
The Kunitz-Type Protein ShPI-1 Inhibits Serine Proteases and Voltage-Gated Potassium Channels

 , ,
, ,
Abstract
:
1. Introduction
2. Results and Discussion
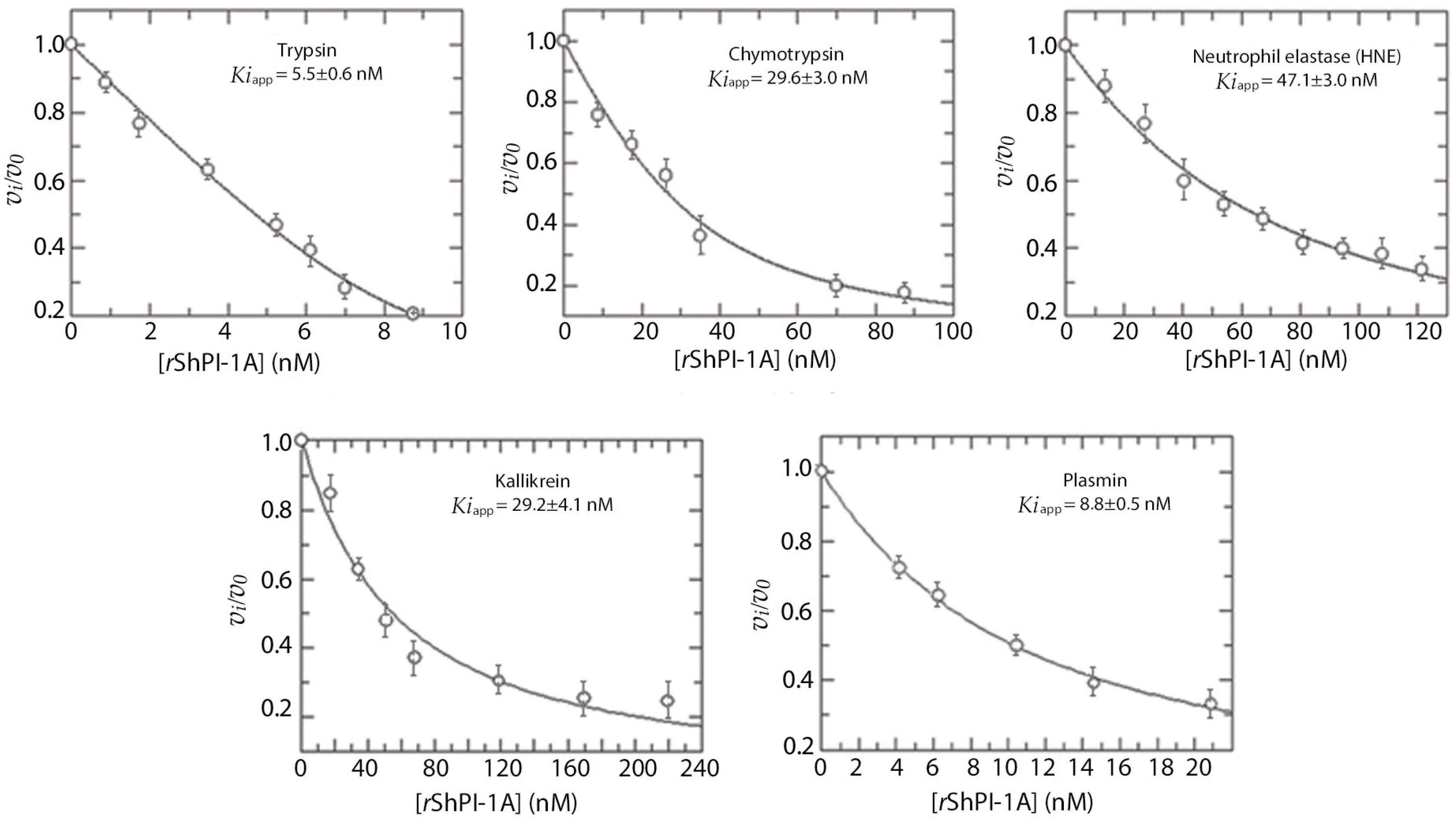
2.1. rShPI-1 Specificity and Dissociation Constants against Serine Proteases
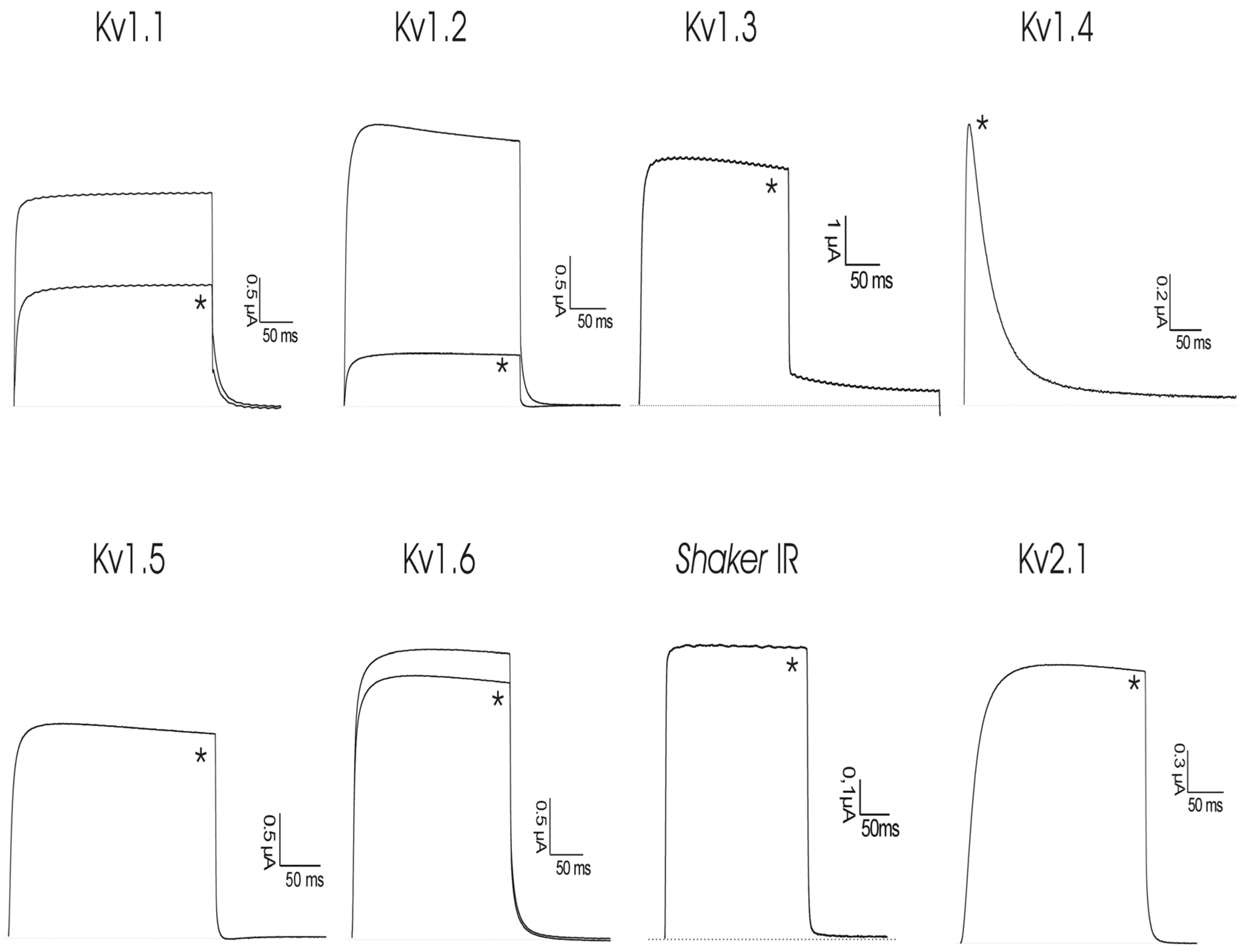
2.2. Activity toward Kv Channels: Electrophysiological Recordings
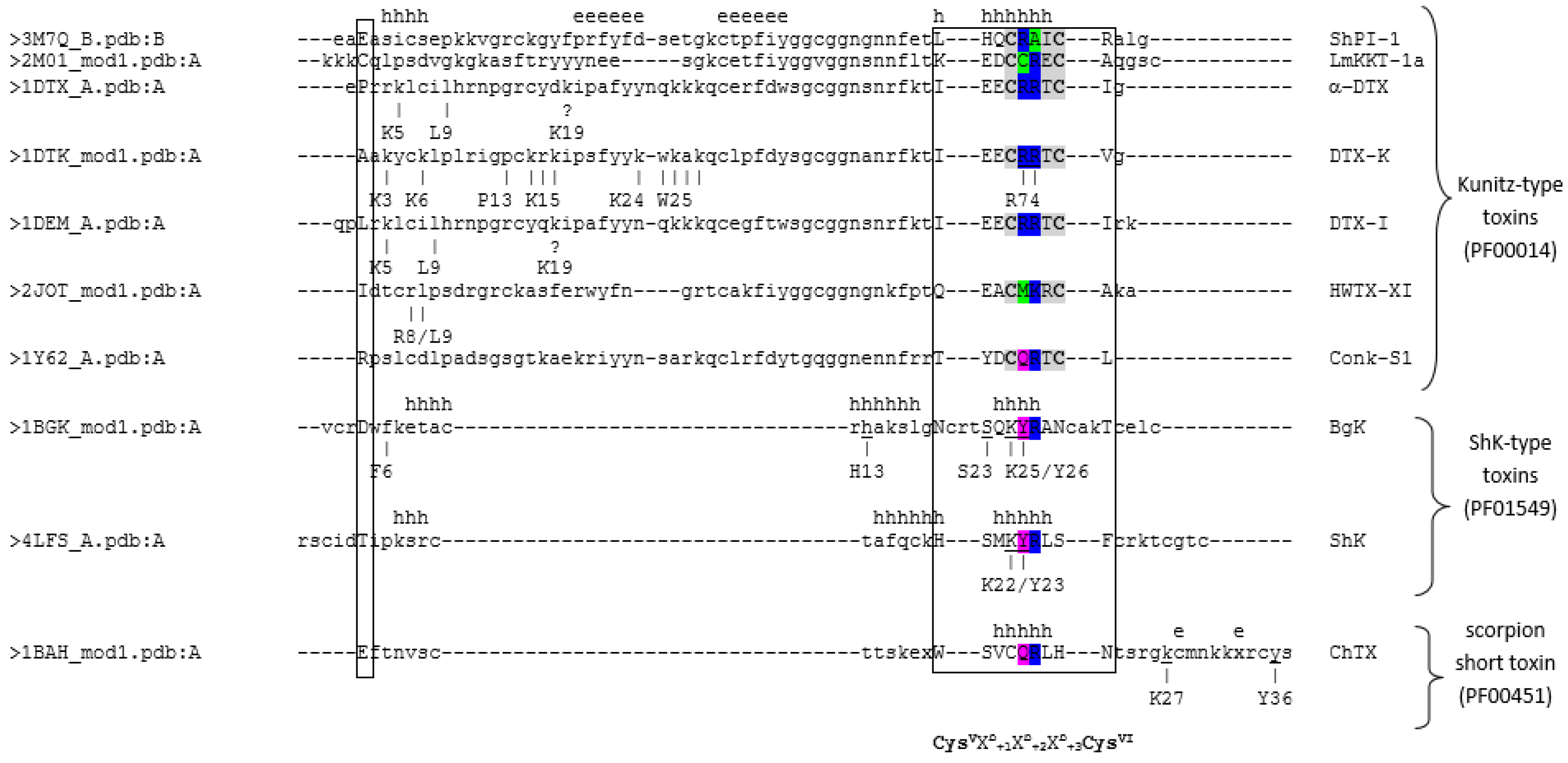
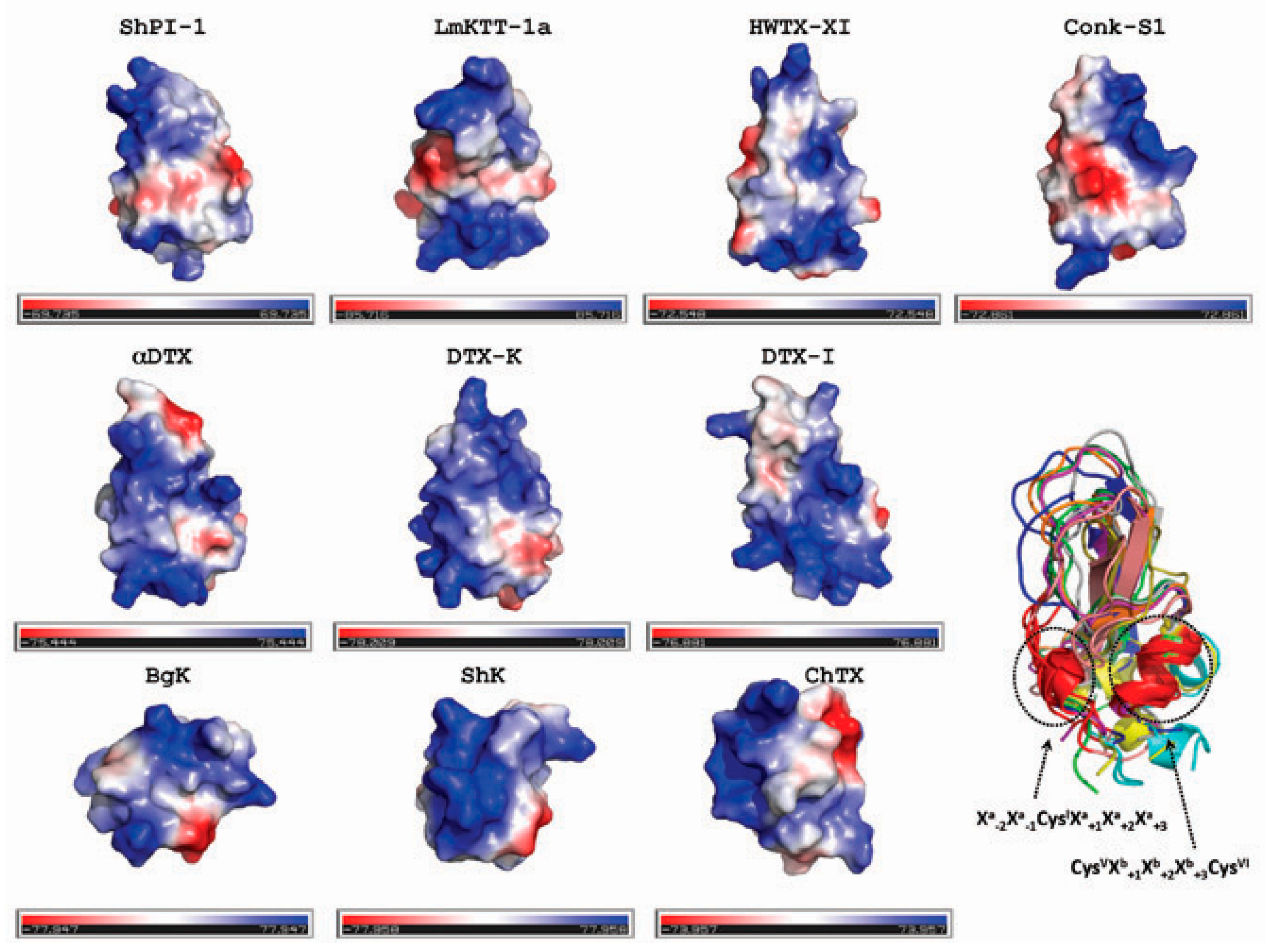
2.3. Sequence and Structural Comparison of ShPI-1 and Other Kv Channel Toxins
3. Materials and Methods
3.1. Protein Production and Purification
3.2. Protease Inhibition Studies
3.2.1. Inhibitory Specificity toward Serine Proteases
3.2.2. Determination of the Equilibrium Dissociation Constants (Ki)
3.3. Voltage-Gated Ion Channel Inhibition Experiments
3.3.1. Expression of Voltage-Gated Ion Channels in Xenopus laevis Oocytes
3.3.2. Electrophysiological Recordings
3.4. Computational Analyses
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 3D | three-dimensional |
| ASA | accessible surface area |
| BAPNA | N-benzoyl-arginine-p-nitroanilide or Bz-Arg-pNA |
| BPTI | bovine pancreatic trypsin inhibitor |
| DTX | dendrotoxin |
| Ki | equilibrium dissociation constant |
| Kv | voltage-gated potassium channel |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| ShPI-1 | Stichodactyla helianthus protease inhibitor 1 |
References
- Tabakmakher, V.M.; Sintsova, O.V.; Krivoshapko, O.N.; Zelepuga, E.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P. Analgesic effect of novel Kunitz-type polypeptides of the sea anemone Heteractis crispa. Dokl. Biochem. Biophys. 2015, 461, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Gladkikh, I.; Monastyrnaya, M.; Zelepuga, E.; Sintsova, O.; Tabakmakher, V.; Gnedenko, O.; Ivanov, A.; Hua, K.F.; Kozlovskaya, E. New Kunitz-type HCRG polypeptides from the sea anemone Heteractis crispa. Mar. Drugs 2015, 13, 6038–6063. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.A.; Salceda, E.; Garateix, A.G.; Zaharenko, A.J.; Peigneur, S.; Lopez, O.; Pons, T.; Richardson, M.; Diaz, M.; Hernandez, Y.; et al. A novel sea anemone peptide that inhibits acid-sensing ion channels. Peptides 2014, 53, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Frazao, B.; Vasconcelos, V.; Antunes, A. Sea anemone (cnidaria, anthozoa, actiniaria) toxins: An overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Gladkikh, I.; Monastyrnaya, M.; Leychenko, E.; Zelepuga, E.; Chausova, V.; Isaeva, M.; Anastyuk, S.; Andreev, Y.; Peigneur, S.; Tytgat, J.; et al. Atypical reactive center Kunitz-type inhibitor from the sea anemone Heteractis crispa. Mar. Drugs 2012, 10, 1545–1565. [Google Scholar] [CrossRef] [PubMed]
- García Fernández, R.; Reytor, M.L.; Alonso del Rivero, M.; Chávez, M.A. Estudios de relación estructura-función de inhibidores de proteasas tipo BPTI-Kunitz/Structure-function relationship of BPTI-Kunitz type protease inhibitors. Rev. Cuba. Cienc. Biol. 2015, 4, 2–19. (In Spanish) [Google Scholar]
- Delfin, J.; Martinez, I.; Antuch, W.; Morera, V.; Gonzalez, Y.; Rodriguez, R.; Marquez, M.; Saroyan, A.; Larionova, N.; Diaz, J.; et al. Purification, characterization and immobilization of proteinase inhibitors from Stichodactyla helianthus. Toxicon 1996, 34, 1367–1376. [Google Scholar] [CrossRef]
- Gil, D.F.; Garcia-Fernandez, R.; Alonso-del-Rivero, M.; Lamazares, E.; Perez, M.; Varas, L.; Diaz, J.; Chavez, M.A.; Gonzalez-Gonzalez, Y.; Mansur, M. Recombinant expression of ShPI-1A, a non-specific BPTI-Kunitz-type inhibitor, and its protection effect on proteolytic degradation of recombinant human miniproinsulin expressed in Pichia pastoris. FEMS Yeast Res. 2011, 11, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernandez, R.; Pons, T.; Meyer, A.; Perbandt, M.; Gonzalez-Gonzalez, Y.; Gil, D.; de los Angeles Chavez, M.; Betzel, C.; Redecke, L. Structure of the recombinant BPTI/Kunitz-type inhibitor rShPI-1A from the marine invertebrate Stichodactyla helianthus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012, 68, 1289–1293. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Nogueira, N.P.; Morgado-Diaz, J.A.; Menna-Barreto, R.F.; Paes, M.C.; da Silva-Lopez, R.E. Effects of a marine serine protease inhibitor on viability and morphology of Trypanosoma cruzi, the agent of chagas disease. Acta Trop. 2013, 128, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Silva-Lopez, R.E.; Morgado-Diaz, J.A.; Chavez, M.A.; Giovanni-de-Simone, S. Effects of serine protease inhibitors on viability and morphology of Leishmania (Leishmania) amazonensis promastigotes. Parasitol. Res. 2007, 101, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L. Recent studies on dendrotoxins and potassium ion channels. Gen. Pharmacol. 1997, 28, 7–12. [Google Scholar] [CrossRef]
- Harvey, A.L. Twenty years of dendrotoxins. Toxicon 2001, 39, 15–26. [Google Scholar] [CrossRef]
- Schweitz, H.; Bruhn, T.; Guillemare, E.; Moinier, D.; Lancelin, J.M.; Beress, L.; Lazdunski, M. Kalicludines and kaliseptine. Two different classes of sea anemone toxins for voltage sensitive K+ channels. J. Biol. Chem. 1995, 270, 25121–25126. [Google Scholar] [CrossRef] [PubMed]
- Honma, T.; Kawahata, S.; Ishida, M.; Nagai, H.; Nagashima, Y.; Shiomi, K. Novel peptide toxins from the sea anemone Stichodactyla haddoni. Peptides 2008, 29, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Billen, B.; Derua, R.; Waelkens, E.; Debaveye, S.; Beress, L.; Tytgat, J. A bifunctional sea anemone peptide with Kunitz type protease and potassium channel inhibiting properties. Biochem. Pharmacol. 2011, 82, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, O.; Sotolongo, V.; Amor, A.M.; Stocklin, R.; Anderson, A.J.; Harvey, A.L.; Engstrom, A.; Wernstedt, C.; Karlsson, E. Characterization of a potassium channel toxin from the caribbean sea anemone Stichodactyla helianthus. Toxicon 1995, 33, 603–613. [Google Scholar] [CrossRef]
- Pennington, M.W.; Chang, S.C.; Chauhan, S.; Huq, R.; Tajhya, R.B.; Chhabra, S.; Norton, R.S.; Beeton, C. Development of highly selective Kv1.3-blocking peptides based on the sea anemone peptide ShK. Mar. Drugs 2015, 13, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Beeton, C.; Pennington, M.W.; Norton, R.S. Analogs of the sea anemone potassium channel blocker ShK for the treatment of autoimmune diseases. Inflamm. Allergy Drug Targets 2011, 10, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R.; Parten, B.; Pennington, M.W.; Price, D.A.; Dunn, B.M. Isolation, characterization, and amino acid sequence of a polypeptide neurotoxin occurring in the sea anemone Stichodactyla helianthus. Biochemistry 1989, 28, 3483–3489. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.F. The slow-binding and slow, tight-binding inhibition of enzyme-catalysed reactions. Trends Biochem. Sci. 1982, 7, 102–105. [Google Scholar] [CrossRef]
- Bieth, J.G. Theoretical and practical aspects of proteinase inhibition kinetics. Methods Enzymol. 1995, 248, 59–84. [Google Scholar] [PubMed]
- Copeland, R.A.; Lombardo, D.; Giannaras, J.; Decicco, C.P. Estimating Ki values for tight binding inhibitors from dose-response plots. Bioorg. Med. Chem. Lett. 1995, 5, 1947–1952. [Google Scholar] [CrossRef]
- Garcia-Fernandez, R.; Pons, T.; Perbandt, M.; Valiente, P.A.; Talavera, A.; Gonzalez-Gonzalez, Y.; Rehders, D.; Chavez, M.A.; Betzel, C.; Redecke, L. Structural insights into serine protease inhibition by a marine invertebrate BPTI kunitz-type inhibitor. J. Struct. Biol. 2012, 180, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernandez, R.; Perbandt, M.; Rehders, D.; Ziegelmuller, P.; Piganeau, N.; Hahn, U.; Betzel, C.; Chavez Mde, L.; Redecke, L. Three-dimensional structure of a Kunitz-type inhibitor in complex with an elastase-like enzyme. J. Biol. Chem. 2015, 290, 14154–14165. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.F.; Wenzel, H.R.; Tschesche, H.R.; Colman, R.W. Kinetics of inhibition of human plasma kallikrein by a site-specific modified inhibitor Arg15-aprotinin: Evaluation using a microplate system and comparison with other proteases. Blood 1987, 69, 1431–1436. [Google Scholar] [PubMed]
- Czapinska, H.; Otlewski, J. Structural and energetic determinants of the S1-site specificity in serine proteases. Eur. J. Biochem. 1999, 260, 571–595. [Google Scholar] [CrossRef] [PubMed]
- Scheidig, A.J.; Hynes, T.R.; Pelletier, L.A.; Wells, J.A.; Kossiakoff, A.A. Crystal structures of bovine chymotrypsin and trypsin complexed to the inhibitor domain of alzheimer’s amyloid beta-protein precursor (APPI) and basic pancreatic trypsin inhibitor (BPTI): Engineering of inhibitors with altered specificities. Protein Sci. 1997, 6, 1806–1824. [Google Scholar] [CrossRef] [PubMed]
- Hernandez Gonzalez, J.E.; Garcia-Fernandez, R.; Valiente, P.A. Polar desolvation and position 226 of pancreatic and neutrophil elastases are crucial to their affinity for the Kunitz-type inhibitors ShPI-1 and ShPI-1/K13L. PLoS ONE 2015, 10, e0137787. [Google Scholar]
- Rodriguez, A.A.; Cassoli, J.S.; Sa, F.; Dong, Z.Q.; de Freitas, J.C.; Pimenta, A.M.; de Lima, M.E.; Konno, K.; Lee, S.M.; Garateix, A.; et al. Peptide fingerprinting of the neurotoxic fractions isolated from the secretions of sea anemones Stichodactyla helianthus and Bunodosoma granulifera. New members of the apetx-like family identified by a 454 pyrosequencing approach. Peptides 2012, 34, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Huerta, V.; Morera, V.; López, N.; Betancourt, L.; Besada, V.; Padrón, G.; Lima, Y.; Chávez, M.A.; Díaz, J. Characterization and 3D-model of a new protease inhibitor isolated from Stichodactyla helianthus. Biotechnol. Apl. 1998, 15, 108. [Google Scholar]
- Lanio, M.E.; Morera, V.; Alvarez, C.; Tejuca, M.; Gomez, T.; Pazos, F.; Besada, V.; Martinez, D.; Huerta, V.; Padron, G.; et al. Purification and characterization of two hemolysins from Stichodactyla helianthus. Toxicon 2001, 39, 187–194. [Google Scholar] [CrossRef]
- Alvarez, C.; Mancheno, J.M.; Martinez, D.; Tejuca, M.; Pazos, F.; Lanio, M.E. Sticholysins, two pore-forming toxins produced by the caribbean sea anemone Stichodactyla helianthus: Their interaction with membranes. Toxicon 2009, 54, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Robertson, B.; Owen, D.; Stow, J.; Butler, C.; Newland, C. Novel effects of dendrotoxin homologues on subtypes of mammalian Kv1 potassium channels expressed in xenopus oocytes. FEBS Lett. 1996, 383, 26–30. [Google Scholar] [CrossRef]
- Owen, D.G.; Hall, A.; Stephens, G.; Stow, J.; Robertson, B. The relative potencies of dendrotoxins as blockers of the cloned voltage-gated K+ channel, mKv1.1 (MK-1), when stably expressed in chinese hamster ovary cells. Br. J. Pharmacol. 1997, 120, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Hu, Y.T.; Yang, W.S.; He, Y.W.; Feng, J.; Wang, B.; Zhao, R.M.; Ding, J.P.; Cao, Z.J.; Li, W.X.; et al. Hg1, novel peptide inhibitor specific for Kv1.3 channels from first scorpion kunitz-type potassium channel toxin family. J. Biol. Chem. 2012, 287, 13813–13821. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.H.; He, Q.Y.; Peng, K.; Diao, J.B.; Jiang, L.P.; Tang, X.; Liang, S.P. Discovery of a distinct superfamily of Kunitz-type toxin (KTT) from tarantulas. PLoS ONE 2008, 3, e3414. [Google Scholar] [CrossRef]
- Bayrhuber, M.; Vijayan, V.; Ferber, M.; Graf, R.; Korukottu, J.; Imperial, J.; Garrett, J.E.; Olivera, B.M.; Terlau, H.; Zweckstetter, M.; et al. Conkunitzin-S1 is the first member of a new Kunitz-type neurotoxin family. Structural and functional characterization. J. Biol. Chem. 2005, 280, 23766–23770. [Google Scholar] [CrossRef] [PubMed]
- Finol-Urdaneta, R.K.; Becker, S.; Terlau, H.; French, R.J. Conkunitzins—Kv Channel Blockers with Serine Protease Inhibitor Activity. In Proceedings of the 90th Annual Meeting of The German Physiological Society, Regensburg, Germany, 26–29 March 2011; Acta Physiologica 2011. p. 284.
- Dy, C.Y.; Buczek, P.; Imperial, J.S.; Bulaj, G.; Horvath, M.P. Structure of Conkunitzin-S1, a neurotoxin and Kunitz-fold disulfide variant from cone snail. Acta Crystallogr. Sect. D Biol. Crystallogr. 2006, 62, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, S.; Danse, J.M.; Lecoq, A.; Pinkasfeld, S.; Zinn-Justin, S.; Young, L.C.; de Medeiros, C.C.; Rowan, E.G.; Harvey, A.L.; Menez, A. Delineation of the functional site of alpha-dendrotoxin. The functional topographies of dendrotoxins are different but share a conserved core with those of other Kv1 potassium channel-blocking toxins. J. Biol. Chem. 1998, 273, 25393–25403. [Google Scholar] [CrossRef] [PubMed]
- Scott, V.E.; Muniz, Z.M.; Sewing, S.; Lichtinghagen, R.; Parcej, D.N.; Pongs, O.; Dolly, J.O. Antibodies specific for distinct Kv subunits unveil a heterooligomeric basis for subtypes of alpha-dendrotoxin-sensitive K+ channels in bovine brain. Biochemistry 1994, 33, 1617–1623. [Google Scholar] [CrossRef] [PubMed]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: Waltham, MA, USA, 1965; pp. 97–166. [Google Scholar]
- Krissinel, E.; Henrick, K. Secondary-structure matching (SSM), a new tool for fast protein structure alignment in three dimensions. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2256–2268. [Google Scholar] [CrossRef] [PubMed]
- Dauplais, M.; Lecoq, A.; Song, J.; Cotton, J.; Jamin, N.; Gilquin, B.; Roumestand, C.; Vita, C.; de Medeiros, C.L.; Rowan, E.G.; et al. On the convergent evolution of animal toxins. Conservation of a diad of functional residues in potassium channel-blocking toxins with unrelated structures. J. Biol. Chem. 1997, 272, 4302–4309. [Google Scholar] [CrossRef] [PubMed]
- Bontems, F.; Gilquin, B.; Roumestand, C.; Menez, A.; Toma, F. Analysis of side-chain organization on a refined model of charybdotoxin: Structural and functional implications. Biochemistry 1992, 31, 7756–7764. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Chung, S.H. Computational studies of venom peptides targeting potassium channels. Toxins 2015, 7, 5194–5211. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, R.; Cohen, S.L.; Kuo, A.; Lee, A.; Chait, B.T. Structural conservation in prokaryotic and eukaryotic potassium channels. Science 1998, 280, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Shon, K.J.; Stocker, M.; Terlau, H.; Stuhmer, W.; Jacobsen, R.; Walker, C.; Grilley, M.; Watkins, M.; Hillyard, D.R.; Gray, W.R.; et al. Kappa-conotoxin PVIIa is a peptide inhibiting the shaker K+ channel. J. Biol. Chem. 1998, 273, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage t4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Erlanger, B.F.; Kokowsky, N.; Cohen, W. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys. 1961, 95, 271–278. [Google Scholar] [CrossRef]
- Claeson, G.; Aurell, L.; Friberger, P.; Gustavsson, S.; Karlsson, G. Designing of peptide substrates. Different approaches exemplified by new chromogenic substrates for kallikreins and urokinase. Haemostasis 1978, 7, 62–68. [Google Scholar] [PubMed]
- Mattler, L.E.; Bang, N.U. Serine protease specificity for peptide chromogenic substrates. Thromb. Haemost. 1977, 38, 776–792. [Google Scholar] [PubMed]
- Aurell, L.; Simonsson, R.; Arielly, S.; Karlsson, G.; Friberger, P.; Claeson, G. New chromogenic peptide substrates for factor Xa. Haemostasis 1978, 7, 92–94. [Google Scholar] [PubMed]
- Kisiel, W. Human plasma protein C: Isolation, characterization, and mechanism of activation by α-thrombin. J. Clin. Investig. 1979, 64, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Schechter, I.; Berger, A. On the size of the active site in proteases. I. Papain. Biochem. Biophys. Res. Commun. 1967, 27, 157–162. [Google Scholar] [CrossRef]
- Nakajima, K.; Powers, J.C.; Ashe, B.M.; Zimmerman, M. Mapping the extended substrate binding site of cathepsin g and human leukocyte elastase. Studies with peptide substrates related to the alpha 1-protease inhibitor reactive site. J. Biol. Chem. 1979, 254, 4027–4032. [Google Scholar] [PubMed]
- DelMar, E.G.; Largman, C.; Brodrick, J.W.; Geokas, M.C. A sensitive new substrate for chymotrypsin. Anal. Biochem. 1979, 99, 316–320. [Google Scholar] [CrossRef]
- Chase, T., Jr.; Shaw, E. P-nitrophenyl-p′-guanidinobenzoate HCl: A new active site titrant for trypsin. Biochem. Biophys. Res. Commun. 1967, 29, 508–514. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using clustal omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.; Chinea, G.; Lopez, N.; Pons, T.; Vriend, G. Homology modeling, model and software evaluation: Three related resources. Bioinformatics 1998, 14, 523–528. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | I0/E0 | Residual Activity (%) | Ki Values (nM) |
|---|---|---|---|
| Bovine pancreatic trypsin | 0.2 | 63 | (2.7 ± 0.3) |
| Human plasmin | 0.3 | 64 | (4.4 ± 0.2) |
| Porcine pancreatic kallikrein | 0.6 | 63 | (14.6 ± 2.00) |
| Human factor Xa | a | 100 | N.i b |
| Human α-thrombin | a | 100 | N.i b |
| Porcine pancreatic chymotrypsin | 0.9 | 52 | (14.8 ± 1.5) |
| Human neutrophil elastase | 0.6 | 66 | (23.5 ± 2.0) |
| Porcine pancreatic elastase | 1–170 | 100 | N.i b |
| B. licheniformis subtilisin | 1–240 | 100 | N.i b |
| Organism | Inhibitor | UniProt | Antiprotease Ki (nM) | Kv Inhibition a (IC50) | Reference |
|---|---|---|---|---|---|
| Stichodactyla helianthus | ShPI-1 | P31713 | See Table 1 | 117 ± 15 nM (Kv1.1) | this study |
| 9 ± 2 nM (Kv1.2) + (Kv 1.6) | |||||
| Anemonia sulcata | AsKC1 | Q9TWG0 | Tr: <30 | 2.8 µM/Kv1.2 | [14] |
| AsKC2 | Q9TWF9 | Tr: <30 | 1.1 µM/Kv1.2 | ||
| AsKC3 | Q9TWF8 | Tr: <30 | 1.3 µM/Kv1.2 | ||
| Anthopleura elegantissima | APEKTx1 | P86862 | Tr: 120 | 0.9 nM/Kv1.1 | [16] |
| Stichodactyla haddoni | SHTX-3 | B1B5l8 | Tr: 203 IU/mg | 0.65 mM b | [15] |
| (270 nM) c |
| Taxon | Toxin | UniProt | PDB | Target (IC50, nM) | Key Residues * | Reference |
|---|---|---|---|---|---|---|
| snakes | α-DTX | P00980 | 1DTX | Kv1.1 (1.1–150) | R3, R4, K5, L6, I8, L9 | [13] |
| Kv1.2 (0.4) | ||||||
| Kv 1.6 (9.0) | ||||||
| DTX-K | P00981 | 1DTK | Kv1.1 (2.5) (0.03) | K25, Y26, K28, P30, R32, P35, K37-K39, K46-K50, R74, R75 | [34,35] | |
| DTX-I | P00979 | 1DEM 1DEN | Kv1.1 (3.1) | K5, L9, K29 | [13] | |
| Kv1.2 (0.13) | ||||||
| Kv1.6; Kv1.3; Kv1.5 (weaker) | ||||||
| scorpion | Hg1 | P0C8WT | - | Kv1.3 (6.2) | C-terminal (K77, R78, F82, K84) | [36] |
| LmKTT-1a | P0DJ46 | 2M01 | Kv1.3 (>1000) | - | [36] | |
| LmKTT-1b | P0DJ45 | - | Kv1.3 (>1000) | - | [36] | |
| LmKTT-1c | P0DJ48 | - | Kv1.3 (>1000) | - | [36] | |
| BmKTT-1 | P0DJ49 | - | Kv1.3 (129.7) | - | [36] | |
| BmKTT-2 | P0DJ50 | - | Kv1.3 (371.3) | - | [36] | |
| BmKTT-3 | P9DJ47 | - | Kv1.3 (>1000) | - | [36] | |
| spider | HWTX-XI | P68425 | 2JOT | Kv1.1 (2.6 mM) | R38-L39 | [37] |
| Kv1.2; 1.3 (weaker) | ||||||
| Conus mollusk | Conk-S1 | P0C1X2 | 2CA7 | Kv1 (60) | No dyad needed; K47, R60 | [38,39,40] |
| 1Y62 | Kv1.7 (439) |
| Enzyme | [E0] 1 (M or U)2 | Substrate (Reference) | [S0] (mM) 2 | Buffer |
|---|---|---|---|---|
| trypsin | 2.2 10−7 | Bz-Arg-pNa [51] | 1.0 | 20 mM Tris-HCl; 150 mM NaCl; 20 mM CaCl2; pH 8.0 |
| kallikrein | 5.4 10−7 | HD-Val-Leu-Arg-pNA [52] | 0.5 | 50 mM Tris-HCl; 16 mM NaCl; pH 7.8 |
| plasmin | 2.4 10−8 | HD-Val-Leu-Lys-pNA [53] | 0.4 | 50 mM Tris-HCl; 110 mM NaCl; pH 7.4 |
| factor Xa | 0.1 U | Bz-Ile-Glu-Gly-Arg-pNA [54] | 0.4 | 50 mM Tris-HCl; 130 mM NaCl; 20 mM CaCl2; pH 8.3 |
| thrombin | 1.0 U | HD-Phe-Pip-Arg-pNA [55] | 0.4 | 50 mM Tris-HCl; 130 mM NaCl; 20 mM CaCl2; pH 8.3 |
| chymotryp. | 2.8 10−8 | Suc-Ala-Ala-Pro-Phe-pNA [56] | 1.0 | 50 mM Tris-HCl; pH 8.0 |
| neutrophil elastase 2 | 1.0 10−7 | MeOSuc-Ala-Ala-Pro-Val-pNA [57] | 0.14 | 20 mM Tris-HCl; 500 mM NaCl; pH 8 |
| pancreatic elastase | 3.3 10−8 | Suc-Ala-Ala-Ala-pNA [57] | 0.7 | 30 mM Sodium Phosphate; 50 mM NaCl; pH 7.0 |
| subtilisin A | 2.3 10−9 | Suc-Ala-Ala-Pro-Phe-pNA [58] | 0.12 | 100 mM Tris-HCl; 0.1% Triton X-100 ; pH 8.6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Fernández, R.; Peigneur, S.; Pons, T.; Alvarez, C.; González, L.; Chávez, M.A.; Tytgat, J. The Kunitz-Type Protein ShPI-1 Inhibits Serine Proteases and Voltage-Gated Potassium Channels. Toxins 2016, 8, 110. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8040110
García-Fernández R, Peigneur S, Pons T, Alvarez C, González L, Chávez MA, Tytgat J. The Kunitz-Type Protein ShPI-1 Inhibits Serine Proteases and Voltage-Gated Potassium Channels. Toxins. 2016; 8(4):110. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8040110
Chicago/Turabian StyleGarcía-Fernández, Rossana, Steve Peigneur, Tirso Pons, Carlos Alvarez, Lidice González, María A. Chávez, and Jan Tytgat. 2016. "The Kunitz-Type Protein ShPI-1 Inhibits Serine Proteases and Voltage-Gated Potassium Channels" Toxins 8, no. 4: 110. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8040110