
Botulinum Toxin Type A Induces Changes in the Chemical Coding of Substance P-Immunoreactive Dorsal Root Ganglia Sensory Neurons Supplying the Porcine Urinary Bladder

Abstract
:1. Introduction
2. Results and Discussion
2.1. Distribution of SP-Containing BPSN Neurons in Control and BTX-Treated Animals

2.2. Immunohistochemical Characteristic of SP-Containing BPSN Neurons in Control and BTX-Treated Animals




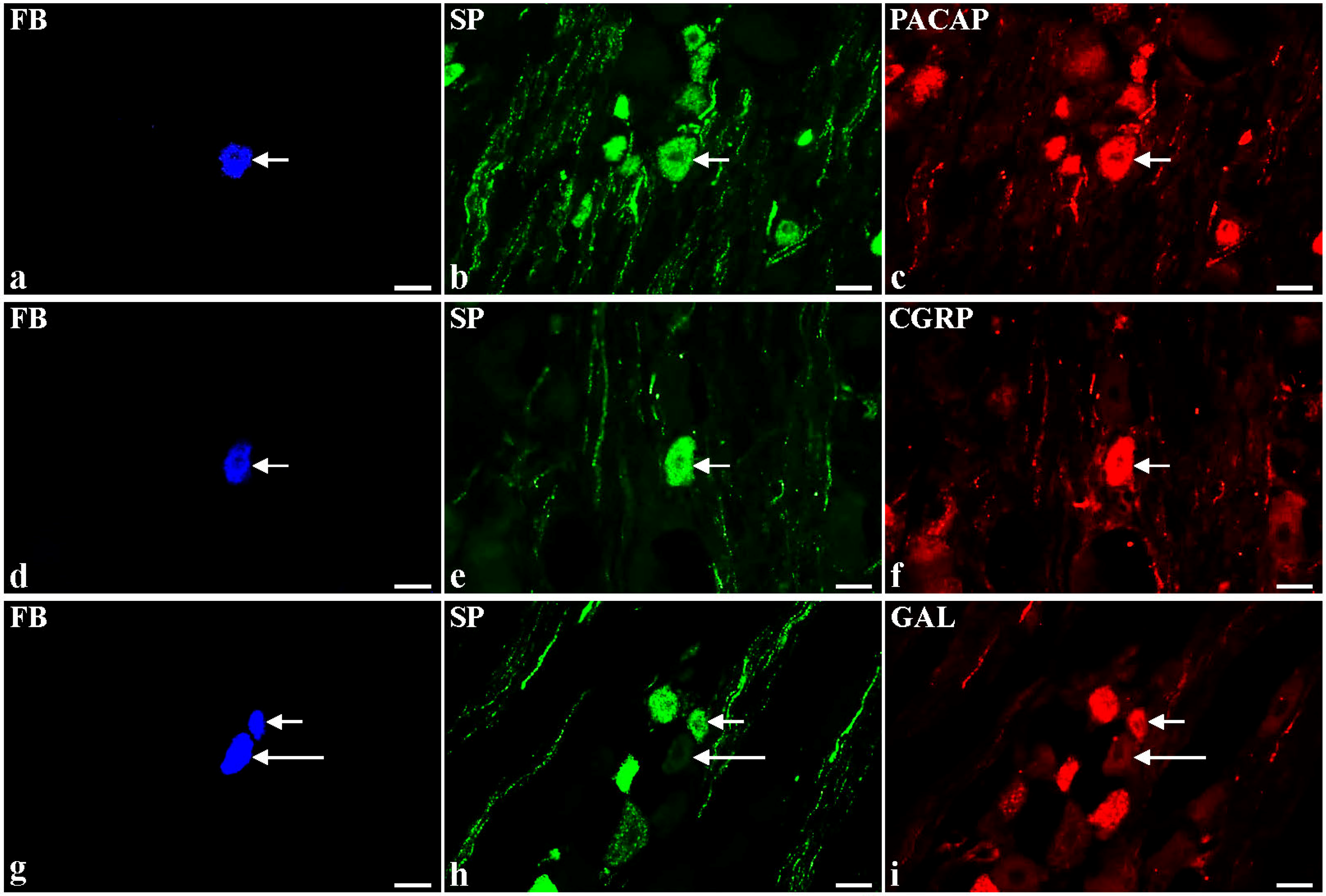{kind=link}
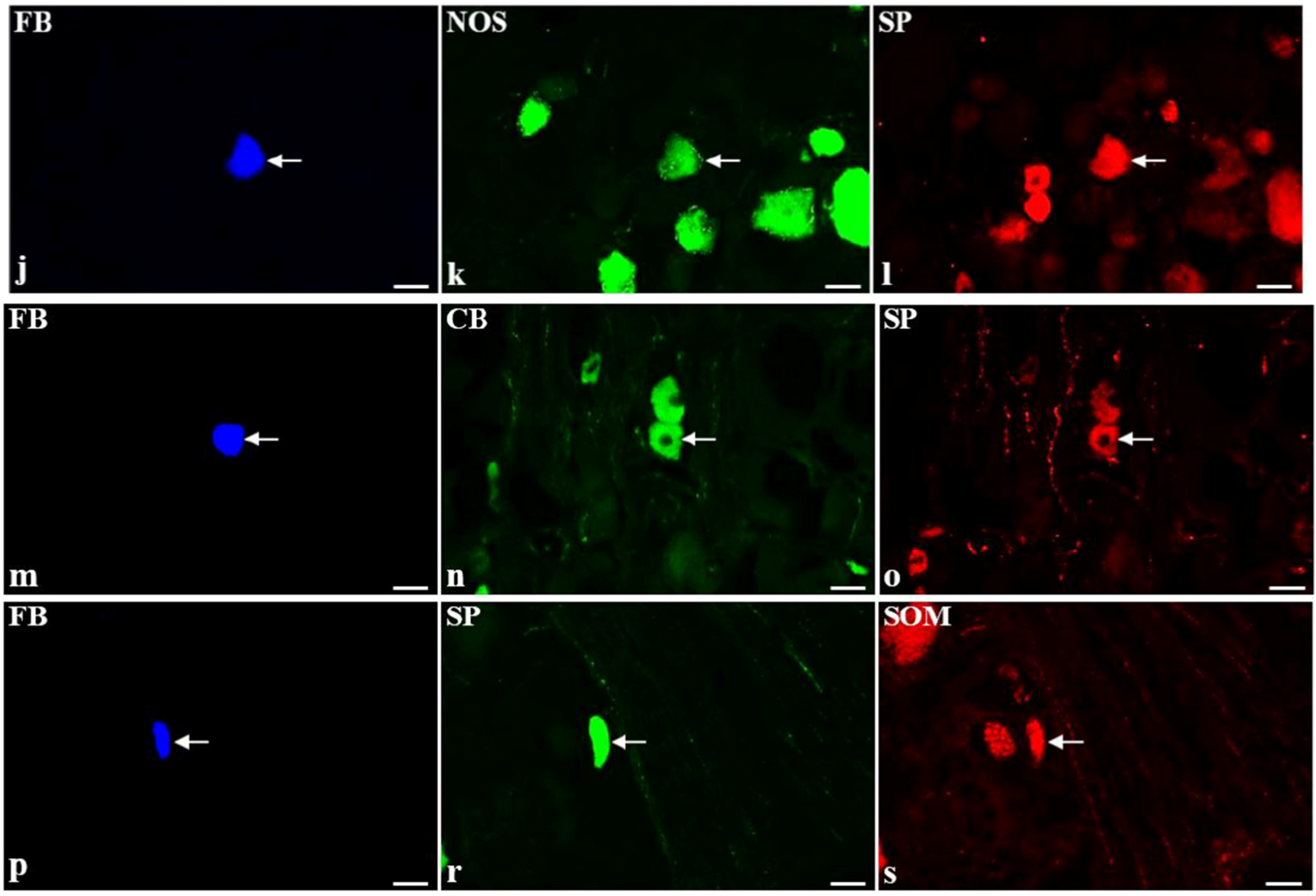{kind=link}
{kind=link}
{kind=link}
| Collocation Patterns of SP with Different Neurotransmitters in the Bladder DRG Neurons | The Group of Control Animals | The Group of BTX-Treated Animals |
|---|---|---|
| SP+/CGRP+/PACAP+ | 11.0% ± 5.1% | 10.9% ± 1.3% |
| SP+/PACAP+/GAL+ | 0.8% ± 0.4% | **13.3% ± 3.5% |
| SP+/PACAP+/nNOS+ | 0.7% ± 0.2% | 1.2% ± 0.4% |
| SP+/CGRP+/GAL+ | 0.5% ± 0.5% | 0% |
| SP+/CGRP+/SOM+ | 0.4% ± 0.4% | 0% |
| SP+/PACAP+/CB+ | 0.3% ± 0.4% | 0% |
| SP+/CGRP+/PACAP+/GAL+ | 1.6% ± 0.9% | 3.1% ± 0.7% |
| SP+/PACAP+/GAL+/nNOS+ | 0.4% ± 0.3% | 0% |
| SP+/CGRP+/PACAP+/nNOS+ | 0.2% ± 0.2% | 0% |
| SP+/CGRP+/PACAP+/SOM+/GAL+ | 0.5% ± 0.4% | 0.6% ± 0.3% |
| SP+/CGRP+/PACAP+/SOM+/GAL+/CB+ | 0.3% ± 0.3% | 0% |
3. Discussion
4. Experimental Section
4.1. Laboratory Animals
4.2. Surgical Procedures
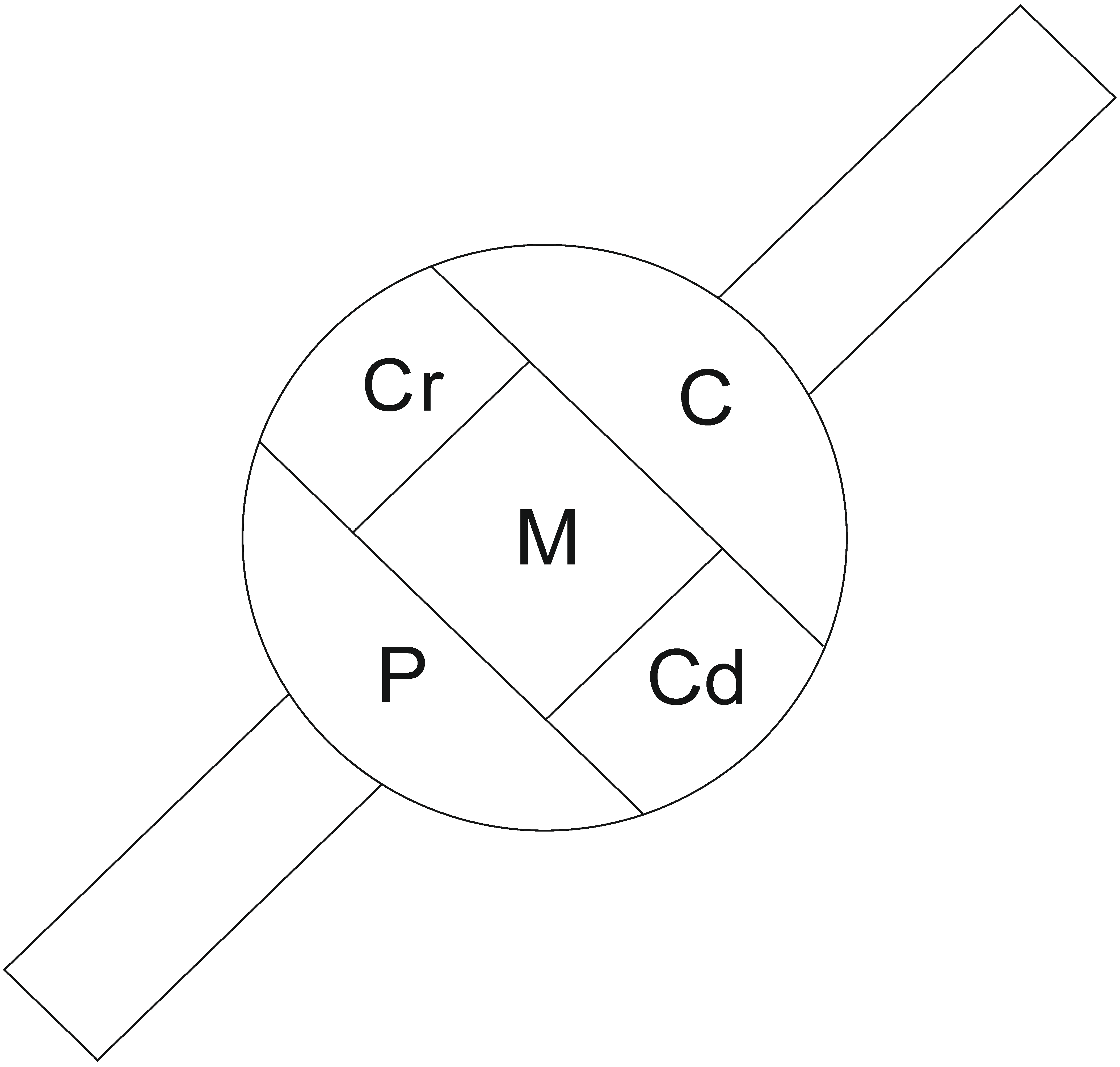
4.3. Sectioning of the Ganglia and Estimation of the Total Number of the DRG—BPSN
4.4. Immunohistochemical Procedure
4.5. Control of Specificity of the Tracer Staining and Immunohistochemical Procedures
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Van Ermengem, E. Ṻber einen neuen anaeroben Bacillus and seine Beziehungen zum Botulismus. Z. Hyg. Infekt. 1897, 26, 1–56. [Google Scholar]
- Haferkamp, A.; Schurch, B.; Reitz, A.; Krengel, U.; Grosse, J.; Kramer, G.; Schumacher, S.; Bastian, P.J.; Buttner, R.; Muller, S.C.; et al. Lack of ultrastructural detrusor changes following endoscopic injection of botulinum toxin type A in overactive neurogenic bladder. Eur. Urol. 2004, 46, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Chancellor, M.B.; Fowler, C.J.; Apostolidis, A.; de Groat, W.C.; Smith, C.P.; Somogyi, G.T.; Aoki, K.R. Drug Insight: Biological effects of botulinum toxin A in the lower urinary tract. Nat. Clin. Pract. Urol. 2008, 5, 319–328. [Google Scholar] [PubMed]
- Jankovic, J. Botulinum toxin in clinical practice. J. Neurol. Neurosurg. Psychiatr. 2004, 75, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.P.; Franks, M.E.; McNeil, B.K.; Ghosh, R.; de Groat, W.C.; Chancellor, M.B.; Somogyi, G.T. Effect of botulinum toxin A on the autonomic nervous system of the rat lower urinary tract. J. Urol. 2003, 169, 1896–1900. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, J.; Carruthers, A. Botox: Beyond wrinkles. Clin. Dermatol. 2004, 22, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Lowe, N.J.; Lowe, P. Botulinum toxins for facial lines: A concise review. Dermatol. Ther. (Heidelb.) 2012, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Filippi, G.M.; Errico, P.; Santarelli, R.; Bagolini, B.; Manni, E. Botulinum A toxin effects on rat jaw muscle spindles. Acta Otolaryngol. 1993, 113, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Welch, M.J.; Purkiss, J.R.; Foster, K.A. Sensitivity of embryonic rat dorsal root ganglia neurons to Clostridium botulinum neurotoxins. Toxicon 2000, 38, 245–258. [Google Scholar] [CrossRef]
- Durham, P.L.; Cady, R. Regulation of calcitonin gene-related peptide secretion from trigeminal nerve cells by botulinum toxin type A: Implications for migraine therapy. Headache 2004, 44, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Wang, J.; Lawrence, G.; Dolly, J.O. Synaptobrevin I mediates exocytosis of CGRP from sensory neurons and inhibition by botulinum toxins reflects their anti-nociceptive potential. J. Cell. Sci. 2007, 120, 2864–2874. [Google Scholar] [CrossRef] [PubMed]
- Dolly, J.O.; O’Connell, M.A. Neurotherapeutics to inhibit exocytosis from sensory neurons for the control of chronic pain. Curr. Opin. Pharmacol. 2012, 12, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Khanijou, S.; Rubino, J.; Aoki, K.R. Subcutaneous administration of botulinum toxin A reduces formalin-induced pain. Pain 2004, 107, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Coggeshall, R.E.; Carlton, S.M. Ultrastructural analysis of NMDA, AMPA, and kainite receptors on unmyelinated and myelinated axons in the periphery. J. Comp. Neurol. 1998, 391, 78–86. [Google Scholar] [CrossRef]
- Jabbari, B.; Maher, N.; Difazio, M.P. Botulinum toxin A improved burning pain and allodynia in two patients with spinal cord pathology. Pain Med. 2003, 4, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Neugart, F.; Groh, R.; Gotz, T.; Horsch, R. Injections of botulinum toxin A into the detrusor vesicae for treatment of refractory detrusor hyperactivity in non-neurological patients. Aktuelle Urol. 2006, 37, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Liao, C.H.; Kuo, H.C. Therapeutic effects of detrusor botulinum toxin A injection on neurogenic detrusor overactivity in patients with different levels of spinal cord injury and types of detrusor sphincter dyssynergia. Spinal Cord 2011, 49, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Kanagarajah, P.; Ayyathurai, R.; Caruso, D.J.; Gomez, C.; Gousse, A.E. Role of botulinum toxin-A in refractory idiopathic overactive bladder patients without detrusor overactivity. Int. Urol. Nephrol. 2012, 44, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Cruz, F.; Herschorn, S.; Aliotta, P.; Brin, M.; Thompson, C.; Lam, W.; Daniell, G.; Heesakkers, J.; Haag-Molkenteller, C. Efficacy and safety of onabotulinumtoxin A in patients with urinary incontinence due to neurogenic detrusor overactivity: A randomized, double-blind, placebo-controlled trial. Eur. Urol. 2011, 60, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Rohrsted, M.; Nordsten, C.B.; Bagi, P. Onabotulinum toxin a (Botox®) in the treatment of neurogenic bladder overactivity. Nephrol. Urol. Mon. 2012, 4, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, A.; da Silva, C.M.; Cruz, F. Botulinum toxin treatment for bladder dysfunction. Int. J. Urol. 2013, 20, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Gottsch, H.P.; Miller, J.L.; Yang, C.C.; Berger, R.E. A pilot study of botulinum toxin for interstitial cystitis/painful bladder syndrome. Neurourol. Urodyn. 2011, 30, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.; Lopes, T.; Silva, J.; Silva, C.; Dinis, P.; Cruz, F. Persistent therapeutic effect of repeated injections of onabotulinum toxin A in refractory bladder pain syndrome/interstitial cystitis. J. Urol. 2013, 189, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Lepiarczyk, E.; Bossowska, A.; Kaleczyc, J.; Majewski, M. The influence of botulinum toxin type A (BTX) on the immunohistochemical characteristics of noradrenergic and cholinergic nerve fibers supplying the porcine urinary bladder wall. Pol. J. Vet. Sci. 2011, 14, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.P.; Radziszewski, P.; Borkowski, A.; Somogyi, G.T.; Boone, T.B.; Chancellor, M.B. Botulinum toxin A has antinociceptive effects in treating interstitial cystitis. Urology 2004, 64, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.C.; Yoshimura, N.; Huang, C.C.; Chiang, P.H.; Chancellor, M.B. Intravesical botulinum toxin A administration produces analgesia against acetic acid induced bladder pain responses in rats. J. Urol. 2004, 172, 1529–1532. [Google Scholar] [CrossRef] [PubMed]
- Reitz, A.; Stohrer, M.; Kramer, G.; Del, P.G.; Chartier-Kastler, E.; Pannek, J.; Burgdorfer, H.; Gocking, K.; Madersbacher, H.; Schumacher, S.; et al. European experience of 200 cases treated with botulinum-A toxin injections into the detrusor muscle for urinary incontinence due to neurogenic detrusor overactivity. Eur. Urol. 2004, 45, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, A.; Popat, R.; Yiangou, Y.; Cockayne, D.; Ford, A.P.; Davis, J.B.; Dasgupta, P.; Fowler, C.J.; Anand, P. Decreased sensory receptors P2X3 and TRPV1 in suburothelial nerve fibers following intradetrusor injections of botulinum toxin for human detrusor overactivity. J. Urol. 2005, 174, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Morenilla-Palao, C.; Planells-Cases, R.; García-Sanz, N.; Ferrer-Montiel, A. Regulated exocytosis contributes to protein kinase C potentiation of vanilloid receptor activity. J. Biol. Chem. 2004, 279, 25665–25672. [Google Scholar] [CrossRef] [PubMed]
- Bossowska, A.; Majewski, M. Botulinum toxin type A-induced changes in the chemical coding of dorsal root ganglion neurons supplying the porcine urinary bladder. Pol. J. Vet. Sci. 2012, 15, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Mersdorf, A.; Schmidt, R.A.; Kaula, N.; Tanagho, E.A. Intrathecal administration of substance P in the rat: The effect on bladder and urethral sphincteric activity. Urology 1992, 40, 87–96. [Google Scholar] [CrossRef]
- Maggi, C.A. Tachykinins as peripheral modulators of primary afferent nerves and visceral sensitivity. Pharmacol. Res. 1997, 36, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Vizzard, M.A. Alterations in neuropeptide expression in lumbosacral bladder pathways following chronic cystitis. J. Chem. Neuroanat. 2001, 21, 125–138. [Google Scholar] [CrossRef]
- Callsen-Cencic, P.; Mense, S. Expression of neuropeptides and nitric oxide synthase in neurons innervating the inflamed rat urinary bladder. J. Auton. Nerv. Syst. 1997, 65, 33–44. [Google Scholar] [CrossRef]
- Duggan, A.W.; Riley, R.C.; Mark, M.A.; MacMillan, S.J.; Schaible, H.G. Afferent volley patterns and the spinal release of immunoreactive substance P in the dorsal horn of the anaesthetized spinal cat. Neuroscience 1995, 65, 849–858. [Google Scholar] [CrossRef]
- Chien, C.T.; Yu, H.J.; Lin, T.B.; Lai, M.K.; Hsu, S.M. Substance P via NK1 receptor facilitates hyperactive bladder afferent signaling via action of ROS. Am. J. Physiol. Renal Physiol. 2003, 284, F840–F851. [Google Scholar] [CrossRef] [PubMed]
- Terai, A.; Matsui, Y.; Ichioka, K.; Ohara, H.; Terada, N.; Yoshimura, K. Comparative analysis of lower urinary tract symptoms and bother in both sexes. Urology 2004, 63, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Dalmose, A.L.; Hvistendahl, J.J.; Olsen, L.H.; Eskild-Jensen, A.; Djurhuus, J.C.; Swindle, M.M. Surgically induced urologic models in swine. J. Investig. Surg. 2000, 13, 133–145. [Google Scholar] [CrossRef]
- Kuzmuk, K.N.; Schook, L.B. Pigs as a model for biomedical sciences. In The Genetics of the Pig, 2nd ed.; Ruvinsky, R., Ed.; CAB International: Oxford Shire, UK, 2011; pp. 426–444. [Google Scholar]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Bossowska, A.; Crayton, R.; Radziszewski, P.; Kmieć, Z.; Majewski, M. Distribution and neurochemical characterization of sensory dorsal root ganglia neurons supplying porcine urinary bladder. J. Physiol. Pharmacol. 2009, 60, 77–81. [Google Scholar] [PubMed]
- Russo, D.; Clavenzani, P.; Sorteni, C.; Bo Minelli, L.; Botti, M.; Gazza, F.; Panu, R.; Ragionieri, L.; Chiocchetti, R. Neurochemical features of boar lumbosacral dorsal root ganglion neurons and characterization of sensory neurons innervating the urinary bladder trigone. J. Comp. Neurol. 2013, 521, 342–366. [Google Scholar] [CrossRef] [PubMed]
- Giannantoni, A.; Di Stasi, S.M.; Nardicchi, V.; Zucchi, A.; Macchioni, L.; Bini, V.; Goracci, G.; Porena, M. Botulinum-A toxin injections into the detrusor muscle decrease nerve growth factor bladder tissue levels in patients with neurogenic detrusor overactivity. J. Urol. 2006, 175, 2341–2344. [Google Scholar] [CrossRef]
- Averill, S.; McMahon, S.B.; Clary, D.O.; Reichardt, L.F.; Priestley, J.V. Immunocytochemical localization of trkA receptors in chemically identified subgroups of adult rat sensory neurons. Eur. J. Neurosci. 1995, 7, 1484–1494. [Google Scholar] [CrossRef] [PubMed]
- Charbel-Issa, P.; Lever, I.J.; Michael, G.J.; Bradbury, E.J.; Malcangio, M. Intrathecally delivered glial cell line-derived neurotrophic factor produces electrically evoked release of somatostatin in the dorsal horn of the spinal cord. J. Neurochem. 2001, 78, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, G.W.; Ovsepian, S.V.; Wang, J.; Aoki, K.R.; Dolly, J.O. Extravesicular intraneuronal migration of internalized botulinum neurotoxins without detectable inhibition of distal neurotransmission. Biochem. J. 2012, 441, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L. The life history of a botulinum toxin molecule. Toxicon 2013, 68, 40–59. [Google Scholar] [CrossRef] [PubMed]
- Papagiannopoulou, D.; Vardouli, L.; Dimitriadis, F.; Apostolidis, A. Retrograde transport of radiolabeled botulinum neurotoxin type A to the CNS after intradetrusor injection in rats. BJU Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lucioni, A.; Bales, G.T.; Lotan, T.L.; McGehee, D.S.; Cook, S.P.; Rapp, D.E. Botulinum toxin type A inhibits sensory neuropeptide release in rat bladder models of acute injury and chronic inflammation. BJU Int. 2008, 101, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Sukiennik, A.; Carr, D.B.; Bonney, I.; Marchand, J.E.; Wurm, H.; Sant, G.R. The effect of short-term epidural local anesthetic blockade on urinary levels of substance P in interstitial cystitis. Anesth. Analg. 2004, 98, 846–850. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, O.; Alm, P.; Larsson, B.; Mattiasson, A.; Andersson, K.E. Facilitatory effect of pituitary adenylate cyclase activating polypeptide on micturition in normal, conscious rats. Neuroscience 1995, 66, 1009–1014. [Google Scholar] [CrossRef]
- Braas, K.M.; May, V.; Zvara, P.; Nausch, B.; Kliment, J.; Dunleavy, J.D.; Nelson, M.T.; Vizzard, M.A. Role for pituitary adenylate cyclase activating polypeptide in cystitis-induced plasticity of micturition reflexes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R951–R962. [Google Scholar] [CrossRef] [PubMed]
- Girard, B.M.; Wolf-Johnston, A.; Braas, K.M.; Birder, L.A.; May, V.; Vizzard, M.A. PACAP-mediated ATP release from rat urothelium and regulation of PACAP/VIP and receptor mRNA in micturition pathways after cyclophosphamide (CYP)-induced cystitis. J. Mol. Neurosci. 2008, 36, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, M.; Brailoiu, G.C.; Shiraki, M.; Dun, N.J.; Paul, K.; Tseng, L.F. Modulation of nociceptive transmission by pituitary adenylate cyclase activating polypeptide in the spinal cord of the mouse. Pain 2002, 100, 27–34. [Google Scholar] [CrossRef]
- Mabuchi, T.; Matsumura, S.; Okuda-Ashitaka, E.; Kitano, T.; Kojima, H.; Nagano, T.; Minami, T.; Ito, S. Attenuation of neuropathic pain by the nociceptin/orphanin FQ antagonist JTC-801 is mediated by inhibition of nitric oxide production. Eur. J. Neurosci. 2003, 17, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- Vizzard, M.A. Up-regulation of pituitary adenylate cyclase-activating polypeptide in urinary bladder pathways after chronic cystitis. J. Comp. Neurol. 2000, 420, 335–348. [Google Scholar] [CrossRef]
- Kim, W.K.; Kan, Y.; Ganea, D.; Hart, R.P.; Gozes, I.; Jonakait, G.M. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide inhibit tumor necrosis factor-alpha production in injured spinal cord and in activated microglia via a cAMP-dependent pathway. J. Neurosci. 2000, 20, 3622–3630. [Google Scholar] [PubMed]
- Zvarova, K.; Dunleavy, J.D.; Vizzard, M.A. Changes in pituitary adenylate cyclase activating polypeptide expression in urinary bladder pathways after spinal cord injury. Exp. Neurol. 2005, 192, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Andrade, J.M.; Zhou, S.; Du, J.; Yamani, A.; Grady, J.J.; Castaneda-Hernandez, G.; Carlton, S.M. Pro-nociceptive role of peripheral galanin in inflammatory pain. Pain 2004, 110, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Maggi, C.A. The dual function of capsaicin-sensitive sensory nerves in the bladder and urethra. Ciba Found. Symp. 1990, 151, 77–83. [Google Scholar] [PubMed]
- Biella, G.; Panara, C.; Pecile, A.; Sotgiu, M.L. Facilitatory role of calcitonin gene-related peptide (CGRP) on excitation induced by substance P (SP) and noxious stimuli in rat spinal dorsal horn neurons. An iontophoretic study in vivo. Brain Res. 1991, 559, 352–356. [Google Scholar] [CrossRef]
- Le Greves, P.; Nyberg, F.; Terenius, L.; Hokfelt, T. Calcitonin gene-related peptide is a potent inhibitor of substance P degradation. Eur. J. Pharmacol. 1985, 115, 309–311. [Google Scholar] [CrossRef]
- Gamse, R.; Saria, A. Potentiation of tachykinin-induced plasma protein extravasation by calcitonin gene-related peptide. Eur. J. Pharmacol. 1985, 114, 61–66. [Google Scholar] [CrossRef]
- Rapp, D.E.; Turk, K.W.; Bales, G.T.; Cook, S.P. Botulinum toxin type A inhibits calcitonin gene-related peptide release from isolated rat bladder. J. Urol. 2006, 175, 1138–1142. [Google Scholar] [CrossRef]
- Meng, J.; Ovsepian, S.V.; Wang, J.; Pickering, M.; Sasse, A.; Aoki, K.R.; Lawrence, G.W.; Dolly, J.O. Activation of TRPV1 mediates calcitonin gene-related peptide release, which excites trigeminal sensory neurons and is attenuated by a retargeted botulinum toxin with anti-nociceptive potential. J. Neurosci. 2009, 29, 4981–4992. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.P.; Khan, I.; Suhail, M.S.; Malkmus, S.; Yaksh, T.L. Spinal botulinum neurotoxin B: Effects on afferent transmitter release and nociceptive processing. PLoS ONE 2011, 29. [Google Scholar] [CrossRef]
- Colvin, L.A.; Mark, M.A.; Duggan, A.W. The effect of a peripheral mononeuropathy on immunoreactive (ir)-galanin release in the spinal cord of the rat. Brain Res. 1997, 766, 259–261. [Google Scholar] [CrossRef]
- Liu, H.X.; Hokfelt, T. The participation of galanin in pain processing at the spinal level. Trends Pharmacol. Sci. 2002, 23, 468–474. [Google Scholar] [CrossRef]
- Zvarova, K.; Murray, E.; Vizzard, M.A. Changes in galanin immunoreactivity in rat lumbosacral spinal cord and dorsal root ganglia after spinal cord injury. J. Comp. Neurol. 2004, 475, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.J.; Wiesenfeld-Hallin, Z.; Villar, M.J.; Hokfelt, T. Intrathecal galanin antagonizes the facilitatory effect of substance P on the nociceptive flexor reflex in the rat. Acta Physiol. Scand. 1989, 137, 463–464. [Google Scholar] [CrossRef] [PubMed]
- Meller, S.T.; Gebhart, G.F. Nitric oxide (NO) and nociceptive processing in the spinal cord. Pain 1993, 52, 127–136. [Google Scholar] [CrossRef]
- Kakizaki, H.; de Groat, W.C. Role of spinal nitric oxide in the facilitation of the micturition reflex by bladder irritation. J. Urol. 1996, 155, 355–360. [Google Scholar] [CrossRef]
- Aley, K.O.; Mc Carter, G.; Levine, J.D. Nitric oxide signaling in pain and nociceptor sensitization in the rat. J. Neurosci. 1998, 18, 7008–7014. [Google Scholar] [PubMed]
- Birder, L.A.; Kanai, A.J.; de Groat, W.C.; Kiss, S.; Nealen, M.L.; Burke, N.E.; Dineley, K.E.; Watkins, S.; Reynolds, I.J.; Caterina, M.J. Vanilloid receptor expression suggests a sensory role for urinary bladder epithelial cells. Proc. Natl Acad. Sci. USA 2001, 98, 13396–13401. [Google Scholar] [CrossRef] [PubMed]
- Sandkuhler, J.; Fu, Q.G.; Helmchen, C. Spinal somatostatin superfusion in vivo affects activity of cat nociceptive dorsal horn neurons: Comparison with spinal morphine. Neuroscience 1990, 34, 565–576. [Google Scholar] [CrossRef]
- Helyes, Z.; Than, M.; Oroszi, G.; Pinter, E.; Nemeth, J.; Keri, G.; Szolcsanyi, J. Anti-nociceptive effect induced by somatostatin released from sensory nerve terminals and by synthetic somatostatin analogues in the rat. Neurosci. Lett 2000, 278, 185–188. [Google Scholar] [CrossRef]
- Than, M.; Nemeth, J.; Szilvassy, Z.; Pinter, E.; Helyes, Z.; Szolcsanyi, J. Systemic anti-inflammatory effect of somatostatin released from capsaicin-sensitive vagal and sciatic sensory fibres of the rat and guinea-pig. Eur. J. Pharmacol. 2000, 399, 251–258. [Google Scholar] [CrossRef]
- Baimbridge, K.G.; Celio, M.R.; Rogers, J.H. Calcium-binding proteins in the nervous system. Trends Neurosci. 1992, 15, 303–308. [Google Scholar] [CrossRef]
- Li, Y.N.; Sakamoto, H.; Kawat, T.; Cheng, C.X.; Li, Y.C.; Shimada, O.; Atsumi, S. An immunocytochemical study of calbindin-D28K in laminae I and II of the dorsal horn and spinal ganglia in the chicken with special reference to the relation to substance P-containing primary afferent neurons. Arch. Histol. Cytol. 2005, 68, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Lepiarczyk, E.; Bossowska, A.; Majewski, M. Changes in chemical coding of sympathetic chain ganglia (SChG) neurons supplying porcine urinary bladder after botulinum toxin (BTX) treatment. Cell. Tissue Res. 2015, 360, 263–272. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bossowska, A.; Lepiarczyk, E.; Mazur, U.; Janikiewicz, P.; Markiewicz, W. Botulinum Toxin Type A Induces Changes in the Chemical Coding of Substance P-Immunoreactive Dorsal Root Ganglia Sensory Neurons Supplying the Porcine Urinary Bladder. Toxins 2015, 7, 4797-4816. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7114797
Bossowska A, Lepiarczyk E, Mazur U, Janikiewicz P, Markiewicz W. Botulinum Toxin Type A Induces Changes in the Chemical Coding of Substance P-Immunoreactive Dorsal Root Ganglia Sensory Neurons Supplying the Porcine Urinary Bladder. Toxins. 2015; 7(11):4797-4816. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7114797
Chicago/Turabian StyleBossowska, Agnieszka, Ewa Lepiarczyk, Urszula Mazur, Paweł Janikiewicz, and Włodzimierz Markiewicz. 2015. "Botulinum Toxin Type A Induces Changes in the Chemical Coding of Substance P-Immunoreactive Dorsal Root Ganglia Sensory Neurons Supplying the Porcine Urinary Bladder" Toxins 7, no. 11: 4797-4816. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7114797