2.1. Influence of Weight in the Processing of Samples
In order to establish the influence of the weight of the sample on the different steps of the sampling processing, we started by registering the whole weight as the sum of the shell, the water inside the shell and the flesh. We then determined the weight of the water inside the shell, a weight that is lost after opening the valves. According to the protocol, the sample to be taken for extraction, 2 g, must be obtained from a 100 g rinsed flesh homogenate. Therefore, after opening the shell, the flesh was weighed and then it was deposited in a sieve, rinsed with water and weighed again. In the case of steaming mussels, we weighed the amount of flesh and adsorbed water and the weight of the shell.
The mussels were manipulated in groups of 1 kg, and the weighing was repeated three times. The groups were: freshly obtained mussels, mussels steamed in the laboratory (to mimic a domestic cooking process), and mussels steamed in an industrial process (industrial steaming is the process utilized to separate the flesh from the shell). Mussels were from two different sources, Bueu and Riveira. Those from Bueu were large, with about 23 pieces in 1 kg (sample C) and with about 25 pieces in 1 kg (sample A), while mussels from Riveira were smaller, with 35 individuals per kg (sample B).
Table 1 shows the weights of fresh, laboratory-cooked (lab steaming) and industrial-steamed (for shucking) mussels from Riveira, which were small (35 individuals/kg, sample B), and from Bueu, which were large (23 mussels/kg, sample C), both with toxin values below legal limits. We also independently processed 25 large mussels from Bueu with a larger toxin concentration well above legal limits (sample A). In the industrial processing of mussels before steaming, only the initial weight of the whole mussels is known; hence, we could not calculate the percentage of the flesh weight.
The weight data show that, in general, in fresh mussels, the flesh represents between 22.5% ± 0.5% (SD) and 29.2% ± 0.3% (SD) of the total weight, and there is a significant amount of valve water that is lost after opening the mussel which does not contribute to the analysis but which it is considered in the weight of the original product, after debris is removed. We measured weights from 28.4% (
Table 1, sample B) to 38.6% (sample C), although very large mussels may have up to 45.9% of valve water (sample C). There is a small valve water loss before the shell is opened; this accounts for 7% (
Table 1, Sample 1), 6% (Sample 2), 7.3% (Sample 3) and 6.4% (Sample 4) of the initial weight. In mussels freshly obtained and processed in the factory, the amount of water in the valve is lower, with a value through several months (March to August 2014) of 23.9 ± 6.2 (SD). We can therefore assume that the amount of weight of valve water in fresh mussels accounts for between one-quarter to one-third of the total weight.
2.2. Influence of Steaming on Weight
Table 1 shows that after steaming, there is a loss of weight that does not depend on the size of the mussels or on the process, as laboratory steaming and industrial steaming provided very similar results. The maximum loss of weight due to steaming was an extra 9.5% of the initial weight (compared to fresh mussels in sample C, 13%
vs. 22.5%) when using industrial steaming, but in general steaming accounts for a small loss of weight. Therefore, laboratory steaming does not alter the values when compared to industrial steaming (19%
vs. 20.6% (sample A), 22.2%
vs. 21.5% (B) and 13%
vs. 14.6% (C)). The average weight of steamed flesh compared to the total mussel weight (discarding the valve water weight) in the industrial process from March to August 2014 was 18.2% ± 5.1% (SD). Also, the size of mussels does not modify the result, as small mussels lost 7.7% and 7% weight for laboratory and industrial steaming, respectively, while large mussels lost 7.9% and 9.5%, respectively.
2.3. Influence of Steaming on Hydrolysis
The hydrolysis of samples is necessary to calculate the amount of toxins in the form of the esters that may render OA, DTX1 or DTX2. This is necessary because DTX3 is not toxic, but its hydrolysis in the digestive system releases the active toxins that may cause intoxications [
7]. The samples used contained mainly OA esters, which is a typical toxin profile for Galician coasts with
Dynophisis acuminata and
Dynophisis acuta [
8,
9,
10]. The samples steamed in the industry were boiled for 2 min at 105 °C and then another 3 min at 100 °C. Also, the samples steamed in the laboratory mimicked a domestic cooking process, with water boiling for 10 min in a 5 L cooking pot with 100 mL of water. The results after hydrolysis show that mussels accumulate a rather important amount of DTX3 in the form of esters of OA and DTX2, although the largest amount of DTX3 corresponds to OA, with DTX2 esters being a maximum of 10% of the total DTX3 in the case of fresh mussels from the location of Riveira (
Table 2). There is a clear difference with regard to the source of mussels. The samples taken from Bueu contain no DTX2 in the form of esters. The percent of OA esters is also different depending on the location due to different exposure levels and the species of microalgae causing the bloom, with values that go from 38% to 82%. It is interesting that the percentage of esterified OA or DTX2 is different in the same sample, such as in the 35 fresh mussels sampled from Riveira, that show 81.6% and 48.6% esterified toxin for OA and DTX2, respectively. With regard to the initial toxin value, the highest increase in percent of OA or DTX2 after hydrolysis is 427.6% and 101.5%, respectively, in fresh product (sample B,
Table 2). This is attributable to mussels located in different places of the farming area, probably with different feeding and algae exposure conditions. The values for DTX2 are close to the limit of quantification (LOQ) (6.24 µg/kg), and therefore must be taken with caution.
In general, steaming increases the amount of toxins per kg (
Table 2), as in all cases the amount of OA/kg is higher after laboratory steaming and even higher after industrial steaming, the increases being 332 < 467 < 670, 14 < 20 < 21, and 80 < 171 < 187 for the levels of OA in the three samples. Total toxin shows similar values in all cases before and after steaming, although there is a slight decrease of total toxin after steaming in small mussels from Riveira, and a slight increase after steaming in large mussels from Bueu. The logical conclusion is that this increase in toxicity is caused by conversion of DTX3 to unesterified acid. In the case of mussels from Riveira, there is a slight decrease of total toxin.
It is a remarkable observation that in the case of the 1 kg sample from Bueu, fresh mussels evolved after steaming from below legal limits to above legal limits (160 μg of OA equivalents/kg), but this is attributable to a variation in the analytical results in different samples, as this was not observed in samples from Riveira, or in the set of 25 mussels with a higher concentration from Bueu.
2.4. Influence of Steaming and Autoclaving on DTX3 Levels with no Loss of Water
Table 2 shows the effect of steaming on the concentration of DTX3. The results show that although the total toxins remains the same, the amount of esterified toxins is greatly decreased, with the values being 203 > 35 > 0, 62 > 38 > 19, and 60 = 76 > 0 for the levels of OA-DTX3 in the three samples. Also, it is clear that the steaming in the industrial process is more aggressive to DTX3 than laboratory steaming. This destruction of DTX3 with heat may be related to the presence of the flesh. We have reported in a previous article that high pressure combined with an acidic pH can destroy OA [
11]. Since sterilization combines pressure with heat, we decided to compare the effect of steaming or autoclave treatment on DTX3 concentration. As a consequence, we studied the effect of sterilization (121 °C and 1.1 atm) in known amounts of DTX3 in a liquid-certified matrix that was not sterilized during the preparation process, using sealed vials to prevent any water loss by evaporation (
Table 3A). Also, we studied the effect of steaming in homogenous mussel tissue (Mussel_Control) with DTX3 levels in sealed vials. In the case of homogenous mussel tissue (Mussel_Control), we did not study the autoclave effect, since the preparation of the matrix involves a sterilization process, hence the matrix was already modified (
Table 3B).
There is a 59.4% decrease in the levels of DTX3 in the liquid-certified matrix (Mussel-DSP-2) after steaming for 10 min (
Table 3A). The results also show that sterilization does not decrease the level of DTX3 compared to the effect of steaming. Also, 20 min steaming does not further modify the levels of DTX3. On the contrary, steaming does not modify the levels of DTX3 in homogenous mussel tissue (Mussel_Control) (
Table 3B), but these results have to be taken with caution, as this matrix was treated with a sterilization process during the preparation. Taking into account the effect shown in
Table 3A, this process may have already decreased the original level of DTX3 to a minimum.
Although the changes in the DTX3 levels are quite clear in liquid matrix after 10 min steaming (
Table 3A), the total DTX3 does not change (669 µg/kg
vs. 539 µg/kg), but there is a rather significant decrease (about 50%) of total toxicity after the autoclave treatment (369 µg/kg
vs. 197 µg/kg or 669 µg/kg
vs. 327 µg/kg). These results are therefore in agreement with previous studies that reported a decrease in concentration after heat treatment for 10 min and 100 °C to 150 °C [
2], although we show a higher decrease of total toxicity after the autoclave treatment, and a slight toxicity reduction after steaming.
2.5. Influence of Matrix and Steaming on Pure DTX3 Stability with No Loss of Water
To understand the effect of heat in DTX3 stability, we used three pure DTX3 compounds, namely A (7-O-palmitoyl okadaic ester), B (7-O-palmytoleyl okadaic ester) and C (mixture of isomers A and B). The same amount was added to 2 mL of water, or 2 g of mussel tissue (Mussel_Control). These solutions were cooked for 10 min and 20 min in sealed vials to avoid water loss (control was at time 0). Although we did not know the initial amount of DTX3 analogues, we estimated the value after hydrolysis by measuring OA. The results, shown in
Table 4, indicate that pure DTX3 compound A is slightly degraded after steaming for 20 min (561 > 530 > 403 µg/kg), and compound B is further degraded by heat (535 > 460 > 338 µg/kg). A surprising result was that the mixture of esters in sample C was not modified by hydrolysis, which seems to suggest the toxin values were below the quantitation limit for sample C. The addition of the compounds to homogenous mussel tissue (Mussel_Control) seems to provide an environment which makes the compounds resistant to heat, as there is no decrease of the toxins. The conclusion to this is that the presence of matrix seems to stabilize DTX3 and makes the analogues more resistant to heat treatment.
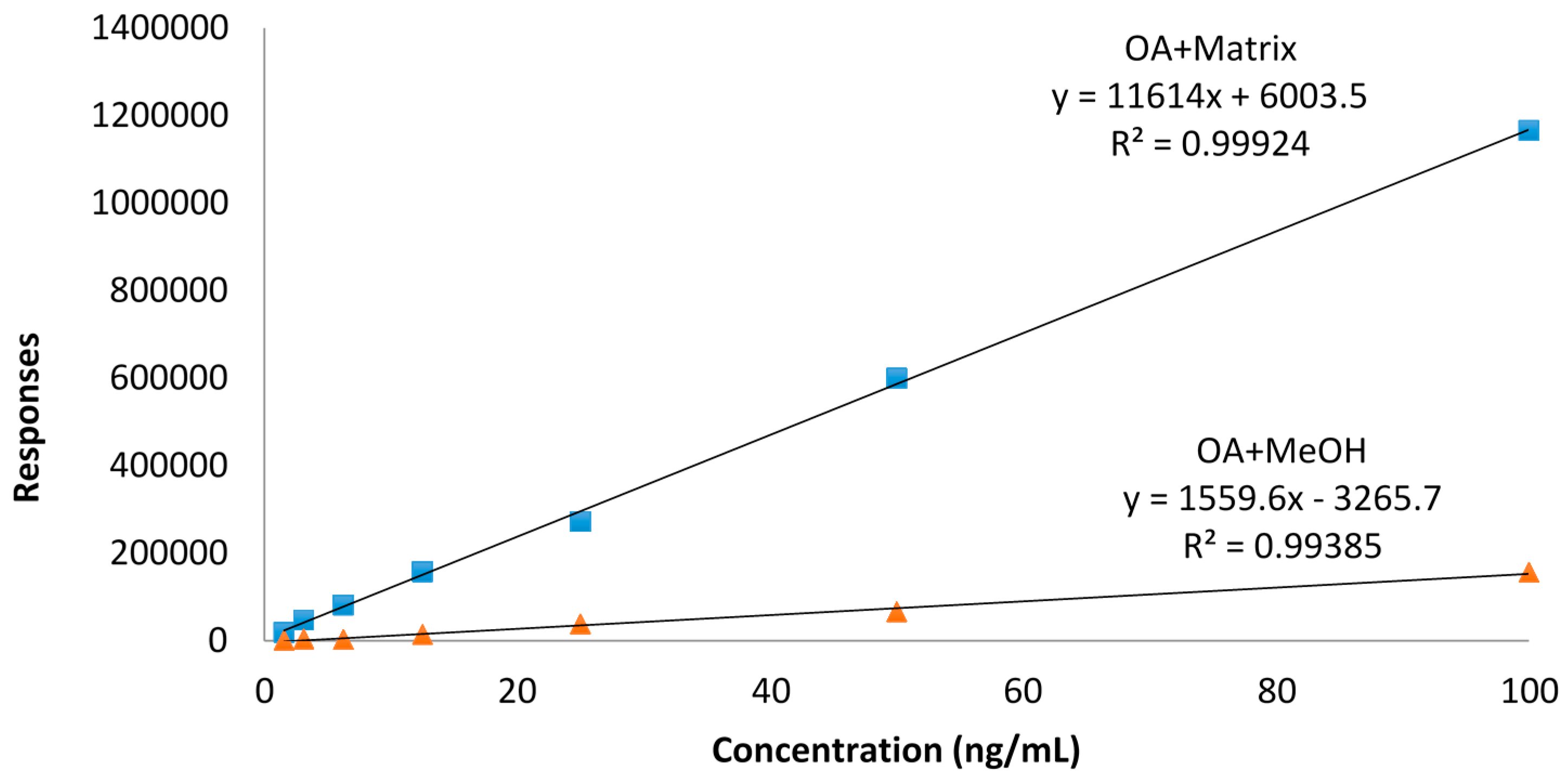
Our data are rather consistent in terms of toxin values, showing a decrease of toxins with higher temperatures. Although there are reports that describe unusual matrix effects that cause an increase in toxin concentrations of both OA, DTX2 [
12] or AZA3 [
13], we have not observed any of these effects, which rules out matrix effects other than those eliminated by the calibration procedure, as shown in
Figure 1.
The overall conclusion to our data is that DTX3 in seafood is destroyed by heat under steaming conditions, and that this destruction depends on the matrix environment. Sterilization does pose a stability issue to the toxins, as steaming treatment means a reduction in the amount of DTX3, but not a reduction in total toxicity, since OA or DTX2 are thermostable, but not at 121 °C, with a 50% decrease in total toxicity. The loss of water has been reported as a source of higher toxicity due to the increase of the concentration of toxins in the matrix [
1,
2], but our data indicates that this loss of water, once the water contained in the valves is discarded, is not higher than 7.8%. Our results do not sustain the observation reported earlier of a 30% water loss after steaming [
12]. The difference is attributable to the stronger conditions used, 130 °C for 70 s, while we used milder conditions, 105 °C for 120 s and then 100 °C for 180 s.
Under the mild industrial conditions we studied, it is clear that water loss is very small and that toxicity is unaltered, even though we show that DTX3 is destroyed at a certain level by heat, and largely (50%) by sterilization. Another technical aspect to bear in mind, although we have not studied it, is the reduction of water content after frying the flesh for the canning industry, where a 20% loss of water has to be added to the steaming process.







{kind=link}
{kind=link}