
Enterotoxin Gene Cluster-Encoded SEI and SElN from Staphylococcus aureus Isolates are Crucial for the Induction of Human Blood Cell Proliferation and Pathogenicity in Rabbits

Abstract
:1. Introduction
2. Results
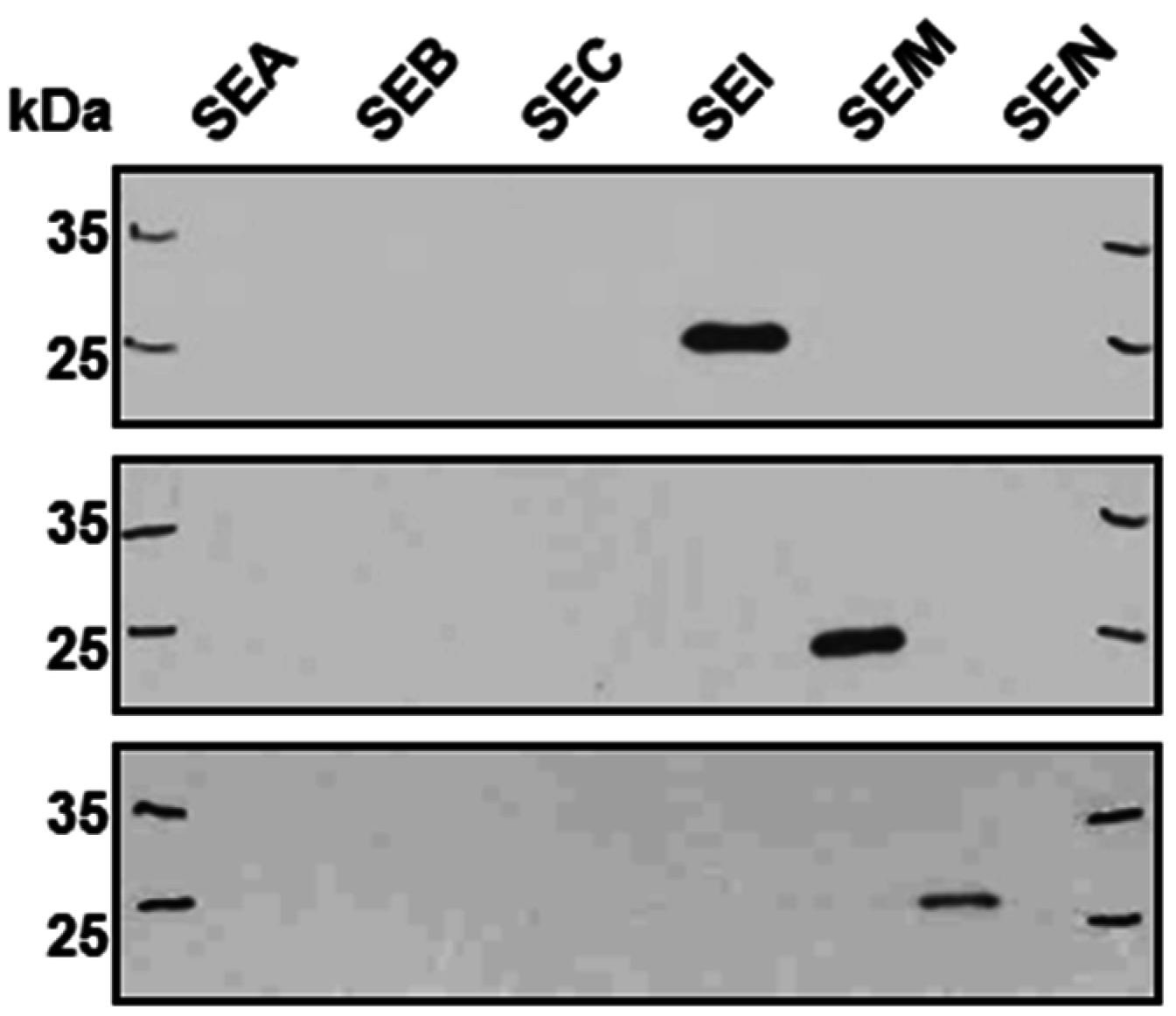
2.1. Superantigens of the egc Operon are Produced in Very Low Amounts
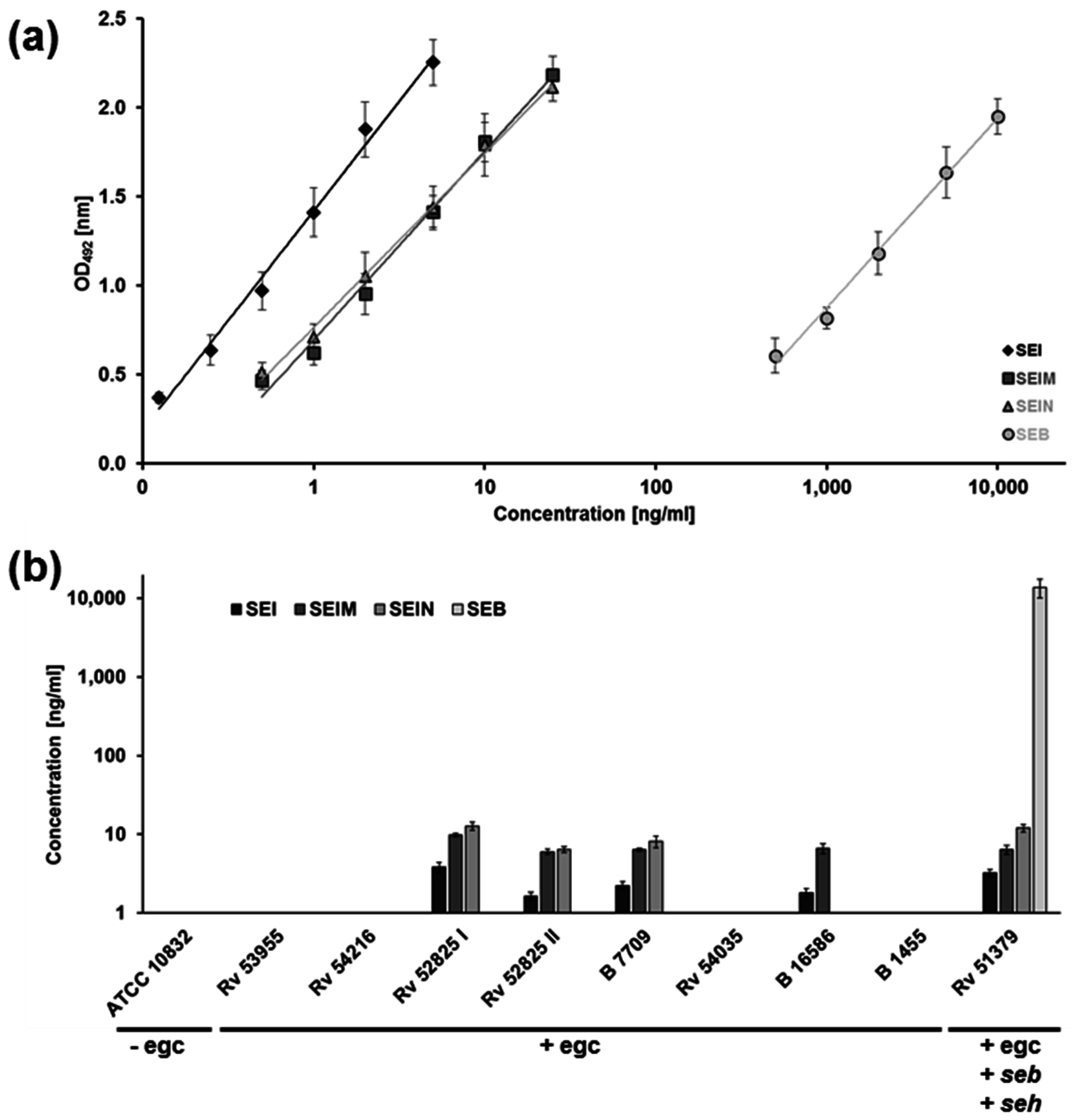
2.2. Similar SEB and egc Superantigen Concentrations Are Required for Immune Cell Activation
2.3. Amplification of LPS Toxicity In Vivo Can Be Demonstrated with Members of the egc Operon
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Extraction of RNA, and Isolation of Supernatants
4.2. Quantitative Real-Time PCR
4.3. Expression and Purification of Proteins
4.4. Enzyme-Linked Immunosorbent Assay
4.5. Lymphocyte Proliferation Assay
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SE | Staphylococcal enterotoxin |
| SEl | Staphylococcal enterotoxin-like |
| TSS | Toxic shock syndrome |
| MNC | Peripheral blood mononuclear cells |
| LPS | Lipopolysaccharide |
References
- Salgado-Pabón, W.; Breshears, L.; Spaulding, A.R.; Merriman, J.A.; Stach, C.S.; Horswill, A.R.; Peterson, M.L.; Schlievert, P.M. Superantigens are critical for Staphylococcus aureus Infective endocarditis, sepsis, and acute kidney injury. mBio 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Talan, D.A. Management of skin abscesses in the era of methicillin-resistant Staphylococcus aureus. N. Engl. J. Med. 2014, 370, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Peacock, S.J.; Moore, C.E.; Justice, A.; Kantzanou, M.; Story, L.; Mackie, K.; O’Neill, G.; Day, N.P.J. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect. Immun. 2002, 70, 4987–4996. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, T.; Preissner, K.T.; Herrmann, M. The anti-inflammatory activities of Staphylococcus aureus. Trends Immunol. 2007, 28, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Jupin, C.; Anderson, S.; Damais, C.; Alouf, J.E.; Parant, M. Toxic shock syndrome toxin 1 as an inducer of human tumor necrosis factors and gamma interferon. J. Exp. Med. 1988, 167, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Kulhankova, K.; King, J.; Salgado-Pabón, W. Staphylococcal toxic shock syndrome: Superantigen-mediated enhancement of endotoxin shock and adaptive immune suppression. Immunol. Res. 2014, 59, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Friedrich, A.W.; Lubritz, G.; Weilert, M.; Peters, G.; Von Eiff, C. Prevalence of genes encoding pyrogenic toxin superantigens and exfoliative toxins among strains of Staphylococcus aureus isolated from blood and nasal specimens. J. Clin. Microbiol. 2003, 41, 1434–1439. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.-L.; Omoe, K.; Inoue, F.; Kasai, T.; Yasujima, M.; Shinagawa, K.; Nakane, A. Comparative prevalence of superantigenic toxin genes in meticillin-resistant and meticillin-susceptible Staphylococcus aureus isolates. J. Med. Microbiol. 2008, 57, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Van Trijp, M.J.C.A.; Melles, D.C.; Snijders, S.V.; Wertheim, H.F.L.; Verbrugh, H.A.; van Belkum, A.; van Wamel, W.J. Genotypes, superantigen gene profiles, and presence of exfoliative toxin genes in clinical methicillin-susceptible Staphylococcus aureus isolates. Diagn. Microbiol. Infect. Dis. 2010, 66, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.A.; Mendoza, M.C.; Méndez, F.J.; Martín, M.C.; Guerra, B.; Rodicio, M.R. Clonal complexes and diversity of exotoxin gene profiles in methicillin-resistant and methicillin-susceptible Staphylococcus aureus isolates from patients in a Spanish hospital. J. Clin. Microbiol. 2009, 47, 2097–2105. [Google Scholar] [CrossRef] [PubMed]
- Jarraud, S.; Peyrat, M.A.; Lim, A.; Tristan, A.; Bes, M.; Mougel, C.; Etienne, J.; Vandenesch, F.; Bonneville, M.; Lina, G. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J. Immunol. 2001, 166, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Munson, S.H.; Tremaine, M.T.; Betley, M.J.; Welch, R.A. Identification and characterization of staphylococcal enterotoxin types G and I from Staphylococcus aureus. Infect. Immun. 1998, 66, 3337–3348. [Google Scholar] [PubMed]
- Letertre, C.; Perelle, S.; Dilasser, F.; Fach, P. Identification of a new putative enterotoxin SEU encoded by the egc cluster of Staphylococcus aureus. J. Appl. Microbiol. 2003, 95, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.Y.; Jarraud, S.; Lemercier, B.; Cozon, G.; Echasserieau, K.; Etienne, J.; Gougeon, M.-L.; Lina, G.; Vandenesch, F. Staphylococcal enterotoxin-like toxins U2 and V, two new staphylococcal superantigens arising from recombination within the enterotoxin gene cluster. Infect. Immun. 2006, 74, 4724–4734. [Google Scholar] [CrossRef] [PubMed]
- Ferry, T.; Thomas, D.; Genestier, A.-L.; Bes, M.; Lina, G.; Vandenesch, F.; Etienne, J. Comparative prevalence of superantigen genes in Staphylococcus aureus isolates causing sepsis with and without septic shock. Clin. Infect. Dis. 2005, 41, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Jarraud, S.; Cozon, G.; Vandenesch, F.; Bes, M.; Etienne, J.; Lina, G. Involvement of enterotoxins G and I in staphylococcal toxic shock syndrome and staphylococcal scarlet fever. J. Clin. Microbiol. 1999, 37, 2446–2449. [Google Scholar] [PubMed]
- Morgan, W.R.; Caldwell, M.D.; Brady, J.M.; Stemper, M.E.; Reed, K.D.; Shukla, S.K. Necrotizing fasciitis due to a methicillin-sensitive Staphylococcus aureus isolate harboring an enterotoxin gene cluster. J. Clin. Microbiol. 2007, 45, 668–671. [Google Scholar] [CrossRef] [PubMed]
- Stach, C.S.; Vu, B.G.; Merriman, J.A.; Herrera, A.; Cahill, M.P.; Schlievert, P.M.; Salgado-Pabón, W. Novel Tissue Level Effects of the Staphylococcus aureus Enterotoxin Gene Cluster Are Essential for Infective Endocarditis. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Grumann, D.; Ruotsalainen, E.; Kolata, J.; Kuusela, P.; Järvinen, A.; Kontinen, V.P.; Bröker, B.M.; Holtfreter, S. Characterization of Infecting Strains and Superantigen-Neutralizing Antibodies in Staphylococcus aureus Bacteremia. Clin. Vaccine Immunol. 2011, 18, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Omoe, K.; Ishikawa, M.; Shimoda, Y.; Hu, D.-L.; Ueda, S.; Shinagawa, K. Detection of seg, seh, and sei genes in Staphylococcus aureus isolates and determination of the enterotoxin productivities of S. aureus isolates Harboring seg, seh, or sei genes. J. Clin. Microbiol. 2002, 40, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Florquin, S.; Goldman, M. Immunoregulatory mechanisms of T-cell-dependent shock induced by a bacterial superantigen in mice. Infect. Immun. 1996, 64, 3443–3445. [Google Scholar] [PubMed]
- Roetzer, A.; Haller, G.; Beyerly, J.; Geier, C.B.; Wolf, H.M.; Gruener, C.S.; Model, N.; Eibl, M.M. Genotypic and phenotypic analysis of clinical isolates of Staphylococcus aureus revealed production patterns and hemolytic potentials unlinked to gene profiles and source. BMC Microbiol. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M. Role of superantigens in human disease. J. Infect. Dis. 1993, 167, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Yarwood, J.M.; Leung, D.Y.; Schlievert, P.M. Evidence for the involvement of bacterial superantigens in psoriasis, atopic dermatitis, and Kawasaki syndrome. FEMS Microbiol. Lett. 2000, 192, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Johler, S.; Giannini, P.; Jermini, M.; Hummerjohann, J.; Baumgartner, A.; Stephan, R. Further Evidence for Staphylococcal Food Poisoning Outbreaks Caused by egc-Encoded Enterotoxins. Toxins 2015, 7, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Holtfreter, S.; Bauer, K.; Thomas, D.; Feig, C.; Lorenz, V.; Roschack, K.; Friebe, E.; Selleng, K.; Lövenich, S.; Greve, T.; et al. egc-Encoded superantigens from Staphylococcus aureus are neutralized by human sera much less efficiently than are classical staphylococcal enterotoxins or toxic shock syndrome toxin. Infect. Immun. 2004, 72, 4061–4071. [Google Scholar] [CrossRef] [PubMed]
- Burian, M.; Grumann, D.; Holtfreter, S.; Wolz, C.; Goerke, C.; Bröker, B.M. Expression of staphylococcal superantigens during nasal colonization is not sufficient to induce a systemic neutralizing antibody response in humans. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Grumann, D.; Scharf, S.S.; Holtfreter, S.; Kohler, C.; Steil, L.; Engelmann, S.; Hecker, M.; Völker, U.; Bröker, B.M. Immune cell activation by enterotoxin gene cluster (egc)-encoded and non-egc superantigens from Staphylococcus aureus. J. Immunol. 2008, 181, 5054–5061. [Google Scholar] [CrossRef] [PubMed]
- Derzelle, S.; Dilasser, F.; Duquenne, M.; Deperrois, V. Differential temporal expression of the staphylococcal enterotoxins genes during cell growth. Food Microbiol. 2009, 26, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Waclavicek, M.; Stich, N.; Rappan, I.; Bergmeister, H.; Eibl, M.M. Analysis of the early response to TSST-1 reveals Vbeta-unrestricted extravasation, compartmentalization of the response, and unresponsiveness but not anergy to TSST-1. J. Leukoc. Biol. 2009, 85, 44–54. [Google Scholar] [CrossRef] [PubMed]
- McCormick, J.K.; Yarwood, J.M.; Schlievert, P.M. Toxic shock syndrome and bacterial superantigens: An update. Annu. Rev. Microbiol. 2001, 55, 77–104. [Google Scholar] [CrossRef] [PubMed]
- Proft, T.; Fraser, J.D. Bacterial superantigens. Clin. Exp. Immunol. 2003, 133, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.M. Bacterial toxins: A table of lethal amounts. Microbiol. Rev. 1982, 46, 86–94. [Google Scholar] [PubMed]
- Spaulding, A.R.; Lin, Y.-C.; Merriman, J.A.; Brosnahan, A.J.; Peterson, M.L.; Schlievert, P.M. Immunity to Staphylococcus aureus secreted proteins protects rabbits from serious illnesses. Vaccine 2012, 30, 5099–5109. [Google Scholar] [CrossRef] [PubMed]
- Gampfer, J.M.; Samstag, A.; Waclavicek, M.; Wolf, H.M.; Eibl, M.M.; Gulle, H. Epitope mapping of neutralizing TSST-1 specific antibodies induced by immunization with toxin or toxoids. Vaccine 2002, 20, 3675–3684. [Google Scholar] [CrossRef]
- Dauwalder, O.; Thomas, D.; Ferry, T.; Debard, A.-L.; Badiou, C.; Vandenesch, F.; Etienne, J.; Lina, G.; Monneret, G. Comparative inflammatory properties of staphylococcal superantigenic enterotoxins SEA and SEG: Implications for septic shock. J. Leukoc. Biol. 2006, 80, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Nowrouzian, F.L.; Ali, A.; Badiou, C.; Dauwalder, O.; Lina, G.; Josefsson, E. Impacts of enterotoxin gene cluster-encoded superantigens on local and systemic experimental Staphylococcus aureus infections. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- De Punder, K.; Pruimboom, L. Stress induces endotoxemia and low-grade inflammation by increasing barrier permeability. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M. Enhancement of host susceptibility to lethal endotoxin shock by staphylococcal pyrogenic exotoxin type C. Infect. Immun. 1982, 36, 123–128. [Google Scholar] [PubMed]
- Salgado-Pabón, W.; Schlievert, P.M. Models matter: The search for an effective Staphylococcus aureus vaccine. Nat. Rev. Microbiol. 2014, 12, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, A.R.; Salgado-Pabón, W.; Merriman, J.A.; Stach, C.S.; Ji, Y.; Gillman, A.N.; Peterson, M.L.; Schlievert, P.M. Vaccination against Staphylococcus aureus pneumonia. J. Infect. Dis. 2014, 209, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Staphylococcal Enterotoxins (-Like) |
|---|---|
| Rv53955 | sei, selm, seln, selo, selu, selx |
| Rv54216 | sei, selm, seln, selo, selu, selx |
| 784N-10 | seg, sei, selm, seln, selo |
| Rv51334 | seg, sei, selm, seln, selo, selx |
| Rv51410 | seg, sei, selm, seln, selo, selu |
| Rv51412 | seg, sei, selm, seln, selo, selu |
| Rv52825 I | seg, sei, selm, seln, selo, selx |
| Rv52825 II | seg, sei, selm, seln, selo |
| B7709 | seg, sei, selm, seln, selo, selu, selx |
| B8186 | seg, sei, selm, seln, selo, selu |
| Rv54035 | seg, sei, selm, seln, selo, selu, selx |
| B16586 | seg, sei, selm, seln, selo, selu |
| B1455 | seg, sei, selm, seln, selo, selw, selx |
| 803N-10 | seg, sei, selm, seln, selo, selu, selx |
| Rv51379 | seb, seg, seh, sei, selm, seln, selo, selu, selx |
| ATCC 10832 | selx |
| Sample | Conc. | SEI 1 | SElM 1 | SElN 1 |
|---|---|---|---|---|
| wt rSEI | 5 ng/mL | 4.67 ng/mL | 0.01 ng/mL | 0.02 ng/mL |
| 1 ng/mL | 0.85 ng/mL | 0.01 ng/mL | <0.01 ng/mL | |
| wt rSElM | 10 ng/mL | 0.01 ng/mL | 9.62 ng/mL | 0.01 ng/mL |
| 1 ng/mL | 0.01 ng/mL | 0.74 ng/mL | 0.01 ng/mL | |
| wt rSElN | 20 ng/mL | 0.01 ng/mL | <0.01 ng/mL | 17.89 ng/mL |
| 2 ng/mL | <0.01 ng/mL | <0.01 ng/mL | 1.53 ng/mL | |
| wt rSEB | 1 ng/mL | 0.01 ng/mL | <0.01 ng/mL | 0.04 ng/mL |
| wt rTSST1 | 1 ng/mL | 0.01 ng/mL | 0.01 ng/mL | 0.05 ng/mL |
| SE/Strain | Conc. | Vol. | Sera | LPS | No.1 |
|---|---|---|---|---|---|
| rSEB | 10 µg | 1 mL | - | 10 µg | 4/4 |
| rSEI | 10 µg | 1 mL | - | 10 µg | 2/2 |
| rSElN | 10 µg | 1 mL | - | 10 µg | 2/2 |
| Rv52825I | 7.7 ng/25.4 ng | 2 mL | - | 100 µg | 3/4 |
| Rv52825I | 7.7 ng/25.4 ng | 2 mL | anti-SEI/SElN | 100 µg | 0/4 |
| SE | Conc. | Vol. | LPS | No.1 |
|---|---|---|---|---|
| rSEB | 1 µg | 1 mL | 10 µg | 0/2 |
| rSEI | 10 ng | 1 mL | 10 µg | 2/2 |
| rSElN | 10 ng | 1 mL | 10 µg | 2/2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roetzer, A.; Gruener, C.S.; Haller, G.; Beyerly, J.; Model, N.; Eibl, M.M. Enterotoxin Gene Cluster-Encoded SEI and SElN from Staphylococcus aureus Isolates are Crucial for the Induction of Human Blood Cell Proliferation and Pathogenicity in Rabbits. Toxins 2016, 8, 314. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8110314
Roetzer A, Gruener CS, Haller G, Beyerly J, Model N, Eibl MM. Enterotoxin Gene Cluster-Encoded SEI and SElN from Staphylococcus aureus Isolates are Crucial for the Induction of Human Blood Cell Proliferation and Pathogenicity in Rabbits. Toxins. 2016; 8(11):314. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8110314
Chicago/Turabian StyleRoetzer, Andreas, Corina S. Gruener, Guenter Haller, John Beyerly, Nina Model, and Martha M. Eibl. 2016. "Enterotoxin Gene Cluster-Encoded SEI and SElN from Staphylococcus aureus Isolates are Crucial for the Induction of Human Blood Cell Proliferation and Pathogenicity in Rabbits" Toxins 8, no. 11: 314. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8110314