc/content/profiles/ochratoxina.pdf (accessed on 20 October 2015).
- Kuiper-Goodman, T.; Scott, P.M. Risk assessment of the mycotoxin ochratoxin A. Biomed. Environ. Sci. 1989, 2, 179–248. [Google Scholar] [PubMed]
- Kuiper-Goodman, T.; Hilts, C.; Billiard, S.M.; Kiparissis, Y.; Richard, I.D.; Hayward, S. Health risk assessment of ochratoxin A for all age-sex strata in a market economy. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2010, 27, 212–240. [Google Scholar] [CrossRef] [PubMed]
- Health Canada. Information Document on Health Canada’s Proposed Maximum Limits (Standards) for the Presence of the Mycotoxin Ochratoxin A in Foods. Bureau of Chemical Safety, Food Directorate, Health Products and Food Branch: Ottawa, ON, Cannada, 2009. Available online: http://www.hc-sc.gc.ca/fn-an/alt_formats/hpfb-dgpsa/pdf/consultation/myco_consult_ochra-eng.pdf (accessed on 20 October 2015).
- Egypt. Organization for Standardazation and Quality: Egyptian Standard 1875-2/2010 Maximum Limits for Mycotoxins in Foods-Ochratoxins; Egyptian Organization for Standardization and Quality Control: Cairo, Egypt, 2010. [Google Scholar]
- China’s Maximum Levels for Mycotoxins in Foods. Available online: http://gain.fas.usda.gov/Recent%20GAIN%20Publications/Maximum%20Levels%20of%20Mycotoxins%20in%20Foods_Beijing_China%20-%20Peoples%20Republic%20of_12-29-2014.pdf (accessed on 20 October 2015).
- Müller, G.; Burkert, B.; Möller, U.; Diller, R.; Rohrmann, B.; Rosner, H.; Köhler, H. Ochratoxin A and some of its derivatives modulate radical formation of porcine blood monocytes and granulocytes. Toxicology 2004, 199, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Peckham, J.; Doupnik, B.J.; Jones, O.J. Acute toxicity of ochratoxins A and B in chicks. Appl. Microbiol. 1971, 21, 492–494. [Google Scholar] [PubMed]
- Doster, R.C.; Sinnhuber, R.O.; Pawlowski, N.E. Acute intraperitoneal toxicity of ochratoxin A and B derivatives in rainbow trout (Salmo gairdneri). Food Cosmet. Toxicol. 1974, 12, 499–505. [Google Scholar] [CrossRef]
- Luhe, A.; Hildebrand, H.; Bach, U.; Dingermann, T.; Ahr, H. A new approach to studying ochratoxin A (OTA)-induced nephrotoxicity: Expression profiling in vivo and in vitro employing cDNA microarrays. Toxicol. Sci. 2003, 73, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, P.; Upadhyay, A.; Agnihotri, A.; Karmakar, S.; Ghoyary, D.; Veer, V. Comparative hematoxicity of fusirium mycotoxin in experimental sprague-dawley rats. Toxicol. Int. 2013, 20, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Bondy, G.S.; Armstrong, C.L. Cytotoxicity of nephrotoxic fungal toxins to kidney-derived LLC-PK1 and OK cell lines. Cell Biol. Toxicol. 1998, 14, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Bruinink, A.; Rasonyi, T.; Sidler, C. Reduction of ochratoxin A toxicity by heat-induced epimerization. In vitro effects of ochratoxins on embryonic chick meningeal and other cell cultures. Toxicology 1997, 118, 205–210. [Google Scholar] [PubMed]
- Knasmüller, S.; Cavin, C.; Chakraborty, A.; Darroudi, F.; Majer, B.J.; Huber, W.W.; Ehrlich, V.A. Structurally related mycotoxins ochratoxin A, ochratoxin B, and citrinin differ in their genotoxic activities and in their mode of action in human-derived liver (HepG2) cells: Implications for risk assessment. Nutr. Cancer 2004, 50, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Schwöbel, F.; Dietrich, D.R. Cytotoxicity of ochratoxin A and B in vitro: Comparison of male and female rat primary renal cortex and distal cells and LLC-PK1 cells. Toxicol. Sci. 1998, 42, 288. Available online: http://www.toxicology.org/pubs/docs/Tox/1998Tox.pdf (accessed on 20 October 2015). [Google Scholar]
- Heussner, A.H.; Stack, M.E.; Hochberg, K.; Dietrich, D.R. Comparison of cytotoxic effects of ochratoxin A and B on human, rat and porcine renal cells. Toxicol. Sci. 2000, 54, 170. Available online: http://www.toxicology.org/pubs/docs/Tox/2000Tox.pdf (accessed on 20 October 2015). [Google Scholar]
- Dietrich, D.R.; O’Brien, E.; Stack, M.E.; Heussner, A.H. Species- and sex-specific renal cytotoxicity of ochratoxin A and B in vitro. Exp. Toxicol. Pathol. 2001, 53, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; O’Brien, E.; Dietrich, D.R. Effects of repeated ochratoxin exposure on renal cells in vitro. Toxicol. Vitr. 2007, 21, 72–80. [Google Scholar] [CrossRef] [PubMed]
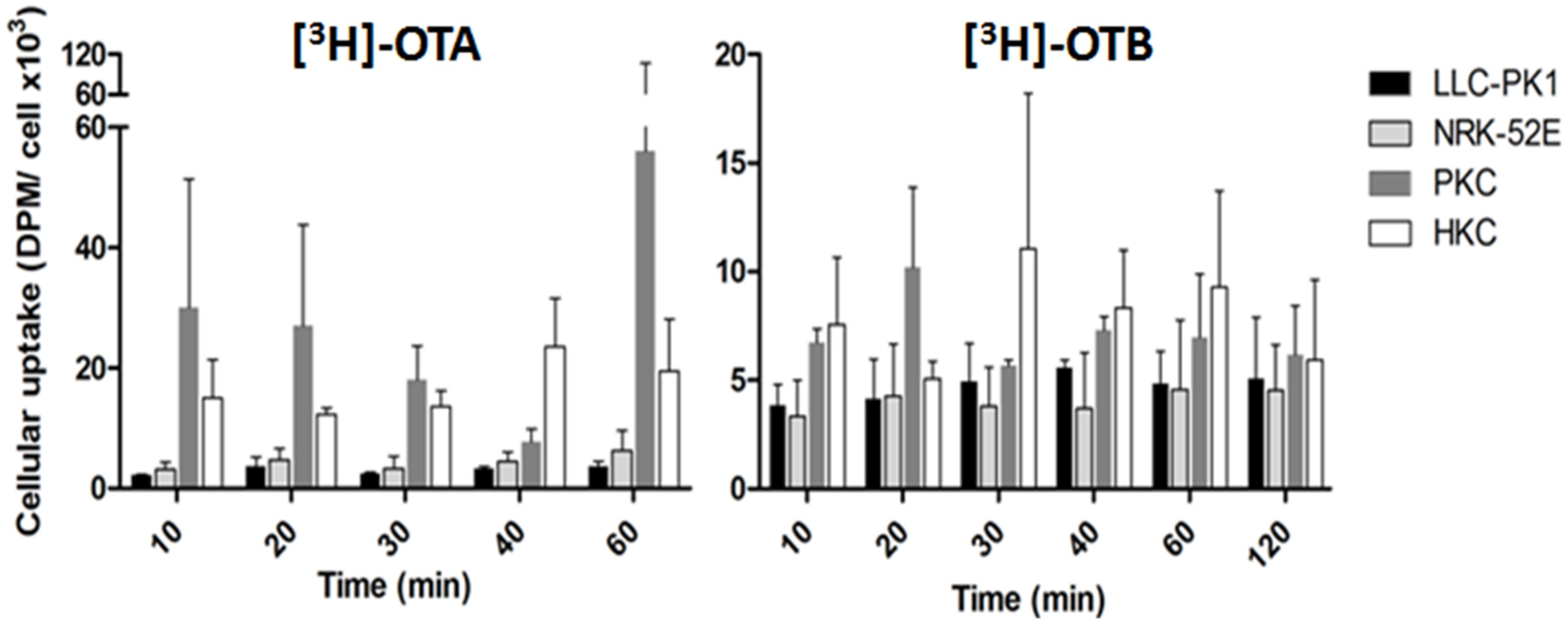
- O’Brien, E.; Heussner, A.H.; Dietrich, D.R. Species-, sex-, and cell type-specific effects of ochratoxin A and B. Toxicol. Sci. 2001, 63, 256–264. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.; Heussner, A.H.; Stack, M.E.; Thiel, R.; Dietrich, D.R. Species comparison of renal [3H]ochratoxin B protein binding and uptake in vitro. Toxicol. Sci. 2002, 66, 82. Available online: http://www.toxicology.org/pubs/docs/Tox/2002Tox.pdf (accessed on 20 October 2015). [Google Scholar]
- Stemmer, K.; Heussner, A.; Stack, M.; Thiel, R.; Dietrich, D. Comparison of ochratoxin binding to human, porcine and rat renal proteins in vitro. Toxicol. Sci. 2002, 66, 82. Available online: http://www.toxicology.org/pubs/docs/Tox/2002Tox.pdf (accessed on 20 October 2015). [Google Scholar]
- Schwerdt, G.; Freudinger, R.; Silbernagl, S.; Gekle, M. Ochratoxin A-binding proteins in rat organs and plasma and in different cell lines of the kidney. Toxicology 1999, 135, 1–10. [Google Scholar] [CrossRef]
- Heussner, A.H.; O’Brien, E.; Dietrich, D.R. Species- and sex-specific variations in binding of ochratoxin A by renal proteins in vitro. Exp. Toxicol. Pathol. 2002, 54, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; O’Brien, E.; Stack, M.E.; Thiel, R.; Dietrich, D.R. Cytotoxicity of OTA and OTB in human and porcine renal cells after repeated exposure. Toxicol. Sci. 2002, 66, 82. Available online: http://www.toxicology.org/pubs/docs/Tox/2002Tox.pdf (accessed on 20 October 2015). [Google Scholar]
- Heussner, A.H.; O’Brien, E.; Stack, M.E.; Dietrich, D.R. Alterations in human and porcine renal cells after repeated exposure to OTA and OTB. Toxicol. Sci. (Toxicologist) 2003, 72, 346–347. [Google Scholar]
- Grauenhorst, A.; Heussner, A.; Dietrich, D. Differential Expression Analysis of Kidney Cells Repeatedly Exposed to Ochratoxin. In Proceedings of the Joint Speciality Symposium on Renal Toxicology and Toxicologic Pathology, Lindau, Germany, 27 September–1 October 2004.
- Richetti, A.; Cavallaro, A.; Ainis, T.; Fimiani, V. Effect of mycotoxins on some activities of isolated human neutrophils. Immunopharmacol. Immunotoxicol. 2005, 27, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Richetti, A.; Cavallaro, A.; Ainis, T.; Fimiani, V. Effect of some mycotoxins on superoxide anion production of isolated human neutrophils and in whole blood. Immunopharmacol. Immunotoxicol. 2003, 25, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Lea, T.; Steien, K.; Størmer, F.C. Mechanism of ochratoxin A-induced immunosuppression. Mycopathologia 1989, 107, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Malaveille, C.; Brun, G.; Bartsch, H. Genotoxicity of ochratoxin A and structurally related compounds in Escherichia coli strains: Studies on their mode of action. IARC Sci. Publ. 1991, 115, 261–266. [Google Scholar] [PubMed]
- O’Brien, E.; Prietz, A.; Dietrich, D.R. Investigation of the teratogenic potential of ochratoxin A and B using the FETAX system. Birth Defects Res. B Dev. Reprod. Toxicol. 2005, 74, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Wiger, R.; Størmer, F.C. Effects of ochratoxins A and B on prechondrogenic mesenchymal cells from chick embryo limb buds. Toxicol. Lett. 1990, 54, 129–134. [Google Scholar] [CrossRef]
- Roth, A.; Creppy, E.E.; Kane, A.; Bacha, H.; Steyn, P.S.; Röschenthaler, R.; Dirheimer, G. Influence of ochratoxin B on the ochratoxin A inhibition of phenylalanyl-tRNA formation in vitro and protein synthesis in hepatoma tissue culture cells. Toxicol. Lett. 1989, 45, 307–313. [Google Scholar] [CrossRef]
- Chu, F.S.; Chang, C.C. Sensitivity of chicks to ochratoxins. J. Assoc. Off. Anal. Chem. 1971, 54, 1032–1034. [Google Scholar] [PubMed]
- Müller, G.; Burkert, B.; Rosner, H.; Köhler, H. Effects of the mycotoxin ochratoxin A and some of its metabolites on human kidney cell lines. Toxicol. Vitr. 2003, 17, 441–448. [Google Scholar] [CrossRef]
- Köhler, H.; Heller, M.; Erler, W.; Müller, G.; Rosner, H.; Gräfe, U. Effect of ochratoxin A and ochratoxin C on the monocyte and lymphocyte function. Mycotoxin Res. 2002, 18, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.; Rosner, H.; Rohrmann, B.; Erler, W.; Geschwend, G.; Gräfe, U.; Burkert, B.; Möller, U.; Diller, R.; Sachse, K.; et al. Effects of the mycotoxin ochratoxin A and some of its metabolites on the human cell line THP-1. Toxicology 2003, 184, 69–82. [Google Scholar] [CrossRef]
- Köhler, H.; Heller, M.; Erler, W.; Müller, G. Effects of a long-term exposure with OTA or OTC on functions of a human monocytic cell line. Mycotoxin Res. 2003, 19, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Steyn, P.S.; Holzapfel, C.W. The isolation of methyl and ethyl esters of ochratoxin A and B, metabolites of Aspergillus ochraceus Wilh. J. S. Afr. Chem. Inst. 1967, 20, 186–189. [Google Scholar]
- Creppy, E.E.; Størmer, F.C.; Kern, D.; Röschenthaler, R.; Dirheimer, G. Effects of ochratoxin A metabolites on yeast phenylalanyl-tRNA synthetase and on the growth and in vivo protein synthesis of hepatoma cells. Chem. Biol. Interact. 1983, 47, 239–247. [Google Scholar] [CrossRef]
- Bittner, A.; Cramer, B.; Harrer, H.; Humpf, H.U. Structure elucidation and in vitro cytotoxicity of ochratoxin α amide, a new degradation product of ochratoxin A. Mycotoxin Res. 2015, 31, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Foellmann, W.; Hillebrand, I.; Creppy, E.; Bolt, H. Sister chromatid exchange frequency in cultured isolated porcine urinary bladder epithelial cells (PUBEC) treated with ochratoxin A and α. Arch. Toxicol. 1995, 69, 280–286. [Google Scholar] [CrossRef]
- Hutchison, R.D.; Steyn, P.S. The isolation and structure of 4-hydroxyochratoxin and 7-carboxy-3,4-dihydro-8-hydroxy-3-methylisocoumarin from Penicillium viridicatum. Tetrahedron Lett. 1971, 43, 4033–4036. [Google Scholar] [CrossRef]
- Faucet-Marquis, V.; Pont, F.; Størmer, F.C.; Rizk, T.; Castegnaro, M.; Pfohl-Leszkowicz, A. Evidence of a new dechlorinated ochratoxin A derivative formed in opossum kidney cell cultures after pretreatment by modulators of glutathione pathways: Correlation with DNA-adduct formation. Mol. Nutr. Food Res. 2006, 50, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Gautier, J.; Richoz, J.; Welti, D.; Markovic, J.; Gremaud, E.; Guengerich, F.; Turesky, R. Metabolism of ochratoxin A: Absence of formation of genotoxic derivatives by human and rat enzymes. Chem. Res. Toxicol. 2001, 14, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E.; Kern, D.; Steyn, P.S.; Vleggaar, R.; Röschenthaler, R.; Dirheimer, G. Comparative study of the effect of ochratoxin A analogues on yeast aminoacyl-tRNA synthetases and on the growth and protein synthesis of hepatoma cells. Toxicol. Lett. 1983, 19, 217–224. [Google Scholar] [CrossRef]
- Richard, J. Some major mycotoxins and their mycotoxicoses—An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Bragulat, M.R.; Martínez, E.; Castellá, G.; Cabañes, F.J. Ochratoxin A and citrinin producing species of the genus Penicillium from feedstuffs. Int. J. Food Microbiol. 2008, 126, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Frank, J.M.; Houbraken, J.A.M.P.; Kuijpers, A.F.A.; Samson, R.A. New ochratoxin A producing species of Aspergillus section Circumdati. Stud. Mycol. 2004, 50, 23–43. [Google Scholar]
- Soares, C.; Calado, T.; Venâncio, A. Mycotoxin production by Aspergillus niger aggregate strains isolated from harvested maize in three Portuguese regions. Rev. Iberoam. Micol. 2013, 30, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Speijers, G.J.; Speijers, M.H. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Elsgaard, L.; Børgesen, C.; Olesen, J.; Siebert, S.; Ewert, F.; Peltonen-Sainio, P.; Rötter, R.; Skjelvåg, A. Shifts in comparative advantages for maize, oat and wheat cropping under climate change in Europe. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1514–1526. [Google Scholar] [CrossRef] [PubMed]
- Van der Fels-Klerx, H.; Olesen, J.; Naustvoll, L.; Friocourt, Y.; Mengelers, M.; Christensen, J. Climate change impacts on natural toxins in food production systems, exemplified by deoxynivalenol in wheat and diarrhetic shellfish toxins. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1647–1659. [Google Scholar] [CrossRef] [PubMed]
- Rumora, L.; Domijan, A.M.; Zanic Grubisic, T.; Segvic Klaric, M. Differential activation of MAPKs by individual and combined ochratoxin A and citrinin treatments in porcine kidney PK15 cells. Toxicon 2014, 90, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Connolly, L.; Frizzell, C.; Elliott, C.T. High content analysis: A sensitive tool to detect and quantify the cytotoxic, synergistic and antagonistic effects of chemical contaminants in foods. Toxicol. Lett. 2015, 233, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Föllmann, W.; Behm, C.; Degen, G.H. Toxicity of the mycotoxin citrinin and its metabolite dihydrocitrinone and of mixtures of citrinin and ochratoxin A in vitro. Arch. Toxicol. 2014, 88, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, B.; He, X.; Cheng, W.H.; Xu, W.; Luo, Y.; Liang, R.; Luo, H.; Huang, K. Analysis of individual and combined effects of ochratoxin A and zearalenone on HepG2 and KK-1 cells with mathematical models. Toxins 2014, 6, 1177–1192. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Connolly, L.; Frizzell, C.; Elliott, C.T. Cytotoxic assessment of the regulated, co-existing mycotoxins aflatoxin B1, fumonisin B1 and ochratoxin, in single, binary and tertiary mixtures. Toxicon 2014, 90, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Anninou, N.; Chatzaki, E.; Papachristou, F.; Pitiakoudis, M.; Simopoulos, C. Mycotoxins’ activity at toxic and sub-toxic concentrations: Differential cytotoxic and genotoxic effects of single and combined administration of sterigmatocystin, ochratoxin A and citrinin on the hepatocellular cancer cell line Hep3B. Int. J. Environ. Res. Public Health 2014, 11, 1855–1872. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Dietrich, D.R.; O’Brien, E. In vitro investigation of individual and combined cytotoxic effects of ochratoxin A and other selected mycotoxins on renal cells. Toxicol. Vitr. 2006, 20, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Bouslimi, A.; Bouaziz, C.; Ayed-Boussema, I.; Hassen, W.; Bacha, H. Individual and combined effects of ochratoxin A and citrinin on viability and DNA fragmentation in cultured Vero cells and on chromosome aberrations in mice bone marrow cells. Toxicology 2008, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bouslimi, A.; Ouannes, Z.; Golli, E.E.; Bouaziz, C.; Hassen, W.; Bacha, H. Cytotoxicity and oxidative damage in kidney cells exposed to the mycotoxins ochratoxin a and citrinin: Individual and combined effects. Toxicol. Mech. Methods 2008, 18, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.S.; Zelježić, D.; Rumora, L.; Peraica, M.; Pepeljnjak, S.; Domijan, A.M. A potential role of calcium in apoptosis and aberrant chromatin forms in porcine kidney PK15 cells induced by individual and combined ochratoxin A and citrinin. Arch. Toxicol. 2012, 86, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, L.; Dhivya, R.; Dhanasekaran, D.; Periasamy, V.S.; Alshatwi, A.A.; Akbarsha, M.A. Hepatotoxic effect of ochratoxin A and citrinin, alone and in combination, and protective effect of vitamin E: In vitro study in HepG2 cell. Food Chem. Toxicol. 2015, 83, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Corcuera, L.A.; Vettorazzi, A.; Arbillaga, L.; Perez, N.; Gil, A.G.; Azqueta, A.; Gonzalez-Penas, E.; Garcia-Jalon, J.A.; Lopez de Cerain, A. Genotoxicity of Aflatoxin B1 and Ochratoxin A after simultaneous application of the in vivo micronucleus and comet assay. Food Chem. Toxicol. 2015, 76, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Cano-Sancho, G.; Gonzalez-Arias, C.A.; Ramos, A.J.; Sanchis, V.; Fernandez-Cruz, M.L. Cytotoxicity of the mycotoxins deoxynivalenol and ochratoxin A on Caco-2 cell line in presence of resveratrol. Toxicol. Vitr. 2015, 29, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Segvic Klaric, M.; Jaksic Despot, D.; Kopjar, N.; Rasic, D.; Kocsube, S.; Varga, J.; Peraica, M. Cytotoxic and genotoxic potencies of single and combined spore extracts of airborne OTA-producing and OTA-non-producing Aspergilli in Human lung A549 cells. Ecotoxicol. Environ. Saf. 2015, 120, 206–214. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heussner, A.H.; Bingle, L.E.H. Comparative Ochratoxin Toxicity: A Review of the Available Data. Toxins 2015, 7, 4253-4282. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7104253
Heussner AH, Bingle LEH. Comparative Ochratoxin Toxicity: A Review of the Available Data. Toxins. 2015; 7(10):4253-4282. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7104253
Chicago/Turabian StyleHeussner, Alexandra H., and Lewis E. H. Bingle. 2015. "Comparative Ochratoxin Toxicity: A Review of the Available Data" Toxins 7, no. 10: 4253-4282. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7104253