
Research on Acute Toxicity and the Behavioral Effects of Methanolic Extract from Psilocybin Mushrooms and Psilocin in Mice

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Acute Toxicity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LD50 values | ||
|---|---|---|
| methanol extract I Pholiotina cyanopus | methanol extract II Psilocybe semilanceata | Psilocin |
| 316.87 ± 1.63 | 324.37 ± 2.04 | 293.07 ± 1.02 |
| Determined compounds | Chemical formula | Percentage content in dry mass of mushroom | Percentage content in dry extract | ||
|---|---|---|---|---|---|
| Ph. cyanopus | P. semilanceata | Ph. cyanopus | P. semilanceata | ||
| Norbaeoystin |  C10H13N2O4P | 0.053 ± 0.004 | 0.077 ± 0.006 | 0.424 ± 0.032 | 0.481 ± 0.038 |
| Baeocystin |  C11H15N2O4P | 0.16 ± 0.01 | ND | 1.28 ± 0.08 | ND |
| Psilocybin |  C12H17N2O4P | 0.9 ± 0.08 | 1.46 ± 0.12 | 7.2 ± 0.64 | 9.125 ± 0.75 |
| Psilocin |  C12H16N2O | 0.17 ± 0.01 | 0.24 ± 0.02 | 1.36 ± 0.08 | 1.5 ± 0.125 |
| Aeruginascin |  C13H20N2O4P | 0.011 ± 0.0007 | 0.022 ± 0.01 | 0.88 ± 0.006 | 0.138 ± 0.062 |
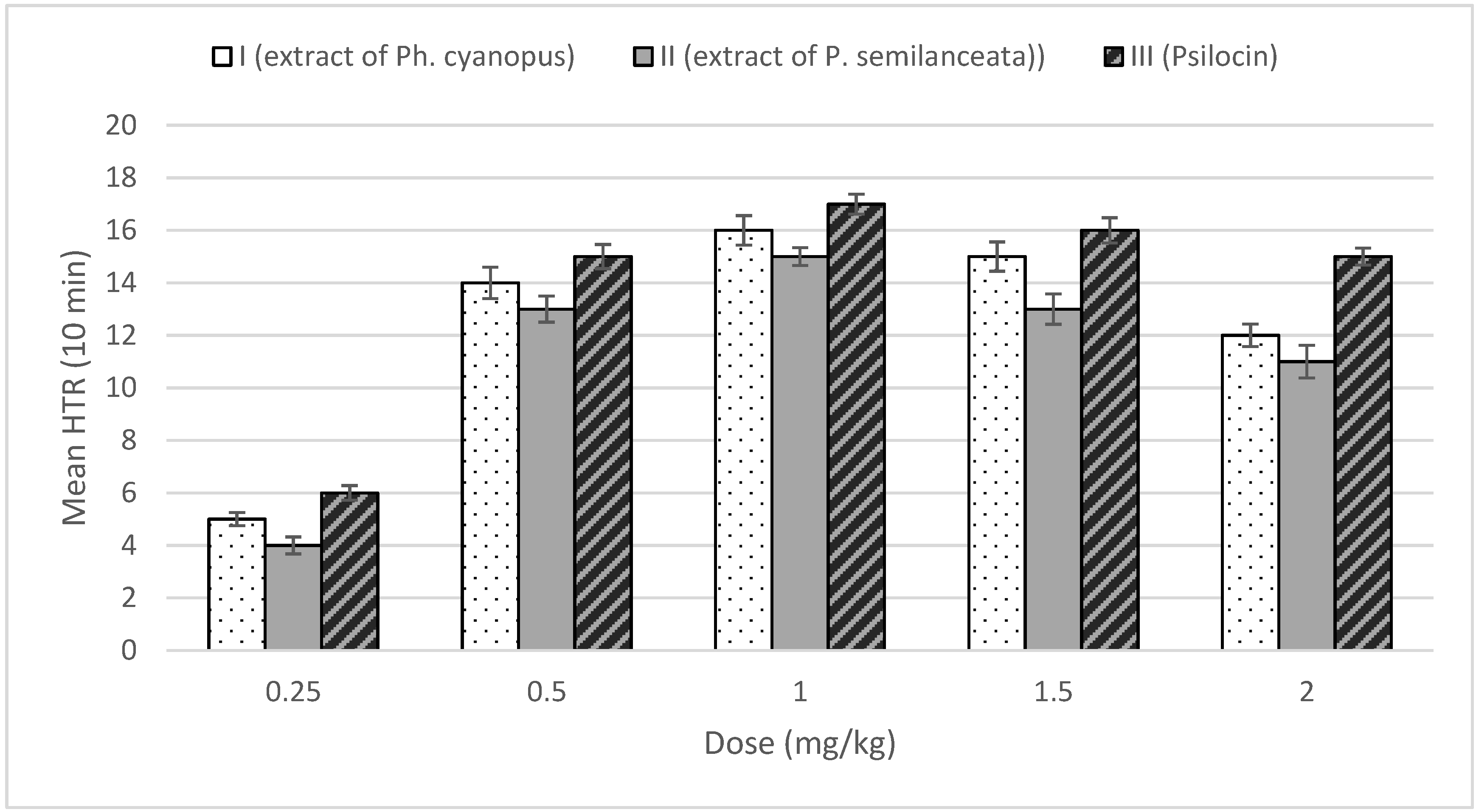
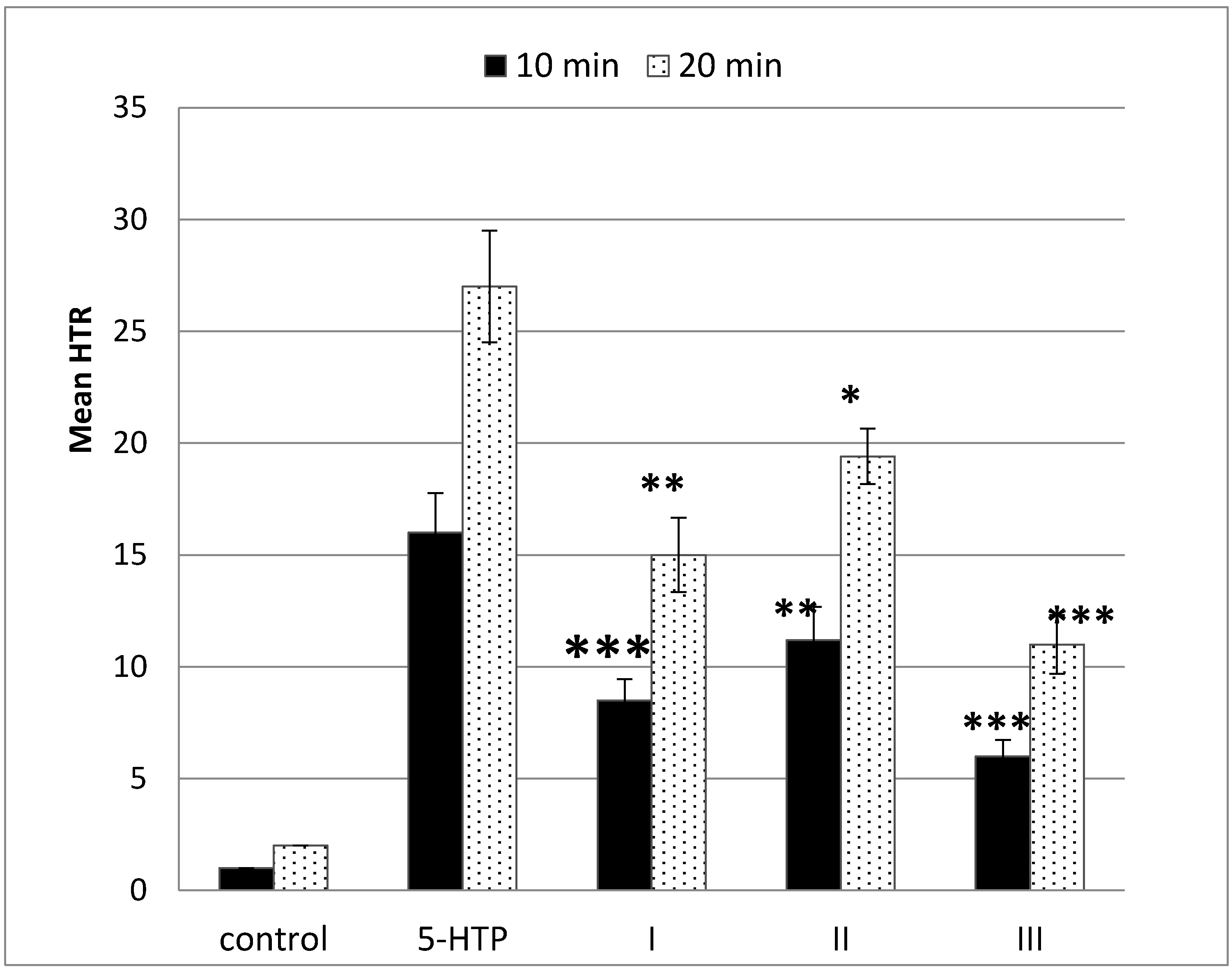
2.2. Behavioral Effect—Head-Twitch Response (HTR)


3. Experimental Section
3.1. Mushroom Material
3.2. LC-MS Analysis
3.3. Experimental Animals
3.4. Experimental Procedure
3.5. Acute Toxicity Assessment
3.6. Behavioral Effect Assessment
3.7. Statistical Methods
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Repke, D.B.; Leslie, D.T.; Guzmán, G. Baeocystin in psilocybe, conocybe and panaeolus. Lloydia 1977, 40, 566–578. [Google Scholar] [PubMed]
- Merlin, M.D.; Allen, J.W. Species identification and chemical analysis of psychoactive fungi in the Hawaiian islands. J. Ethnopharmacol. 1993, 40, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, L.S. Indole derivatives in certain Panaeolus species from East Europe and Siberia. Mycol. Res. 1993, 97, 251–254. [Google Scholar] [CrossRef]
- Gartz, J. Cultivation and analysis of Psilocybe species and an investigation of Galerina steglichii. Ann. Mus. Rov. 1994, 10, 297–306. [Google Scholar]
- Gartz, J. Extraction and analysis of indole derivatives from fungal biomass. J. Basic Microbiol. 1994, 34, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Gartz, J. Analysis of aeruginascin in fruit bodies of the mushroom Inocybe aeruginascens. Int. J. Crude Drug Res. 1989, 27, 141–144. [Google Scholar]
- Gartz, J. Inocybe aeruginascens Babos. Eleusis 1995, 3, 31–34. [Google Scholar]
- Leung, A.Y.; Paul, A.G. Baeocystin, a mono-methyl analog of psilocybin from Psilocybe baeocystis saphrophytic culture. J. Pharm. Sci. 1967, 56, 146. [Google Scholar] [CrossRef] [PubMed]
- Stijve, T.; Kuyper, T.W. Occurrence of psilocybin in various higher fungi from several European countries. Planta. Med. 1985, 51, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Borner, S.; Brenneisen, R. Determination of tryptamine derivatives in hallucinogenic mushrooms using high-performance liquid chromatography with photodiode array detection. J. Chromatogr. A 1987, 408, 402–408. [Google Scholar] [CrossRef]
- Vanhaelen-Fastré, R.; Vanhaelen, M. Qualitative and quantitative determinations of hallucinogenic components of Psilocybe mushrooms by reversed-phase high-performance liquid chromatography. J. Chromatogr. A. 1984, 312, 467–472. [Google Scholar] [CrossRef]
- Christiansen, A.L.; Rasmussen, K.E. Analysis of indole alkaloids in Norwegian Psilocybe semilanceata using high-performance liquid chromatography and mass spectrometry. J. Chromatogr. A 1982, 244, 357–364. [Google Scholar] [CrossRef]
- Stebelska, K. Fungal Hallucinogens psilocin, ibotenic acid, and muscimol: Analytical methods and biologic activities. Ther. Drug Monit. 2013, 35, 420–442. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.E. Hallucinogens. Pharmacol. Ther. 2004, 101, 131–181. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Maeso, J.; Weisstaub, N.V.; Zhou, M.; Chan, P.; Ivic, L.; Ang, R.; Lira, A.; Bradley-Moore, M.; Ge, Y.; Zhou, Q.; et al. Hallucinogens recruit specific cortical 5-HT(2A) receptor-mediated signaling pathways to affect behavior. Neuron 2007, 53, 439–452. [Google Scholar] [CrossRef]
- Dooley, D.J.; Quock, R.M. Tryptamine and 5-hydroxytryptamine-induced hypothermia in mice. J. Pharm Pharmacol. 1976, 10, 775–776. [Google Scholar] [CrossRef]
- Canal, C.E.; Olaghere da Silva, U.B.; Gresch, P.J.; Watt, E.E.; Sanders-Bush, E.; Airey, D.C. The serotonin 2C receptor potently modulates the head-twitch response in mice induced by a phenethylamine hallucinogen. Psychopharmacology (Berl) 2010, 209, 163–174. [Google Scholar] [CrossRef]
- Pranzatelli, M.R. Regulation of 5-HT2 receptors in rat cortex. Studies with a putative selective agonist and an antagonist. Biochem. Pharmacol. 1991, 42, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.J.; Peroutka, S.J. Differentiation of 5-hydroxytryptamine 2 receptor subtypes using 125I-R-(-)2,5-dimethoxy-4-iodo-phenyl-isopropylamine and 3H-ketanserin. J. Neurosci. 1989, 10, 3482–3490. [Google Scholar]
- Matsumoto, K.; Mizowaki, M.; Takayama, H.; Sakai, S.I.; Aimi, N.; Watanabe, H. Suppressive effect of mitragynine on the 5-Methoxy-N,N-dimethyltryptamine-induced Head-Twitch response in mice. Pharmacol. Biochem. Behav. 19 1997, 57, 319–323. [Google Scholar] [CrossRef]
- Apud, J.A. The 5-HT2 receptor in brain: Recent biochemical and molecular biological developments and new perspectives in its regulation. Pharmacol. Res. 1991, 23, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.B.; Kurrasch-Orbaugh, D.; Marona-Lewicka, D.; Cumbay, M.G.; Watts, V.J.; Barker, E.L.; Nichols, D.E. Effect of ring fluorination on the pharmacology of hallucinogenic tryptamines. J. Med. Chem. 2000, 43, 4701–4710. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.J.; Repke, D.B.; Lo, L.; Peroutka, S.J. Differential interactions of indolealkylamines with 5-hydroxytryptamine receptor subtypes. Neuropharmacology 1990, 29, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Lindenblatt, H.; Kramer, E.; Holzmann-Erens, P.; Gouzoulis-Mayfrank, E.; Kovar, K.-A. Quantitation of psilocin in human plasma by high-performance liquid chromatography and electrochemical detection: Comparison of liquid extraction with automated on-line solid-phase extraction. J. Chromatogr. B 1998, 709, 255–263. [Google Scholar] [CrossRef]
- Canal, C.E.; Morgan, D. Head-twitch response in rodents induced by the hallucinogen 2,5-dimethoxy-4-iodoamphetamine: a comprehensive history, a re-evaluation of mechanisms, and its utility as a model. Drug Test Anal. 2012, 4, 556–576. [Google Scholar] [CrossRef] [PubMed]
- Halberstadt, A.L.; Geyer, M.A. Multiple receptors contribute to the behavioral effects of indoleamine hallucinogens. Neuropharmacology 2011, 61, 364–381. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, E.; Kafarski, P. Mannitol in Amanita muscaria—An osmotic blood-brain barrier disruptor enhancing its hallucinogenic action. Med. Hypothesis 2013, 81, 766–767. [Google Scholar] [CrossRef]
- Colpaert, F.C.; Janssen, P.A. The head-twitch response to intraperitoneal injection of 5-hydroxytryptophan in the rat: antagonist effects of purported 5-hydroxytryptamine antagonists and of pirenperone, an LSD antagonist. Psychopharmacology 2010, 209, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Halberstadt, A.L.; Koedood, L.; Powell, S.B.; Geyer, M.A. Differential contributions of serotonin receptors to the behavioral effects of indoleamine hallucinogens in mice. J. Psychopharmacol. 2011, 25, 1548–1561. [Google Scholar] [CrossRef] [PubMed]
- Corne, S.J.; Pickering, R.W.; Warner, B.T. Method for assessing the effects of drugs on the central actions of 5-hydroxytryptamine. Br. J. Pharmacol. 1963, 20, 106–120. [Google Scholar]
- Soloviov, V.H.; Firsov, A.A.; Filov, B.A. Farmakokinetyka; Medycyna: Moskwa, Rosja, 1980. [Google Scholar]
- Young, S.N. Single treatments that have lasting effects: Some thoughts on the antidepressant effects of ketamine and botulinum toxin and the anxiolytic effect of psilocybin. J. Psychiatry Neurosci. 2013, 38, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Grob, C.S.; Danforth, A.I.; Chopra, G.S.; Hagerty, M.; McKay, C.; Halberstadt, A.; Greer, G.R. Pilot study of psilocybin treatment for anxiety in patients with advanced-stage cancer. Arch. Gen. Psychiatry 2011, 68, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Studerus, E.; Kometer, M.; Hasler, F.; Vollenweider, F. Acute, subacute and longterm subjective effects of psilocybin in healthy humans: A pooled analysis of experimental studies. J. Psychopharmacol. 2011, 25, 1434–1452. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, D.; Barnes, G.; Giaroli, G.; Tracy, D. Classical hallucinogens as antidepressants? A review of pharmacodynamics and putative clinical roles. Ther. Adv. Psychopharmacol. 2014, 4, 156–169. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuk, O.; Jasicka-Misiak, I.; Poliwoda, A.; Kazakova, A.; Godovan, V.V.; Halama, M.; Wieczorek, P.P. Research on Acute Toxicity and the Behavioral Effects of Methanolic Extract from Psilocybin Mushrooms and Psilocin in Mice. Toxins 2015, 7, 1018-1029. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7041018
Zhuk O, Jasicka-Misiak I, Poliwoda A, Kazakova A, Godovan VV, Halama M, Wieczorek PP. Research on Acute Toxicity and the Behavioral Effects of Methanolic Extract from Psilocybin Mushrooms and Psilocin in Mice. Toxins. 2015; 7(4):1018-1029. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7041018
Chicago/Turabian StyleZhuk, Olga, Izabela Jasicka-Misiak, Anna Poliwoda, Anastasia Kazakova, Vladlena V. Godovan, Marek Halama, and Piotr P. Wieczorek. 2015. "Research on Acute Toxicity and the Behavioral Effects of Methanolic Extract from Psilocybin Mushrooms and Psilocin in Mice" Toxins 7, no. 4: 1018-1029. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7041018