
Separation of Binding Protein of Celangulin V from the Midgut of Mythimna separata Walker by Affinity Chromatography

Abstract
:1. Introduction

2. Results and Discussion
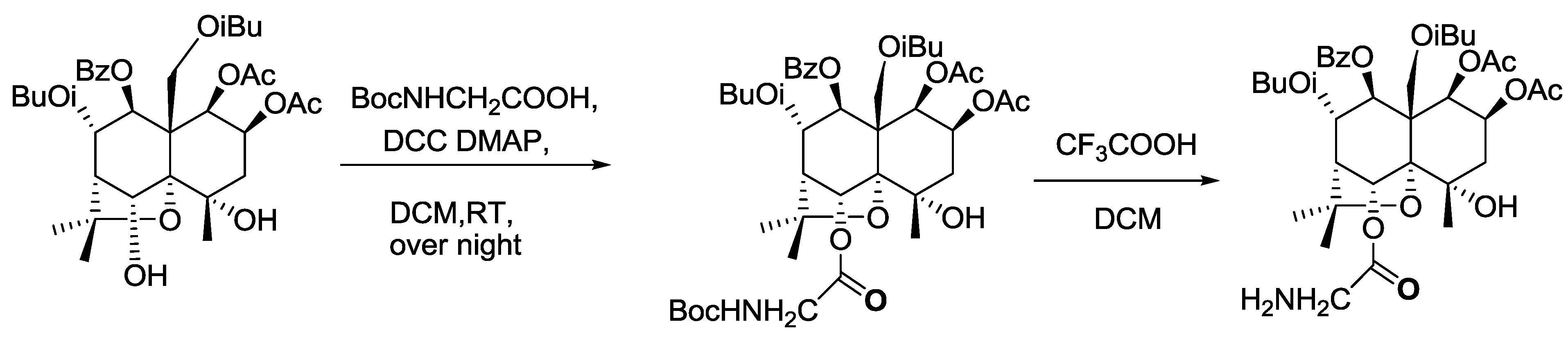
2.1. Identification of the Ligand (Celangulin V-6-aminoacetic Acid Ester)

2.2. Separation of Binding Protein


{kind=link}
{kind=link}
{kind=link}
| Number | Protein | Genbank gi number | Species |
|---|---|---|---|
| 1 | Actin | gi|108879764 | Aedes aegypti |
| 2 | Zinc finger protein | gi|157127505 | Aedes aegypti |
| 3 | Thioredoxin peroxidase | gi|7230426 | Drosophila melanogaster |
| 4 | Glyceraldehyde-3-phosphate dehydrogenase | gi|112983816 | Bombyx mori |
| 5 | Transmembrane protein 1 | gi|108872951 | Aedes aegypti |
| 6 | E3 SUMO-protein ligase RanBP2 | gi|307201149 | Harpegnathos saltator |
| 7 | Amino peptidase N3 | gi|21327773 | Plutella xylostella |
| 8 | Protease m1 zinc metalloprotease | gi|108875833 | Aedes aegypti |
| 9 | Vacuolar ATPase subunit a | gi|5852166 | Manduca sexta |
| 10 | Vacuolar ATPase subunit B | gi|8810 | Drosophila melanogaster |
| 11 | Vacuolar ATPase subunit H | gi|5852164 | Manduca sexta |
3. Experimental Section
3.1. Insects
3.2. Chemicals
3.3. Isolation of BBMV from M. Separata Walker
3.4. Synthesis of Ligand
3.4.1. Synthesis of Celangulin V-6-Boc Aminoacetic Acid Ester
3.4.2. Synthesis of Celangulin V-6-aminoacetic Acid Ester
3.5. Preparing the Medium and Coupling the Ligand
3.6. Binding, Elution, and Regeneration
3.7. SDS-PAGE and LC/Q-TOF-MS
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Rosell, G.; Quero, C.; Coll, J.; Guerrero, A. Biorational insecticides in pest management. J. Pestic. Sci. 2008, 33, 103–121. [Google Scholar] [CrossRef]
- Grzybowski, A.; Tiboni, M.; Silva, M.A.; Chitolina, R.F.; Passos, M.; Fontana, J.D. Synergistic larvicidal effect and morphological alterations induced by ethanolic extracts of Annona muricata and Piper nigrum against the dengue fever vector Aedes aegypti. Pest Manag. Sci. 2013, 69, 589–601. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, L. A brief review on insecticidal function of azadirachta indica. Pestic. Sci. Adm. 2012, 33, 23–28. [Google Scholar]
- Wang, L.; He, L.; Shen, Y. Advances in studies on insecticidal function of meliaceae. Pestic. Sci. Adm. 2013, 34, 6–9. [Google Scholar]
- Deepa, M.A.; Bai, V.N. Bioinsecticidal compounds of celastraceae-the spindle tree family. Int. J. Bot. 2010, 6, 220. [Google Scholar] [CrossRef]
- Wei, S.; Wang, M.; Ji, Z.; Shi, B.; Li, S.; Zhang, J. Three new insecticidal sesquiterpene polyol esters from Celastrus angulatus. Nat. Prod. Commun. 2010, 5, 355–359. [Google Scholar] [PubMed]
- Karamaouna, F.; Kimbaris, A.; Michaelakis, A.; Papachristos, D.; Polissiou, M.; Papatsakona, P.; Tsora, E. Insecticidal activity of plant essential oils against the vine mealybug, Planococcus ficus. J. Insect Sci. 2013, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Raturi, R.; Badoni, P.P.; Ballabha, R. Insecticidal and fungicidal activities of stem bark of Zanthoxylum Armatum (Rutaceae). Word J. Pharm. Pharm. Sci. 2014, 3, 1838–1843. [Google Scholar]
- Isman, M.B.; Grieneisen, M.L. Botanical insecticide research: many publications, limited useful data. Trends Plant Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.M.; Wu, W.J.; Zhang, J.W.; Konishi, Y. The dihydro-β-agarofuran sesquiterpenoids. Nat. Prod. Rep. 2007, 5, 1153–1189. [Google Scholar] [CrossRef]
- Ji, Z.; Wu, W.; Yang, H.; Shi, B.; Wang, M. Four novel insecticidal sesquiterpene esters from Celastrus angulatus. Nat. Prod. Res. 2007, 21, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Zhang, Q.; Shi, B.; Wei, S.; Wang, M.; Wu, W. Three new insecticidal sesquiterpene pyridine alkaloids from Celastrus angulatus. Nat. Prod. Res. Former. Nat. Prod. Lett. 2009, 23, 470–478. [Google Scholar]
- Wu, W.J.; Li, S.B.; Zhu, J.B.; Liu, H.X. New sequiterpenoid CelangulinV: Isolation and determination. Acta Univ. Agric. Boreali-Occidentalis 1994, 22, 116–117. [Google Scholar]
- Yang, R.Y.; Liu, H.X.; Wu, W.J.; Wang, J.L. Study on the functioning mechanism of celangulin V. Northwest Sci.-Technol. Univ. Agri. For. (Nat. Sci. Ed.) 2001, 29, 77–79. [Google Scholar]
- Qi, Z.; Shi, B.; Hu, Z.; Zhang, Y.; Wu, W. Ultrastructural effects of Celangulin V on midgut cells of the oriental armyworm, Mythimna separata walker (Lepidoptera: Noctuidae). Ecotoxicol. Environ. Saf. 2011, 74, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Ji, Z.Q.; Hu, Z.N. Natural products and digestive poisons. Pesticides 1997, 36, 6–9. [Google Scholar]
- Wu, W.J.; Hu, Z.N.; Liu, H.X.; Qi, Z.J. Insecticidal mechanisms of the major active components from the Chinese bittersweet, Celastrus angulatus and their application. Acta Entomol. Sin. 2005, 48, 770–777. [Google Scholar]
- Zhang, J.; Hu, Z.; Li, S.; Wu, W. Synthesis and insecticidal activities of new ester-derivatives of Celangulin-V. Int. J. Mol. Sci. 2011, 12, 9596–9604. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Zhao, F.; Li, Y. Advances in research on zinc finger protein. Lett. Biotechnol. 2009, 20, 131–134. [Google Scholar]
- Zhang, P.; Liu, B.; Kang, S.W.; Seo, M.S.; Rhee, S.G.; Obeid, L.M. Thioredoxin peroxidase is a novel inhibitor of apoptosis with a mechanism distinct from that of Bcl-2. J. Biol. Chem. 1997, 272, 30615–30618. [Google Scholar] [CrossRef] [PubMed]
- Tarze, A.; Deniaud, A.; Bras, M.L.; Maillier, E.; Molle, D.; Larochette, N.; Zamzami, N.; Jan, G.; Kroemer, G.; Brenner, C. GAPDH, a novel regulator of the pro-apoptotic mitochondrial membrane permeabilization. Oncogene 2006, 26, 2606–2620. [Google Scholar] [CrossRef] [PubMed]
- Zala, D.; Hinckelmann, M.-V.; Yu, H.; Cunha, M.M.L.D.; Liot, G.; Cordelières, F.P.; Marco, S.; Saudou, F. Vesicular glycolysis provides on-board energy for fast axonal transport. Cell 2013, 152, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Hay, R.T. SUMO: A history of modification. Mol. Cell 2005, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pichler, A.; Gast, A.; Seeler, J.S.; Dejean, A.; Melchior, F. The nucleoporin RanBP2 has SUMO1 E3 ligase activity. Cell 2002, 108, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Kirsh, O.; Seeler, J.S.; Pichler, A.; Gast, A.; Muller, S.; Miska, E.; Mathieu, M.; Harel-Bellan, A.; Kouzarides, T.; Melchior, F.; et al. The SUMO E3 ligase RanBP2 promotes modification of the HDAC4 deacetylase. EMBO J. 2002, 21, 2682–2691. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Ding, X.; Xia, L.; Yin, J.; Huang, S.; Huang, F. Proteomic analysisi of BBMV in Helicoverpa armigera midgut with and without Cry1Ac toxin treatment. Biocontrol Sci. Technol. 2011, 21, 139–151. [Google Scholar] [CrossRef]
- Doherty, G.J.; McMahon, H.T. Mediation, modulation, and consequences of membrane-cytoskeleton interactions. Annu. Rev. Biophys. 2008, 37, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Wu, Q.; Wang, S.; Xu, B.; Zhang, Y. Studying progress on aminopeptidase N of insects. Chin. J. Pestic. Sci. 2011, 13, 213–220. [Google Scholar]
- Knight, P.; Carroll, J.; Ellar, D. Analysis of glycan structures on the 120 kDa aminopeptidase N of Manduca sexta and their interactions with Bacillus thuringiensis Cry1Ac toxin. Insect Biochem. Mol. Biol. 2004, 34, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Tetreau, G.; Bayyareddy, K.; Jones, C.M.; Stalinski, R.; Riaz, M.A.; Paris, M.; David, J.-P.; Adang, M.J.; Després, L. Larval midgut modifications associated with Bti resistance in the yellow fever mosquito using proteomic and transcriptomic approaches. BMC Genomics 2012, 13, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnepf, E.; Crickmore, N.; Rie, J.V.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [PubMed]
- Nelson, N. A journey from mammals to yeast with vacuolar H+-ATPase (V-ATPase). J. Bioenerg. Biomembr. 2003, 35, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, H.; Huss, M.; Merzendorfer, H.; Reineke, S.; Vitavska, O.; Zeiske, W. The insect plasma membrane H+ V-ATPase: Intra-, Inter-, and supramolecular aspects. J. Bioenerg. Biomembr. 2003, 35, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, H.; Beyenbach, K.W.; Huss, M.; Vitavska, O. Vacuolar-type proton pumps in insect epithelia. J. Exp. Biol. 2009, 212, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Bowman, E.J.; Bowman, B.J. V-ATPases as drug targets. J. Bioenerg. Biomembr. 2005, 37, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Wolfersberger, M.G.; Luthy, P.; Maurer, A.; Parenti, P.; Sacchi, V.F.; Giordana, B.; Hanozet, G.M. Preparation and partial characterization of amino acid transporting brush border membrane vesicles from the larval midgut of the cabbage butterfly (Pieris brassicae). Comp. Biochem. Physiol. Part A Physiol. 1987, 86, 301–308. [Google Scholar] [CrossRef]
- Ferré, J.; Real, M.D.; Rie, J.V.; Jansens, S.; Peferoen, M. Resistance to the Bacillus thuringiensis bioinsecticide in a field population of Plutella xylostella is due to a change in a midgut membrane receptor. Proc. Natl. Acad. Sci. USA 1991, 88, 5119–5123. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Z.; Liang, G.M.; Zhang, J.; Wu, K.M.; Guo, Y.Y.; Rector, B.G. Proteomic analysis of novel Cry1Ac binding proteins in Helicoverpa armigera (Hübner). Arch. Insect Biochem. Physiol. 2010, 73, 61–73. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.; Qi, Z.; Zhang, J.; Wu, W. Separation of Binding Protein of Celangulin V from the Midgut of Mythimna separata Walker by Affinity Chromatography. Toxins 2015, 7, 1738-1748. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7051738
Lu L, Qi Z, Zhang J, Wu W. Separation of Binding Protein of Celangulin V from the Midgut of Mythimna separata Walker by Affinity Chromatography. Toxins. 2015; 7(5):1738-1748. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7051738
Chicago/Turabian StyleLu, Lina, Zhijun Qi, Jiwen Zhang, and Wenjun Wu. 2015. "Separation of Binding Protein of Celangulin V from the Midgut of Mythimna separata Walker by Affinity Chromatography" Toxins 7, no. 5: 1738-1748. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7051738