
Gastrointestinal Degradation of Fumonisin B1 by Carboxylesterase FumD Prevents Fumonisin Induced Alteration of Sphingolipid Metabolism in Turkey and Swine

Abstract
:1. Introduction
2. Results and Discussion
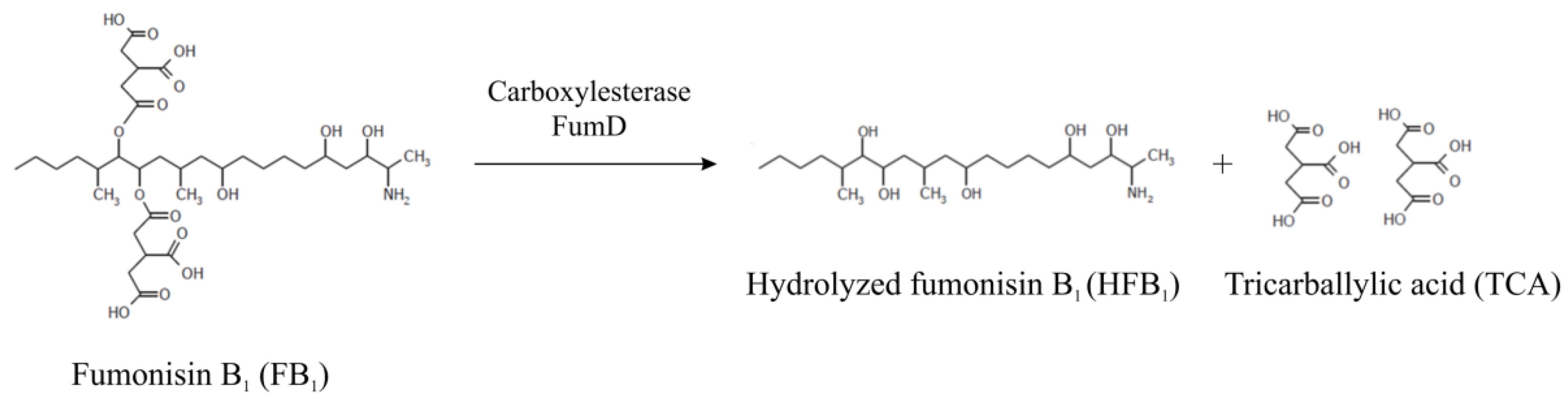
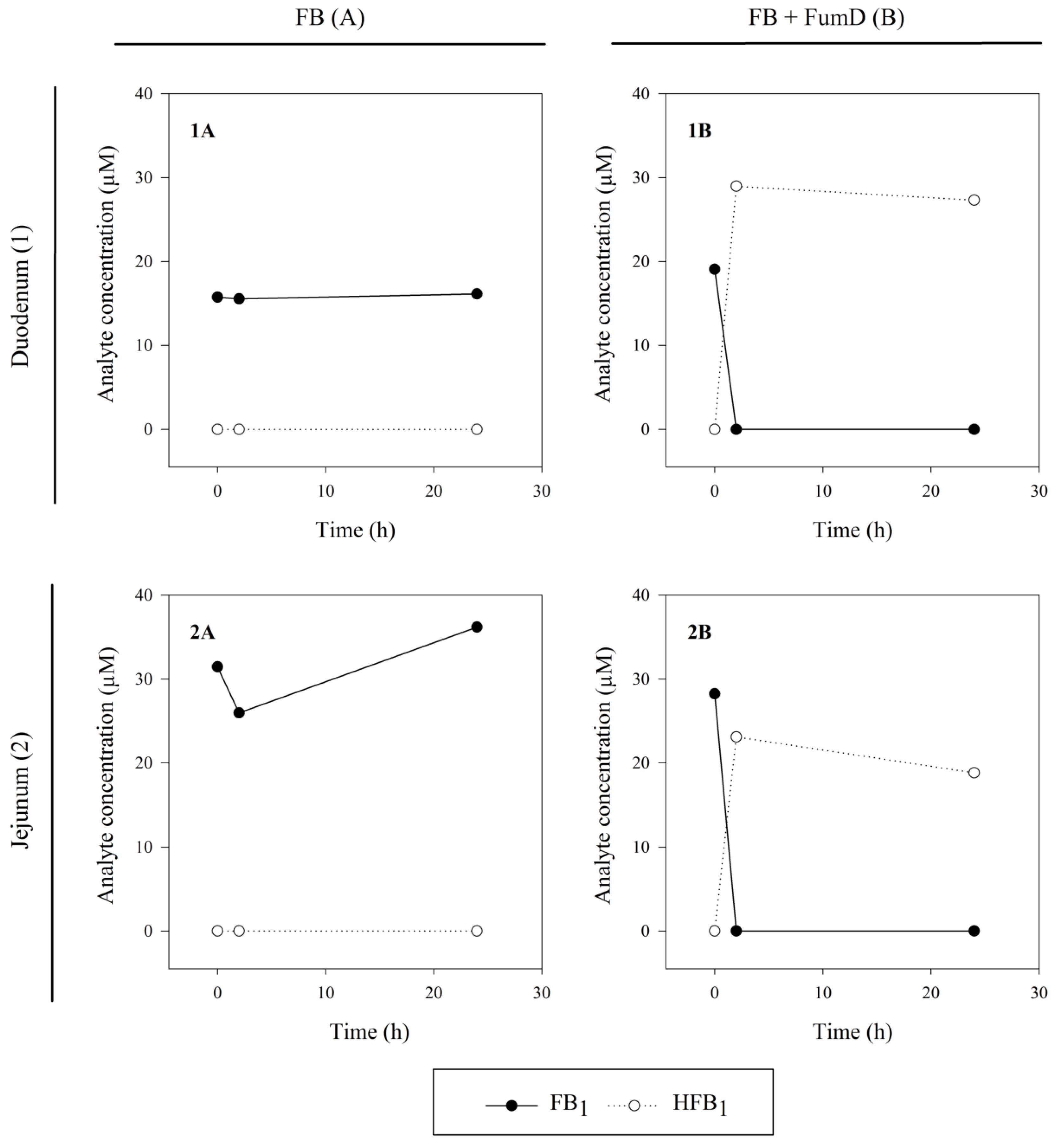
2.1. Enzyme Activity under Intestinal Conditions (Ex Vivo Model)
2.2. Efficacy of FUMzyme in Turkeys
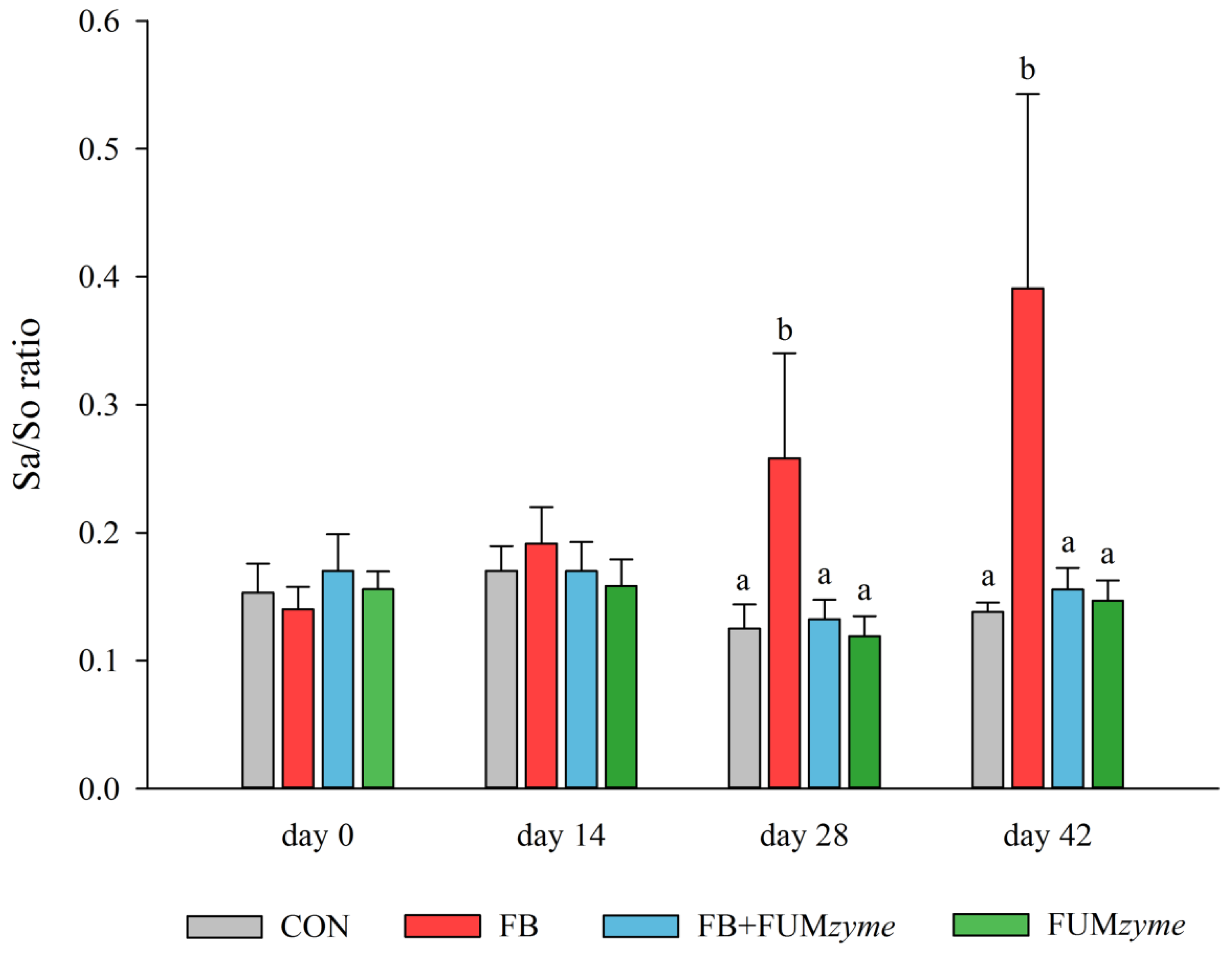
2.3. Efficacy of FUMzyme in Pigs
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Ex Vivo Model
3.2.1. Preparation of Enzyme and Toxin Solutions
3.2.2. Experimental Setup
3.2.3. Sample Clean-Up and Analysis of FB1 and HFB1 by Liquid Chromatography-Mass Spectrometry
3.3. In Vivo Trials
3.3.1. Animals and Study Design
3.3.2. Feed Composition
3.3.3. HPLC-MS/MS Analysis of Sphingolipids
3.3.4. HPLC-MS/MS Analysis of FB1, pHFB1a, pHFB1b, and HFB1
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| FB1 | fumonisin B1 |
| FB2 | fumonisin B2 |
| FB3 | fumonisin B3 |
| HFB1 | hydrolyzed fumonisin B1 |
| pHFB1a | partially hydrolyzed fumonisin B1a |
| pHFB1b | partially hydrolyzed fumonisin B1b |
| HPLC-MS/MS | high performance liquid chromatography-tandem mass spectrometry |
| LOD | limit of detection |
| LOQ | limit of quantification |
References
- Rheeder, J.P.; Marasas, W.F.; Vismer, H.F. Production of fumonisin analogs by fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.; Riley, R. Fumonisin toxicity and mechanism of action: Overview and current perspectives. Food Saf. 2013, 1, 2013006–2013006. [Google Scholar] [CrossRef]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: Facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Shier, W.T. The fumonisin paradox: A review of research on oral bioavailability of fumonisin b1, a mycotoxin produced by fusarium moniliforme. Toxin Rev. 2000, 19, 161–187. [Google Scholar] [CrossRef]
- Merrill, A.H., Jr.; van Echten, G.; Wang, E.; Sandhoff, K. Fumonisin b1 inhibits sphingosine (sphinganine) n-acyltransferase and de novo sphingolipid biosynthesis in cultured neurons in situ. J. Biol. Chem. 1993, 268, 27299–27306. [Google Scholar] [PubMed]
- Caloni, F.; Cortinovis, C. Effects of fusariotoxins in the equine species. Vet. J. 2010, 186, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Haschek, W.M.; Gumprecht, L.A.; Smith, G.; Tumbleson, M.E.; Constable, P.D. Fumonisin toxicosis in swine: An overview of porcine pulmonary edema and current perspectives. Environ. Health Perspect. 2001, 109 (Suppl. 2), 251–257. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vandenbroucke, V.; Li, S.; Haesebrouck, F.; van Immerseel, F.; Croubels, S. The impact of fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Marin, D.E.; Bouhet, S.; Pascale, F.; Bailly, J.D.; Miller, J.D.; Pinton, P.; Oswald, I.P. Mycotoxin fumonisin b1 alters the cytokine profile and decreases the vaccinal antibody titer in pigs. Toxicol. Sci. 2005, 84, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Tessari, E.N.C.; Oliveira, C.A.F.; Cardoso, A.; Ledoux, D.R.; Rottinghaus, G.E. Effects of aflatoxin b1 and fumonisin b1 on body weight, antibody titres and histology of broiler chicks. Br. Poult. Sci. 2006, 47, 357–364. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin a, t-2 and ht-2 and fumonisins in products intended for animal feeding (2006/576/ec). Off. J. Eur. Union 2006, L229, 7–9. [Google Scholar]
- Food and Drug Administration. FDA Mycotoxin Regulatory Guidance. Available online: https://www.ngfa.org/wp-content/uploads/NGFAComplianceGuide-FDARegulatoryGuidanceforMycotoxins8-2011.pdf (accessed on 30 March 2015).
- Grenier, B.; Loureiro-Bracarense, A.P.; Lucioli, J.; Pacheco, G.D.; Cossalter, A.M.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Individual and combined effects of subclinical doses of deoxynivalenol and fumonisins in piglets. Mol. Nutr. Food Res. 2011, 55, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Schwartz-Zimmermann, H.E.; Caha, S.; Moll, W.D.; Schatzmayr, G.; Applegate, T.J. Dose-dependent effects on sphingoid bases and cytokines in chickens fed diets prepared with fusarium verticillioides culture material containing fumonisins. Toxins 2015, 7, 1253–1272. [Google Scholar] [CrossRef] [PubMed]
- Wu, F. Measuring the economic impacts of fusarium toxins in animal feeds. Anim. Feed Sci. Technol. 2007, 137, 363–374. [Google Scholar] [CrossRef]
- Blandino, M.; Reyneri, A.; Colombari, G.; Pietri, A. Comparison of integrated field programmes for the reduction of fumonisin contamination in maize kernels. Field Crops Res. 2009, 111, 284–289. [Google Scholar] [CrossRef]
- Milani, J.; Maleki, G. Effects of processing on mycotoxin stability in cereals. J. Sci. Food Agric. 2014, 94, 2372–2375. [Google Scholar] [CrossRef] [PubMed]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’h, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. Part A 2011, 28, 1590–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugesan, G.; Ledoux, D.; Naehrer, K.; Berthiller, F.; Applegate, T.; Grenier, B.; Phillips, T.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, M.C.; Neeff, D.V.d.; Jager, A.V.; Corassin, C.H.; Carão, Á.C.d.P.; Albuquerque, R.d.; Azevedo, A.C.d.; Oliveira, C.A.F. Mineral adsorbents for prevention of mycotoxins in animal feeds. Toxin Rev. 2014, 33, 125–135. [Google Scholar] [CrossRef]
- McCormick, S.P. Microbial detoxification of mycotoxins. J. Chem. Ecol. 2013, 39, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Vekiru, E.; Krska, R.; Schatzmayr, G.; Moll, W.-D.; Grabherr, R. Degradation of fumonisin b1 by the consecutive action of two bacterial enzymes. J. Biotechnol. 2010, 145, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, D.; Schwartz, H.; Hametner, C.; Schatzmayr, G.; Haltrich, D.; Moll, W.-D. Enzyme characteristics of aminotransferase fumi of sphingopyxis sp. Mta144 for deamination of hydrolyzed fumonisin b1. Appl. Microbiol. Biotechnol. 2011, 91, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, D.; Moll, W. Fumonisin elimination and prospects for detoxification by enzymatic transformation. World Mycotoxin J. 2011, 4, 271–283. [Google Scholar] [CrossRef]
- Collins, T.F.; Sprando, R.L.; Black, T.N.; Olejnik, N.; Eppley, R.M.; Shackelford, M.E.; Howard, P.C.; Rorie, J.I.; Bryant, M.; Ruggles, D.I. Effects of aminopentol on in utero development in rats. Food Chem. Toxicol. 2006, 44, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Howard, P.C.; Couch, L.H.; Patton, R.E.; Eppley, R.M.; Doerge, D.R.; Churchwell, M.I.; Marques, M.M.; Okerberg, C.V. Comparison of the toxicity of several fumonisin derivatives in a 28-day feeding study with female B6C3F1 mice. Toxicol. Appl. Pharmacol. 2002, 185, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Riley, R.T.; Snook, M.E.; Gelineau-van Waes, J. Reproductive and sphingolipid metabolic effects of fumonisin b1 and its alkaline hydrolysis product in lm/bc mice: Hydrolyzed fumonisin b1 did not cause neural tube defects. Toxicol. Sci. 2009, kfp215. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Bracarense, A.-P.F.L.; Schwartz, H.E.; Trumel, C.; Cossalter, A.-M.; Schatzmayr, G.; Kolf-Clauw, M.; Moll, W.-D.; Oswald, I.P. The low intestinal and hepatic toxicity of hydrolyzed fumonisin b1 correlates with its inability to alter the metabolism of sphingolipids. Biochem. Pharmacol. 2012, 83, 1465–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracarense, A.P.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, S.; Hourcade, E.; Loiseau, N.; Fikry, A.; Martinez, S.; Roselli, M.; Galtier, P.; Mengheri, E.; Oswald, I.P. The mycotoxin fumonisin b1 alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2004, 77, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Basso, K.; Gomes, F.; Bracarense, A.P.L. Deoxynivanelol and fumonisin, alone or in combination, induce changes on intestinal junction complexes and in e-cadherin expression. Toxins 2013, 5, 2341–2352. [Google Scholar] [CrossRef] [PubMed]
- Devriendt, B.; Gallois, M.l.; Verdonck, F.; Wache, Y.; Bimczok, D.; Oswald, I.P.; Goddeeris, B.M.; Cox, E. The food contaminant fumonisin b1 reduces the maturation of porcine cd11r1+ intestinal antigen presenting cells and antigen-specific immune responses, leading to a prolonged intestinal etec infection. Vet. Res. 2009, 40, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P.; Lessard, M.; Oswald, I.P.; David, J.-C. Consumption of fumonisin b 1 for 9 days induces stress proteins along the gastrointestinal tract of pigs. Toxicon 2010, 55, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Burel, C.; Tanguy, M.; Guerre, P.; Boilletot, E.; Cariolet, R.; Queguiner, M.; Postollec, G.; Pinton, P.; Salvat, G.; Oswald, I.P. Effect of low dose of fumonisins on pig health: Immune status, intestinal microbiota and sensitivity to salmonella. Toxins 2013, 5, 841–864. [Google Scholar] [CrossRef] [PubMed]
- Oswald, I.P.; Desautels, C.; Laffitte, J.; Fournout, S.; Peres, S.Y.; Odin, M.; Le Bars, P.; Le Bars, J.; Fairbrother, J.M. Mycotoxin fumonisin b1 increases intestinal colonization by pathogenic escherichia coli in pigs. Appl. Environ. Microbiol. 2003, 69, 5870–5874. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Bracarense, A.-P.F.L.; Schwartz, H.E.; Lucioli, J.; Cossalter, A.-M.; Moll, W.-D.; Schatzmayr, G.; Oswald, I.P. Biotransformation approaches to alleviate the effects induced by fusarium mycotoxins in swine. J. Agric. Food Chem. 2013, 61, 6711–6719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benlasher, E.; Geng, X.; Xuan Nguyen, N.T.; Tardieu, D.; Bailly, J.-D.; Auvergne, A.; Guerre, P. Comparative effects of fumonisins on sphingolipid metabolism and toxicity in ducks and turkeys. Avian Dis. 2012, 56, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Shephard, G.S.; Thiel, P.G.; Sydenham, E.W.; Savard, M.E. Fate of a single dose of 14c-labelled fumonisin b1 in vervet monkeys. Nat. Toxins 1995, 3, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Fodor, J.; Balogh, K.; Weber, M.; Mezes, M.; Kametler, L.; Posa, R.; Mamet, R.; Bauer, J.; Horn, P.; Kovacs, F. Absorption, distribution and elimination of fumonisin b1 metabolites in weaned piglets. Food Addit. Contam. 2008, 25, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Hahn, I.; Nagl, V.; Schwartz-Zimmermann, H.E.; Varga, E.; Schwarz, C.; Slavik, V.; Reisinger, N.; Malachová, A.; Cirlini, M.; Generotti, S. Effects of orally administered fumonisin B1 (FB1), partially hydrolysed FB1, hydrolysed FB1 and N-(1-deoxy-d-fructos-1-yl) FB1 on the sphingolipid metabolism in rats. Food Chem. Toxicol. 2015, 76, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Guerre, P. Fusariotoxins in avian species: Toxicokinetics, metabolism and persistence in tissues. Toxins 2015, 7, 2289–2305. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Bailly, J.-D.; Skiba, F.; Grosjean, F.; Guerre, P. Toxicokinetics of fumonisin B1 in turkey poults and tissue persistence after exposure to a diet containing the maximum european tolerance for fumonisins in avian feeds. Food Chem. Toxicol. 2008, 46, 3213–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weibking, T.S.; Ledoux, D.R.; Brown, T.P.; Rottinghaus, G.E. Fumonisin toxicity in turkey poults. J. Vet. Diagn. Investig. 1993, 5, 75–83. [Google Scholar] [CrossRef]
- Li, Y.C.; Ledoux, D.R.; Bermudez, A.J.; Fritsche, K.L.; Rottinghaus, G.E. The individual and combined effects of fumonisin B1 and moniliformin on performance and selected immune parameters in turkey poults. Poult. Sci. 2000, 79, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, A.J.; Ledoux, D.R.; Turk, J.R.; Rottinghaus, G.E. The chronic effects of fusarium moniliforme culture material, containing known levels of fumonisin B1, in turkeys. Avian Dis. 1996, 231–235. [Google Scholar] [CrossRef]
- Ledoux, D.; Bermudez, A.; Rottinghaus, G. Effects of feeding fusarium moniliforme culture material, containing known levels of fumonisin B1, in the young turkey poult. Poult. Sci. 1996, 75, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Broomhead, J.N.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Chronic effects of fumonisin B1 in broilers and turkeys fed dietary treatments to market age. Poult. Sci. 2002, 81, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Bailly, J.D.; Skiba, F.; Metayer, J.P.; Grosjean, F.; Guerre, P. Chronic toxicity of fumonisins in turkeys. Poult. Sci. 2007, 86, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Fodor, J.; Meyer, K.; Gottschalk, C.; Mamet, R.; Kametler, L.; Bauer, J.; Horn, P.; Kovacs, F.; Kovacs, M. In vitro microbial metabolism of fumonisin B1. Food Addit. Contam. 2007, 24, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.T.; An, N.-H.; Showker, J.L.; Yoo, H.-S.; Norred, W.P.; Chamberlain, W.J.; Wang, E.; Merrill, A.H.; Motelin, G.; Beasley, V.R. Alteration of tissue and serum sphinganine to sphingosine ratio: An early biomarker of exposure to fumonisin-containing feeds in pigs. Toxicol. Appl. Pharmacol. 1993, 118, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Zimmermann, H.E.; Hahn, I.; Moll, W.-D.; Grenier, B.; Nagl, V.; Hartinger, D.; Slavik, V.; Caha, S.G.O.; Krska, R.; Berthiller, F. Analytical and biological validation of methods for determining fumonisin biomarkers in pigs, chicken and rats. Manuscript in preparation.
- European Commission. Directive 2010/63/eu of the european parliament and of the council of of 22 september 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, L 276/33, 1–47. [Google Scholar]
- European Union Reference Laboratory. Evaluation Report on the Analytical Methods Submitted in Connection with the Application for Authorisation of a Feed Additive According to Regulation (ec) No 1831/2003. Available online: https://ec.europa.eu/jrc/sites/default/files/FinRep-FAD-2013-0002-FUMzyme.doc_.pdf (accessed on 13 August 2015).
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Day | Treatment Group (n = 5) | FB1 ± SD (ng/g) | pHFB1a ± SD (ng/g) | pHFB1b ± SD (ng/g) | HFB1 ± SD (ng/g) |
|---|---|---|---|---|---|
| 0 | CON | <LOD | <LOD | <LOD | <LOD |
| FB | <LOD | <LOD | <LOD | <LOD | |
| FB+FUMzyme | <LOD | <LOD | <LOD | <LOD | |
| 7 | CON | <LOD | <LOD | <LOD | <LOD |
| FB | 2690 ± 1470 | 40.8 ± 32.3 | <LOQ | 194 ± 54.2 a | |
| FB+FUMzyme | 1450 ± 929 | 62.4 ± 46.1 | 308 ± 248 | 1340 ± 382 b | |
| 14 | CON | <LOD | <LOD | <LOD | <LOD |
| FB | 5240 ± 1930 a | <LOD | <LOD | <LOD | |
| FB+FUMzyme | 1190 ± 652 b | <LOD | 407 ± 380 | 1650 ± 368 |
| Treatment Group | Sa ± SD (ng/mL) | So ± SD (ng/mL) | Sa/So ± SD |
|---|---|---|---|
| CON | 6.61 ± 2.51 | 42.1 ± 17.4 | 0.16 ± 0.02 a |
| FB | 8.03 ± 1.31 | 34.5 ± 7.29 | 0.24 ± 0.02 b |
| FB+FUMzyme | 8.00 ± 3.49 | 41.4 ± 17.4 | 0.19 ± 0.02 a |
| Day | Treatment Group (n = 10) | FB1 ± SD (ng/g) | pHFB1a ± SD (ng/g) | pHFB1b ± SD (ng/g) | HFB1 ± SD (ng/g) |
|---|---|---|---|---|---|
| 0 | CON | <LOQ | <LOQ | <LOQ | <LOQ |
| FB | <LOQ | <LOQ | <LOQ | 184 ± 285 a | |
| FB+FUMzyme | <LOQ | <LOQ | 32.5 ± 17.2 | <LOQ | |
| FUMzyme | <LOQ | <LOD | <LOQ | 545 ± 548 b | |
| 14 | CON | 2350 ± 1960 a | 314 ± 174 a | 366 ± 221 a | 355 ± 190 a |
| FB | 6870 ± 815 b | 275 ± 153 a | 244 ± 177 a | 305 ± 225 a | |
| FB+FUMzyme | 1980 ± 394 a | 844 ± 223 b | 929 ± 246 b | 1820 ± 269 b | |
| FUMzyme | 1580 ± 609 a | < LOQ | 142 ± 41.8 a | 231 ± 72.1 a | |
| 28 | CON | <LOQ | <LOQ | <LOQ | <LOQ |
| FB | 11,900 ± 1300 a | <LOQ | 106 ± 26.5 a | <LOQ | |
| FB+FUMzyme | 2020 ± 442 b | 689 ± 201 | 703 ± 213 b | 1510 ± 212 | |
| FUMzyme | <LOQ | <LOQ | 122 ± 30.9 a | <LOQ | |
| 42 | CON (n = 9) | 3170 ± 235 a | <LOQ | 256 ± 48.2 a | <LOQ |
| FB | 14,900 ± 860 b | 252 ± 95.9 a | 326 ± 40.5 b | 349 ± 298 a | |
| FB+FUMzyme | 5650 ± 1390 c | 1170 ± 113 b | 983 ± 104 c | 1820 ± 293 b | |
| FUMzyme | 549 ± 322 d | <LOQ | 175 ± 26.5 d | 321 ± 153 a |
| Trial | Treatment Group | ∑ FB1+FB2 (mg/kg) | FUMzyme (U/kg) |
|---|---|---|---|
| Turkey | CON | - | - |
| FB | 15 | - | |
| FB+FUMzyme | 15 | 15 | |
| Pig | CON | - | - |
| FB | 2 | - | |
| FB+FUMzyme | 2 | 60 | |
| FUMzyme | - | 60 |
| Ingredient (%) | Turkey | Pig |
|---|---|---|
| Corn | 44.2 | 42.3 |
| Soybean meal | 31.0 | 24.0 |
| Wheat | 15.0 | 20.0 |
| Whey powder | - | 4.5 |
| Vegetable fat | - | 1.5 |
| Vegetable protein | - | 1.1 |
| Sunflower/palm kernel oil | 3.3 | 1.9 |
| Pumpkin seed cake | 0.7 | - |
| Lignocellulose | 0.6 | - |
| Calcium carbonate | 1.9 | 0.8 |
| Calcium phosphate | 1.8 | 1.4 |
| Natrium carbonate | 0.3 | - |
| Natrium chloride | 0.2 | 0.2 |
| Magnesium phosphate | 0.1 | - |
| Potassium diformiate | 0.1 | |
| Lysine | 0.3 | 0.6 |
| Methionine | 0.2 | 0.2 |
| Threonine | 0.1 | 0.2 |
| Tryptophan | - | 0.1 |
| Valine | - | 0.1 |
| Vitamin/mineral-premix | 0.3 1 | 1 2 |
| Analyzed Composition 3 | ||
| Crude protein (g) | 227 | 193 |
| Crude fibre (g) | 36 | 27 |
| Starch (g) | 441 | 515 |
| Metabolizable energy (MJ) | 14.2 | 16.2 |
| Matrix | Analyte | RA a ± RSD (%) | SSE b ± RSD (%) | RE c ± RSD (%) |
|---|---|---|---|---|
| Turkey serum | Sa | 93.4 ± 1.1 | 98.2 ± 2.7 | 95.2 ± 2.9 |
| So | 90.6 ± 0.5 | 93.4 ± 4.1 | 97.3 ± 4.5 | |
| Pig serum | Sa | 82.5 ± 0.5 | 96.7 ± 7.4 | 85.6 ± 7.7 |
| So | 79.9 ± 5.7 | 94.2 ± 5.9 | 85.2 ± 10.5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masching, S.; Naehrer, K.; Schwartz-Zimmermann, H.-E.; Sărăndan, M.; Schaumberger, S.; Dohnal, I.; Nagl, V.; Schatzmayr, D. Gastrointestinal Degradation of Fumonisin B1 by Carboxylesterase FumD Prevents Fumonisin Induced Alteration of Sphingolipid Metabolism in Turkey and Swine. Toxins 2016, 8, 84. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8030084
Masching S, Naehrer K, Schwartz-Zimmermann H-E, Sărăndan M, Schaumberger S, Dohnal I, Nagl V, Schatzmayr D. Gastrointestinal Degradation of Fumonisin B1 by Carboxylesterase FumD Prevents Fumonisin Induced Alteration of Sphingolipid Metabolism in Turkey and Swine. Toxins. 2016; 8(3):84. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8030084
Chicago/Turabian StyleMasching, Sabine, Karin Naehrer, Heidi-Elisabeth Schwartz-Zimmermann, Mihai Sărăndan, Simone Schaumberger, Ilse Dohnal, Veronika Nagl, and Dian Schatzmayr. 2016. "Gastrointestinal Degradation of Fumonisin B1 by Carboxylesterase FumD Prevents Fumonisin Induced Alteration of Sphingolipid Metabolism in Turkey and Swine" Toxins 8, no. 3: 84. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8030084