
Biodegradation of Ochratoxin A by Bacterial Strains Isolated from Vineyard Soils

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
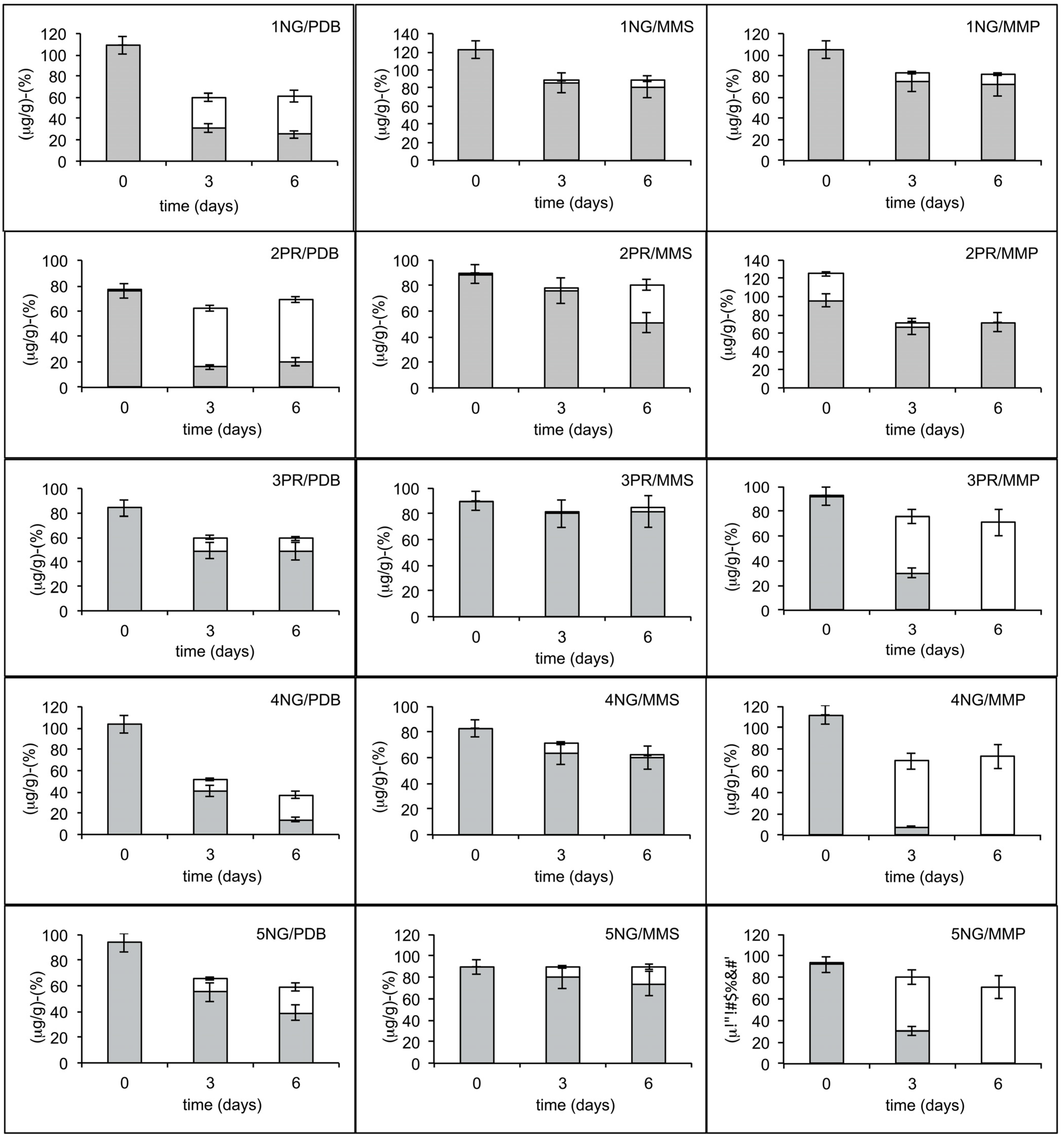
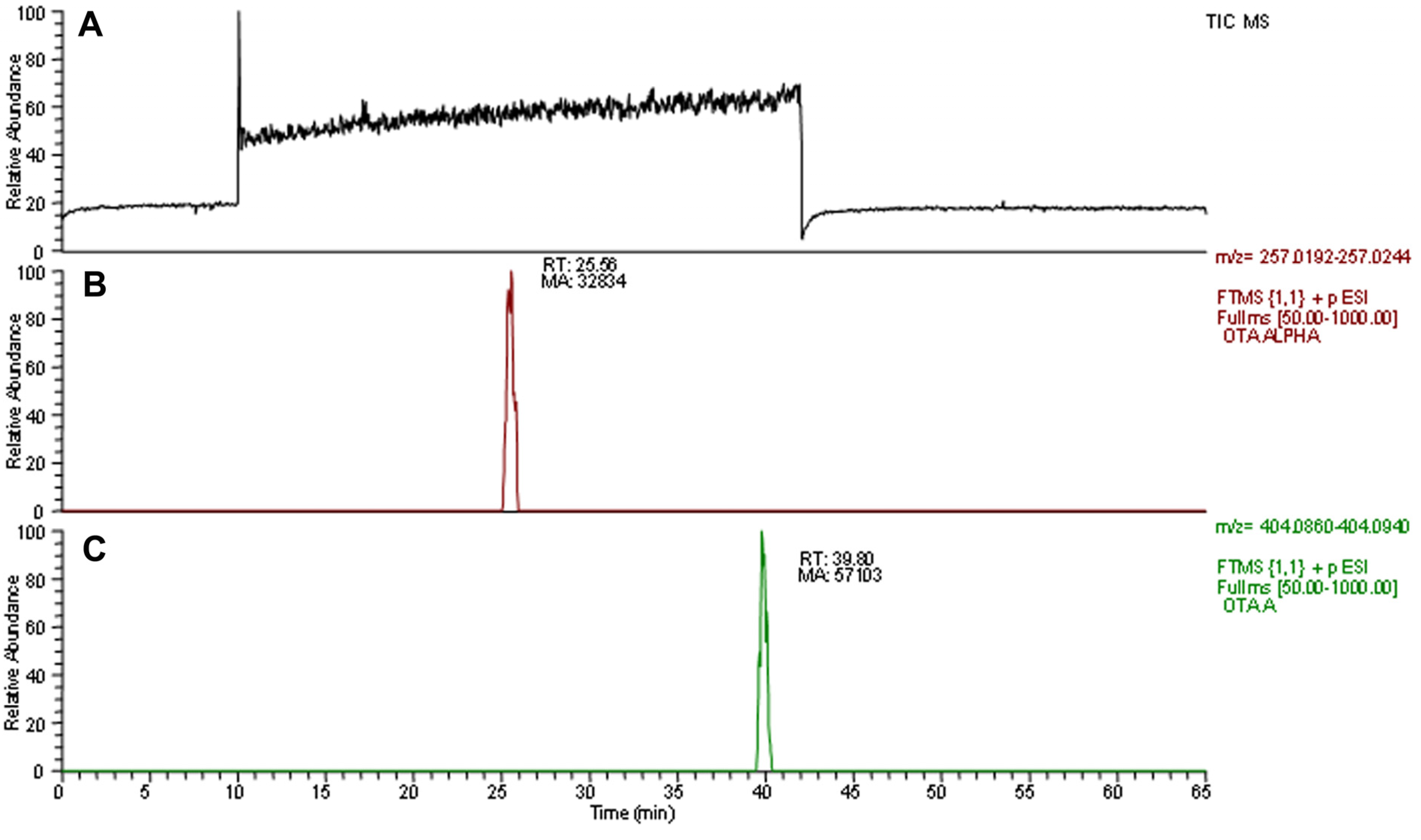
2.1. Degradation Activity of Soil and Branch Samples



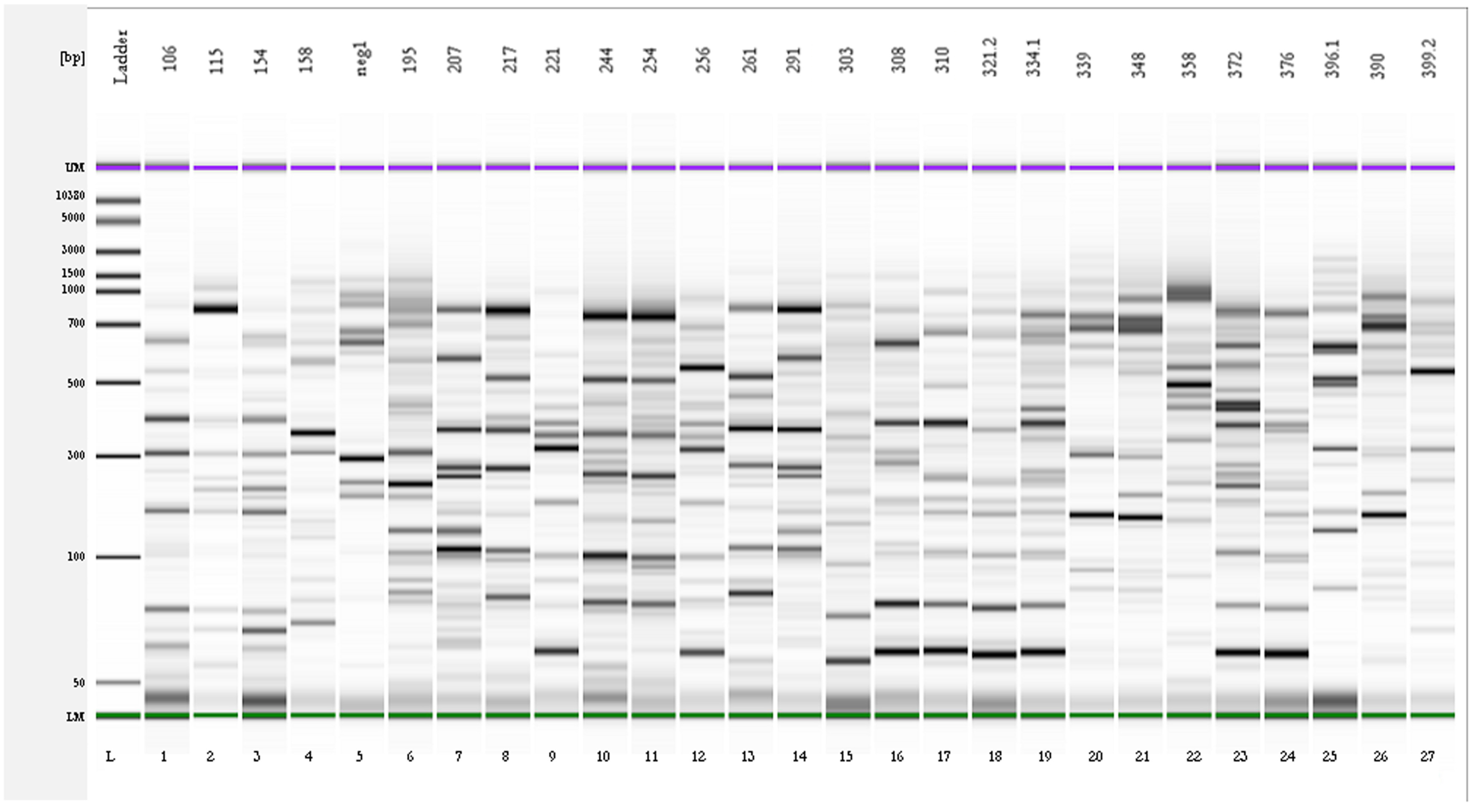
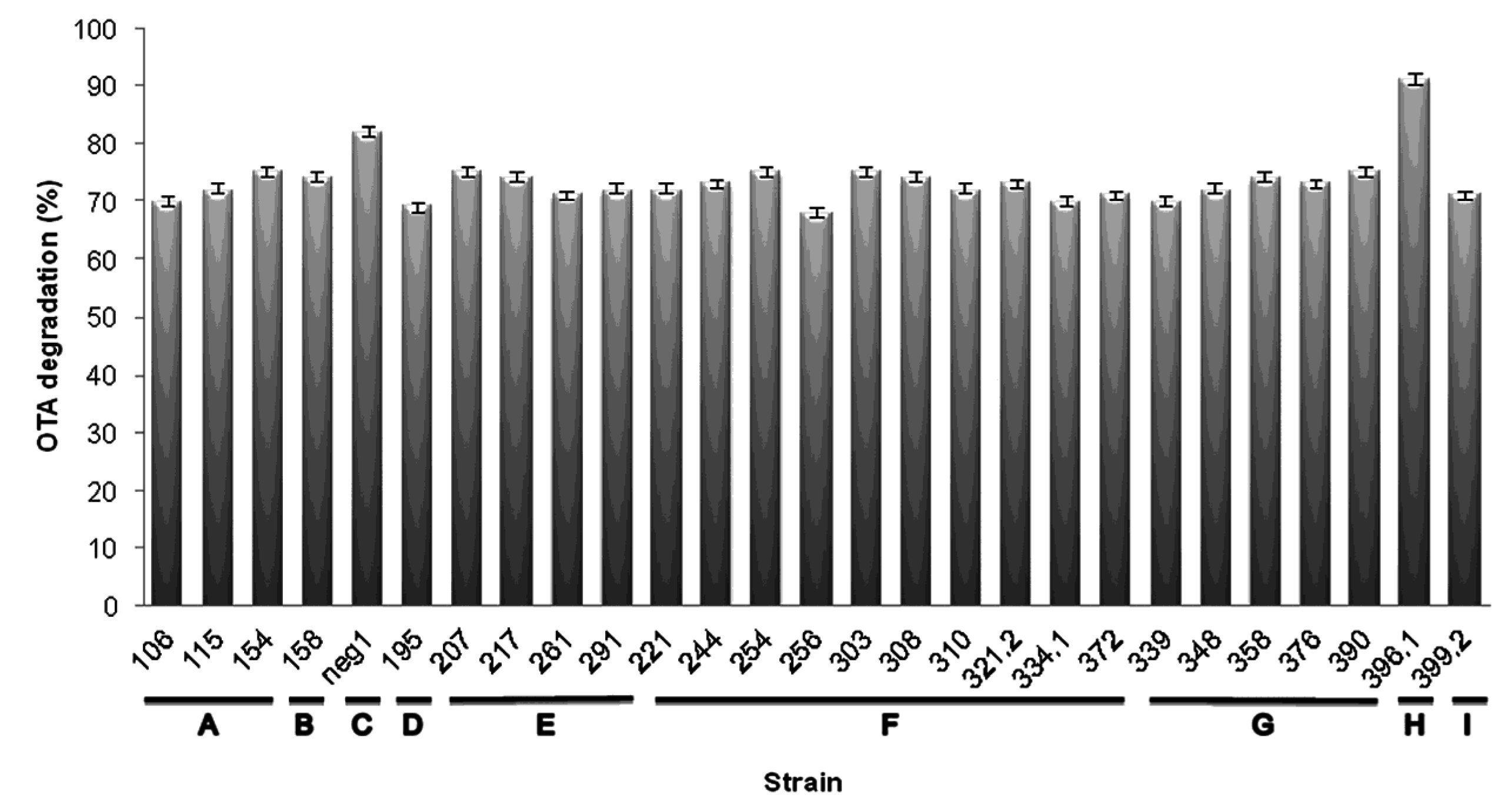
2.2. Genotypic Characterization and Identification of Bacteria from OTA-Degrading Soils

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain (REP-PCR Profile) | Isolation Soil | |
|---|---|---|---|
| Pseudomonas taiwanensis | 106 (1), 115 (2), 154 (3) | 4NG | |
| Leclercia adecarboxylata | 158 (4) | 4NG | |
| Acinetobacter sp. | neg1 (5) | 4NG | |
| Pantoea agglomerans | 195 (6) | 4NG | |
| Pseudomonas reinekei | 207 (7), 217 (8), 261 (13), 291 (14) | 5NG | |
| Pseudomonas koreensis | 221 (9), 244 (10), 254 (11), 256 (12) | 5NG | |
| 303 (15), 308 (16), 310 (17), 321.2 (18), 334.1 (19), 372 (23) | 3PR | ||
| Enterobacter aerogenes | 339 (20), 348 (21), 358 (22), 376 (24), 390 (26) | 3PR | |
| Acinetobacter calcoaceticus | 396.1 (25) | 3PR | |
| Enterobacter xiangfangensis | 399.2 (27) | 3PR | |
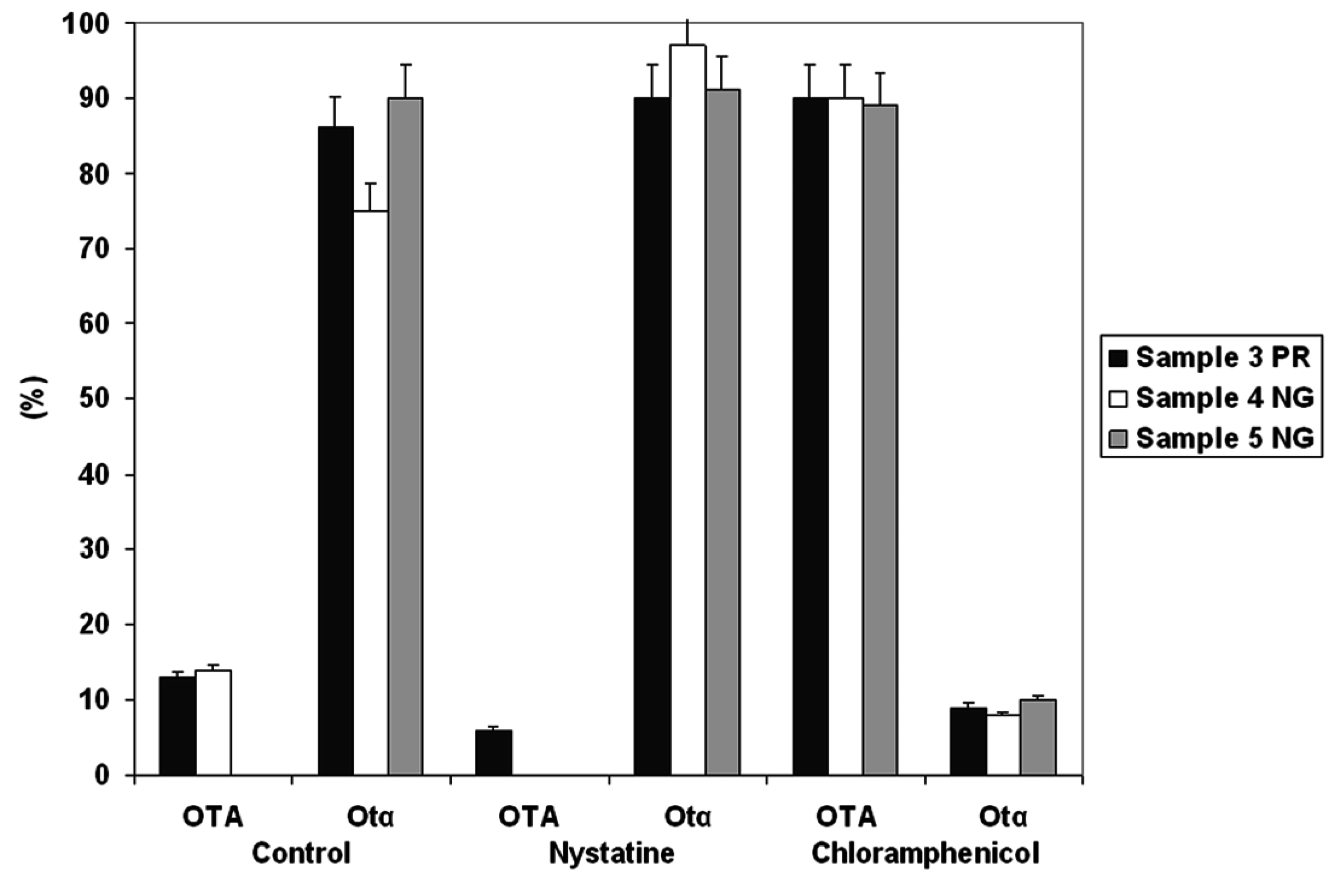
2.3. OTA Degradation by Acinetobacter spp.



3. Discussion
4. Experimental Section
4.1. Soil and Branch Sampling from Vineyards
| Sample ID | Origin | Vineyard n. | Cultivar | Site | Training System |
|---|---|---|---|---|---|
| 1 NG | soil | 1 | Negroamaro | Nardò (LE) | Bower system |
| 1 NGT | branches | 1 | Negroamaro | Nardò (LE) | Bower system |
| 2 PR | soil | 2 | Primitivo | Nardò (LE) | Bower system |
| 2 PRT | branches | 2 | Primitivo | Nardò (LE) | Bower system |
| 3 PR | soil | 3 | Primitivo | Manduria (TA) | Espalier |
| 3 PRT | branches | 3 | Primitivo | Manduria (TA) | Espalier |
| 4 NG | soil | 4 | Negroamaro | Cellino S. Marco (BR) | Head system |
| 4 NGT | branches | 4 | Negroamaro | Cellino S. Marco (BR) | Head system |
| 5 NG | soil | 5 | Negroamaro | Brindisi | Spur-pruned cordon |
| 5 NGT | branches | 5 | Negroamaro | Brindisi | Spur-pruned cordon |
4.2. Evaluation of the OTA-Degrading Ability of Soil and Branch Samples and Isolation of Microorganisms
4.3. rep-PCR
| Primer | Primer Sequence | Reference |
|---|---|---|
| REP-1R-Dt | 5′-IIINCGNCGNCATCNGGC-3′ | [63] |
| REP-2R-Dt | 5′- NCGNCTTATCNGGCCTAC-3′ | [63] |
| (GTG)5 | 5′-GTGGTGGTGGTGGTG-3′ | [64] |
| P0 | 5′-GAGAGTTTGATCCTGGCTCAG-3′ | [65] |
| P6 | 5′-CTACGGCTACCTTGTTACGA-3′ | [65] |
| 27f-YM | 5′-AGAGTTTGATYMTGGCTCAG-3′ | [66] |
| 1492r | 5′-TACCTTGTTACGACTT-3′ | [66] |
| ENT-16S-for | 5′-CAGCCACACTGGAACTGAGA-3′ | This study |
| ENT-16S-rev | 5′-GACAGCCATGCAGCACCT-3′ | This study |
| ENT-16S-revII | 5′-TTATGAGGTCCGCTTGCTCT-3′ | This study |
| PSE-16S-for | 5′-GGTCTTCGGATTGTAAAGCAC-3′ | This study |
| PSE-16S-rev | 5′-GACGACAGCCATGCAGC-3′ | This study |
4.4. Identification of Bacterial Strains
4.5. Biodegradation of OTA by Bacterial Isolates
4.6. Preparation of Standards
4.7. Chemical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Reddy, K.R.N.; Salleh, B.; Saad, B.; Abbas, H.K.; Abel, C.A.; Shier, W.T. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.D. Food safety and increasing hazard of mycotoxin occurrence in foods and feeds. Crit. Rev. Food Sci. Nutr. 2013, 53, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Benford, D.; Boyle, C.; Dekant, W.; Fuchs, R.; Gaylor, D.W.; Hard, G.; McGregor, D.B.; Pitt, J.I.; Plestina, R.; Shephard, G.; et al. Ochratoxin A. WHO Food Additives Series: 47. Safety evaluation of certain mycotoxins in food. In Proceedings of the 56th Joint FAO/WHO Expert Committee on Food Additives (JECFA), Geneva, Switzerland, 6–15 February 2001; Available online: http://www.inchem.org/documents /jecfa/jecmono/v47je04.htm (accessed on 10 June 2015).
- Wangikar, P.B.; Dwivedi, P.; Sinha, N.; Sharma, A.K.; Telang, A.G. Teratogenic effects in rabbits of simultaneous exposure to ochratoxin A and aflatoxin B1 with special reference to microscopic effects. Toxicology 2005, 215, 37–47. [Google Scholar] [CrossRef] [PubMed]
- IARC. Monograph on the Evaluation of Carcinogenic Risks to Human. In Some Traditional Herbal Medicines, Mycotoxins, Naphthalene and Styrene, Volume 82; World Health Organization, International Agency for Research on Cancer: Lyon, France, 2002. [Google Scholar]
- Kuiper-Goodman, T.; Scott, P.M. Risk assessment of the mycotoxin ochratoxin A. Biomed. Environ. Sci. 1989, 2, 179–248. [Google Scholar] [PubMed]
- Birzele, B.; Prange, A.; Krämer, J. Deoxynivalenol and ochratoxin A in German wheat and changes of level in relation to storage parameters. Food Addit. Contam. 2000, 17, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Fazekas, B.; Tar, A.K.; Zomborszky-Kovács, M. Ochratoxin A contamination of cereal grains and coffee in Hungary in the year 2001. Acta Vet. Hung. 2002, 50, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Roussos, S.; Zaouia, N.; Salih, G.; Tantaoui-Elaraki, A.; Lamrani, K.; Cheheb, M.; Hassouni, H.; Verhe, F.; Gaime, I.; Augur, C.; et al. Characterization of filamentous fungi isolated from Moroccan olive and olive cake: Toxinogenic potential of Aspergillus strains. Mol. Nutr. Food Res. 2006, 50, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Magan, N.; Logrieco, A. European research on ochratoxin A in grapes and wine. Int. J. Food Microbiol. 2006, 111, S2–S4. [Google Scholar] [CrossRef] [PubMed]
- Rosa, C.A.R.; Magnoli, C.E.; Fraga, M.E.; Dalcero, A.M.; Santana, D.M.N. Occurrence of ochratoxin A in wine and grape juice marketed in Rio de Janeiro, Brazil. Food Addit. Contam. 2004, 21, 358–364. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Bellí, N.; Mitchell, D.; Marín, S.; Alegre, I.; Ramos, A.J.; Magan, N.; Sanchis, V. Ochratoxin A-producing fungi in Spanish wine grapes and their relationship with meteorological conditions. Eur. J. Plant Pathol. 2005, 113, 233–239. [Google Scholar] [CrossRef]
- Battilani, P.; Giorni, P.; Bertuzzi, T.; Formenti, S.; Pietri, A. Black aspergilli and ochratoxin A in grapes in Italy. Int. J. Food Microbiol. 2006, 111, S53–S60. [Google Scholar] [CrossRef] [PubMed]
- Lucchetta, G.; Bazzo, I.; Cortivo, G.D.; Stringher, L.; Bellotto, D.; Borgo, M.; Angelini, E. Occurrence of black aspergilli and ochratoxin A on grapes in Italy. Toxins 2010, 2, 840–855. [Google Scholar] [CrossRef] [PubMed]
- Chiotta, M.L.; Ponsone, M.L.; Sosa, D.M.; Combina, M.; Chulze, S.N. Biodiversity of Aspergillus section Nigri populations in Argentinian vineyards and ochratoxin A contamination. Food Microbiol. 2013, 36, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Shi, J.; Zhu, C. Fruit spoilage and ochratoxin A production by Aspergillus carbonarius in the berries of different grape cultivars. Food Control. 2013, 30, 93–100. [Google Scholar] [CrossRef]
- Belkacem-Hanfi, N.; Fhoula, I.; Semmar, N.; Guesmi, A.; Perraud-Gaime, I.; Ouzari, H.I.; Boudabous, A.; Roussos, S. Lactic acid bacteria against post-harvest moulds and ochratoxin A isolated from stored wheat. Biol. Control 2014, 76, 52–59. [Google Scholar] [CrossRef]
- Visconti, A.; Perrone, G.; Cozzi, G.; Solfrizzo, M. Managing ochratoxin A risk in the grape-wine food chain. Food Addit. Contam. Part A 2008, 25, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Paterson, R.R.M.; Venâncio, A. Biodegradation of ochratoxin A for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitout, M.J. The hydrolysis of ochratoxin A by some proteolytic enzymes. Biochem. Pharmacol. 1969, 18, 485–491. [Google Scholar] [CrossRef]
- Wu, Q.; Dohnal, V.; Huang, L.; Kuča, K.; Wang, X.; Chen, G.; Yuan, Z. Metabolic pathways of ochratoxin A. Curr. Drug Metab. 2011, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ferenczi, S.; Cserháti, M.; Krifaton, C.; Szoboszlay, S.; Kukolya, J.; Szőke, Z.; Kőszegi, B.; Albert, M.; Barna, T.; Mezes, M.; et al. A New ochratoxin a biodegradation strategy using Cupriavidus basilensis Őr16 Strain. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Li, S.; Marquardt, R.R.; Frohlich, A.A.; Vitti, T.G.; Crow, G. Pharmacokinetics of ochratoxin A and its metabolites in rats. Toxicol. Appl. Pharmacol. 1997, 145, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Wegst, W.; Lingens, F. Bacterial degradation of ochratoxin A. FEMS Microbiol. Lett. 1983, 17, 341–344. [Google Scholar] [CrossRef]
- Hwang, C.A.; Draughon, F.A. Degradation of ochratoxin A by Acinetobacter calcoaceticus. J. Food Protec. 1994, 57, 410–414. [Google Scholar]
- Schatzmayr, G.; Zehner, F.; Täubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res. 2006, 50, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, H.; Reveron, I.; Doria, F.; Costantini, A.; de Las Rivas, B.; Muñoz, R.; Garcia-Moruno, E. Degradation of ochratoxin A by Brevibacterium species. J. Agric. Food Chem. 2011, 59, 10755–10760. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Inês, A.; Rodrigues, A.I.; Guimarães, A.; Pereira, V.L.; Parpot, P.; Mendes-Faia, A.; Venâncio, A. Biodegradation of ochratoxin A by Pediococcus parvulus isolated from Douro wines. Int. J. Food Microbiol. 2014, 188, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, J.; Rigó, K.; Téren, J. Degradation of ochratoxin A by Aspergillus species. Int. J. Food Microbiol. 2000, 59, 1–7. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Venâncio, A. Isolation and purification of an enzyme hydrolyzing ochratoxin A from Aspergillus niger. Biotechnol. Lett. 2007, 29, 1909–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amézqueta, S.; González-Peñas, E.; Murillo-Arbizu, M.; López de Cerain, A. Ochratoxin A decontamination: A review. Food Control 2009, 20, 326–333. [Google Scholar] [CrossRef]
- Islam, R.; Zhou, T.; Young, J.C.; Goodwin, P.H.; Pauls, K.P. Aerobic and anaerobic de-epoxydation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil. World J. Microbiol. Biotechnol. 2012, 28, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Ito, M.; Ishizaka, M.; Ikunaga, Y.; Sato, Y.; Yoshida, S.; Koitabashi, M.; Tsushima, S. Thirteen novel deoxynivalenol-degrading bacteria are classified within two genera with distinct degradation mechanisms. FEMS Microbiol. Lett. 2012, 327, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Nagarajan, K.; Loh, K.C. Biodegradation of aromatic compounds: Current status and opportunities for biomolecular approaches. Appl. Microbiol. Biotechnol. 2009, 85, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl. Environ. Microbiol. 1997, 63, 3825–3830. [Google Scholar] [PubMed]
- Fanelli, F.; Chiara, M.; Liuzzi, V.C.; Haidukowski, M.; Tristezza, M.; Caterina, M.; D’Erchia, A.M.; Pesole, G.; Horner, D.S.; Mulè, G. Draft genome sequence of Acinetobacter sp. neg1 capable of degrading ochratoxin A. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef]
- Karigar, C.S.; Rao, S.S. Role of microbial enzymes in the bioremediation of pollutants: A review. Enzyme Res. 2011, 805187. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, G.K.; Strobel, B.W.; Hansen, H.C.B. Determination of zearalenone and ochratoxin A in soil. Anal. Bioanal. Chem. 2003, 376, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Somma, S.; Perrone, G.; Logrieco, A.F. Diversity of black Aspergilli and mycotoxin risks in grape, wine and dried vine fruits. Phytopathol. Mediterr. 2012, 51, 131–147. [Google Scholar] [CrossRef]
- Mortensen, G.K.; Strobel, B.W.; Hansen, H.C.B. Degradation of zearalenone and ochratoxin A in three Danish agricultural soils. Chemosphere 2006, 62, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Chekol, T.; Vough, L.R.; Chaney, R.L. Phytoremediation of polychlorinated biphenyl-contaminated soils: The rhizosphere effect. Environ. Int. 2004, 30, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.E.R.; Joseph, S.J.; Janssen, P.H. Effects of growth medium, inoculum size, and incubation time on culturability and isolation of soil bacteria. Appl. Environ. Microbiol. 2005, 71, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; He, J.; Young, J.C.; Zhu, H.; Li, X.Z.; Ji, C.; Zhou, T. Transformation of trichothecene mycotoxins by microorganisms from fish digesta. Aquaculture 2009, 290, 290–295. [Google Scholar] [CrossRef]
- Furukawa, K.; Chakrabarty, A.M. Involvement of plasmids in total degradation of chlorinated biphenyls. Appl. Environ. Microbiol. 1982, 44, 619–626. [Google Scholar] [PubMed]
- Chaudhry, G.R.; Ali, A.N.; Wheeler, W.B. Isolation of a methyl parathion-degrading Pseudomonas sp. that possesses DNA homologous to the opd gene from a Flavobacterium sp. Appl. Environ. Microbiol. 1988, 54, 288–293. [Google Scholar] [PubMed]
- Jung, J.; Park, W. Acinetobacter species as model microorganisms in environmental microbiology: Current state and perspectives. Appl. Microbiol. Biotechnol. 2015, 99, 2533–2548. [Google Scholar] [CrossRef] [PubMed]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): Screening for aflatoxin B1 and ochratoxin A detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Skrinjar, M.; Rasić, J.L.; Stojicić, V. Lowering of ochratoxin A level in milk by yoghurt bacteria and bifidobacteria. Folia Microbiol. (Praha) 1996, 41, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Böhm, J.; Grajewski, J.; Asperger, H.; Rabus, B.; Razzazi, E. Study on biodegradation of some A- and B-trichothecenes and ochratoxin A by use of probiotic microorganisms. Mycotoxin. Res. 2000, 16, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Wu, Z.; Wu, S.; Dai, Y.; Sun, C. Degradation of ochratoxin A by Bacillus amyloliquefaciens ASAG1. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2014, 32, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Marquard, R.R.; Frohlich, A.A.; Ling, Y.Z. Synthesis and structure elucidation of analogues of ochratoxin A. J. Agric. Food Chem. 1995, 43, 524–530. [Google Scholar] [CrossRef]
- Bittner, A.; Cramer, B.; Harrer, H.; Humpf, H.U. Structure elucidation and in vitro cytotoxicity of ochratoxin α amide, a new degradation product of ochratoxin A. Mycotoxin Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Doster, R.C.; Sinnhuber, R.O. Comparative rates of hydrolysis of ochratoxins A and B in vitro. Food Cosmet. Toxicol. 1972, 10, 389–394. [Google Scholar] [CrossRef]
- Stander, M.A.; Steyn, P.S.; van der Westhuizen, F.H.; Payne, B.E. A kinetic study into the hydrolysis of the ochratoxins and analogues by carboxypeptidase A. Chem. Res. Toxicol. 2001, 14, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Santos, L.; Venâncio, A. Degradation of ochratoxin A by proteases and by a crude enzyme of Aspergillus niger. Food Biotechnol. 2006, 20, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Battilani, P.; Logrieco, A.; Giorni, P.; Cozzi, G.; Bertuzzi, T.; Pietri, A. Ochratoxin A production by Aspergillus carbonarius on some grape varieties grown in Italy. J. Sci. Food. Agric. 2004, 84, 1736–1740. [Google Scholar] [CrossRef]
- Bleve, G.; Lezzi, C.; Chiriatti, M.A.; D’Ostuni, I.; Tristezza, M.; Di Venere, D.; Sergio, L.; Mita, G.; Grieco, F. Selection of non-conventional yeasts and their use in immobilized form for the bioremediation of olive oil mill wastewaters. Bioresour. Technol. 2011, 102, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucl. Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef] [PubMed]
- Tobes, R.; Ramos, J.L. REP code: Defining bacterial identity in extragenic space. Environ. Microbiol. 2005, 7, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Hyytiä-Trees, E.; Lyhs, U.; Korkeala, H.; Björkroth, J. Characterisation of ropy slime-producing Lactobacillus sakei using repetitive element sequence-based PCR. Int. J. Food Microbiol. 1999, 50, 215–219. [Google Scholar] [CrossRef]
- Coudeyras, S.; Marchandin, H.; Fajon, C.; Forestier, C. Taxonomic and strain-specific identification of the probiotic strain Lactobacillus rhamnosus 35 within the Lactobacillus casei group. Appl. Environ. Microbiol. 2008, 74, 2679–2689. [Google Scholar] [CrossRef] [PubMed]
- Di Cello, F.; Bevivino, A.; Chiarini, L.; Fani, R.; Paffetti, D.; Tabacchioni, S.; Dalmastri, C. Biodiversity of a Burkholderia cepacia population isolated from maize rhizosphere at different plant growth stages. Appl. Environ. Microbiol. 1997, 63, 4485–4493. [Google Scholar] [PubMed]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Ladero, V.; Beneduce, L.; Fernández, M.; Alvarez, M.A.; Benoit, B.; Laurent, B.; Grieco, F.; Spano, G. Isolation and characterization of tyramine-producing Enterococcus faecium strains from red wine. Food Microbiol. 2011, 28, 434–439. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov (accessed on 10 July 2015).
- Gallo, A.; Bruno, K.S.; Solfrizzo, M.; Perrone, G.; Mulè, G.; Visconti, A.; Baker, S.E. New insight into the ochratoxin A biosynthetic pathway through deletion of a nonribosomal peptide synthetase gene in Aspergillus carbonarius. Appl. Environ. Microbiol. 2012, 78, 8208–8218. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Bellis, P.; Tristezza, M.; Haidukowski, M.; Fanelli, F.; Sisto, A.; Mulè, G.; Grieco, F. Biodegradation of Ochratoxin A by Bacterial Strains Isolated from Vineyard Soils. Toxins 2015, 7, 5079-5093. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7124864
De Bellis P, Tristezza M, Haidukowski M, Fanelli F, Sisto A, Mulè G, Grieco F. Biodegradation of Ochratoxin A by Bacterial Strains Isolated from Vineyard Soils. Toxins. 2015; 7(12):5079-5093. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7124864
Chicago/Turabian StyleDe Bellis, Palmira, Mariana Tristezza, Miriam Haidukowski, Francesca Fanelli, Angelo Sisto, Giuseppina Mulè, and Francesco Grieco. 2015. "Biodegradation of Ochratoxin A by Bacterial Strains Isolated from Vineyard Soils" Toxins 7, no. 12: 5079-5093. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7124864