
The Sialidase NanS Enhances Non-TcsL Mediated Cytotoxicity of Clostridium sordellii

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Molecular and Genetic Techniques
2.3. Construction of a nanS-Targeted TargeTron and a C. sordellii ATCC9714 nanS Mutant
2.4. Complementation of the C. sordellii nanS Mutant
2.5. Sialidase Quantification Assays
2.6. Cell Cytotoxicity Assays
2.7. Cell Viability Assays
3. Results
3.1. Construction of a C. sordellii nanS Mutant
3.2. Complementation of the nanS Mutation
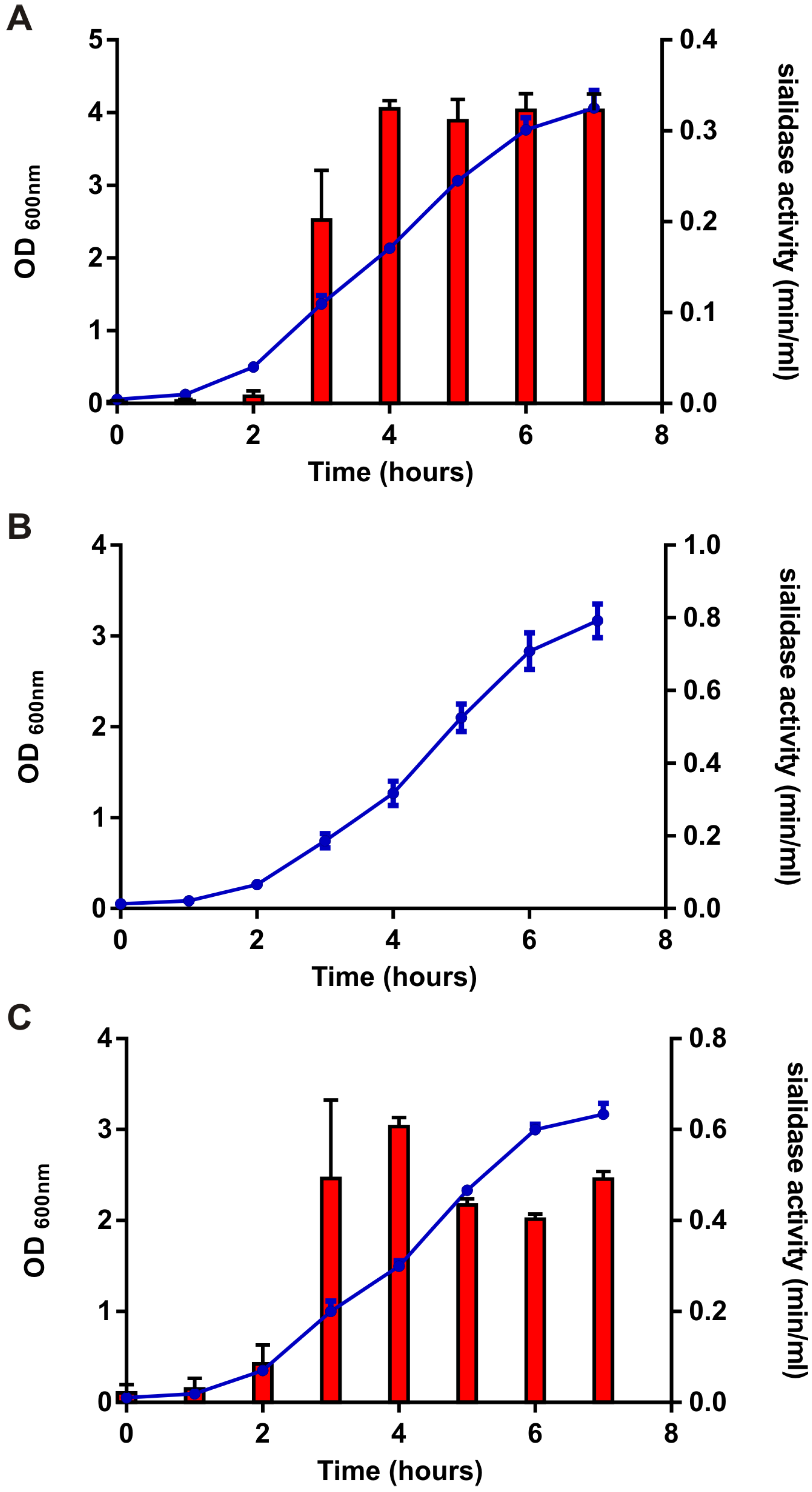
3.2.1. NanS is the Only Sialidase Produced by ATCC9714
3.2.2. NanS Is Not Cytotoxic to Eukaryotic Cells but Augments C. sordellii Cytotoxicity
3.2.3. C. sordellii Cytotoxicity Enhanced by NanS Is Not TcsL-Mediated and Results from the Action of an Unknown Toxin or Toxins
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aldape, M.J.; Bryant, A.E.; Ma, Y.; Stevens, D.L. The leukemoid reaction in Clostridium sordellii infection: Neuraminidase induction of promyelocytic cell proliferation. J. Infect. Dis. 2007, 195, 1838–1845. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Bhatnagar, J.; Guarner, J.; Reagan, S.; Hacker, J.K.; Van Meter, S.H.; Poukens, V.; Whiteman, D.B.; Iton, A.; Cheung, M.; et al. Fatal toxic shock syndrome associated with Clostridium sordellii after medical abortion. N. Engl. J. Med. 2005, 353, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Al-Mashat, R.R.; Taylor, D.J. Clostridium sordellii in enteritis in an adult sheep. Vet. Rec. 1983, 112, 19. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.J.; Naylor, R.D. Sudden death in sheep associated with Clostridium sordellii. Vet. Rec. 1998, 142, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Aldape, M.J.; Bryant, A.E.; Stevens, D.L. Clostridium sordellii infection: Epidemiology, clinical findings, and current perspectives on diagnosis and treatment. Clin. Infect. Dis. 2006, 43, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.; Garg, R. Fulminant leukemoid reaction due to postpartum Clostridium sordellii infection. J. Glob. Infect. Dis. 2012, 4, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Voth, D.E.; Martinez, O.V.; Ballard, J.D. Variations in lethal toxin and cholesterol-dependent cytolysin production correspond to differences in cytotoxicity among strains of Clostridium sordellii. FEMS Microbiol. Lett. 2006, 259, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Awad, M.M.; Hao, Y.; Thelen, T.; Bergin, I.L.; Howarth, P.M.; Seemann, T.; Rood, J.I.; Aronoff, D.M.; Lyras, D. TcsL is an essential virulence factor in Clostridium sordellii ATCC 9714. Infect. Immun. 2011, 79, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Geny, B.; Khun, H.; Fitting, C.; Zarantonelli, L.; Mazuet, C.; Cayet, N.; Szatanik, M.; Prevost, M.C.; Cavaillon, J.M.; Huerre, M.; et al. Clostridium sordellii lethal toxin kills mice by inducing a major increase in lung vascular permeability. Am. J. Pathol. 2007, 170, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Honma, K.; Mishima, E.; Sharma, A. Role of Tannerella forsythia NanH sialidase in epithelial cell attachment. Infect. Immun. 2011, 79, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Vimr, E.R.; Kalivoda, K.A.; Deszo, E.L.; Steenbergen, S.M. Diversity of microbial sialic acid metabolism. Microbiol. Mol. Biol. Rev. 2004, 68, 132–153. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Glycan-based interactions involving vertebrate sialic-acid-recognizing proteins. Nature 2007, 446, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Severi, E.; Hood, D.W.; Thomas, G.H. Sialic acid utilization by bacterial pathogens. Microbiology 2007, 153, 2817–2822. [Google Scholar] [CrossRef] [PubMed]
- Plumbridge, J.; Vimr, E. Convergent pathways for utilization of the amino sugars N-acetylglucosamine, N-acetylmannosamine, and N-acetylneuraminic acid by Escherichia coli. J. Bacteriol. 1999, 181, 47–54. [Google Scholar] [PubMed]
- Lewis, A.L.; Lewis, W.G. Host sialoglycans and bacterial sialidases: A mucosal perspective. Cell. Microbiol. 2012, 14, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sayeed, S.; Robertson, S.; Chen, J.; McClane, B.A. Sialidases affect the host cell adherence and epsilon toxin-induced cytotoxicity of Clostridium perfringens type D strain CN3718. PLoS Pathog. 2011, 7, e1002429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarezza, M.; Lyras, D.; Pidot, S.J.; Flores-Diaz, M.; Awad, M.M.; Kennedy, C.L.; Cordner, L.M.; Phumoonna, T.; Poon, R.; Hughes, M.L.; et al. The nani and nanj sialidases of Clostridium perfringens are not essential for virulence. Infect. Immun. 2009, 77, 4421–4428. [Google Scholar] [CrossRef] [PubMed]
- Manco, S.; Hernon, F.; Yesilkaya, H.; Paton, J.C.; Andrew, P.W.; Kadioglu, A. Pneumococcal neuraminidases A and B both have essential roles during infection of the respiratory tract and sepsis. Infect. Immun. 2006, 74, 4014–4020. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.H.; Blue, L.E.; James, M.A.; DeMaria, T.F. Evaluation of the virulence of a Streptococcus pneumoniae neuraminidase-deficient mutant in nasopharyngeal colonization and development of otitis media in the chinchilla model. Infect. Immun. 2000, 68, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.; Soong, G.; Planet, P.; Brower, J.; Ratner, A.J.; Prince, A. The nana neuraminidase of Streptococcus pneumoniae is involved in biofilm formation. Infect. Immun. 2009, 77, 3722–3730. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, S.; Tamai, E.; Matsushita, O.; Minami, J.; Okabe, A.; Miyata, S. Changes in ganglioside content affect the binding of Clostridium perfringens epsilon-toxin to detergent-resistant membranes of madin-darby canine kidney cells. Microbiol. Immunol. 2005, 49, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Galen, J.E.; Ketley, J.M.; Fasano, A.; Richardson, S.H.; Wasserman, S.S.; Kaper, J.B. Role of Vibrio cholerae neuraminidase in the function of cholera toxin. Infect. Immun. 1992, 60, 406–415. [Google Scholar] [PubMed]
- Vilei, E.M.; Johansson, A.; Schlatter, Y.; Redhead, K.; Frey, J. Genetic and functional characterization of the NanA sialidase from Clostridium chauvoei. Vet. Res. 2011, 42, 2. [Google Scholar] [CrossRef] [PubMed]
- Flores-Diaz, M.; Alape-Giron, A.; Clark, G.; Catimel, B.; Hirabayashi, Y.; Nice, E.; Gutierrez, J.M.; Titball, R.; Thelestam, M. A cellular deficiency of gangliosides causes hypersensitivity to Clostridium perfringens phospholipase c. J. Biol. Chem. 2005, 280, 26680–26689. [Google Scholar] [CrossRef] [PubMed]
- Chong, E.; Winikoff, B.; Charles, D.; Agnew, K.; Prentice, J.L.; Limbago, B.M.; Platais, I.; Louie, K.; Jones, H.E.; Shannon, C.; et al. Vaginal and rectal Clostridium sordellii and Clostridium perfringens presence among women in the United States. Obstet. Gynecol. 2016, 127, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Couchman, E.C.; Browne, H.P.; Dunn, M.; Lawley, T.D.; Songer, J.G.; Hall, V.; Petrovska, L.; Vidor, C.; Awad, M.; Lyras, D.; et al. Clostridium sordellii genome analysis reveals plasmid localized toxin genes encoded within pathogenicity loci. BMC Genom. 2015, 16, 392. [Google Scholar] [CrossRef] [PubMed]
- Palombo, E.A.; Yusoff, K.; Stanisich, V.A.; Krishnapillai, V.; Willetts, N.S. Cloning and genetic analysis of tra cistrons of the Tra 2/Tra 3 region of plasmid RP1. Plasmid 1989, 22, 59–69. [Google Scholar] [CrossRef]
- Miller, J. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Boyer, H.W.; Roulland-Dussoix, D. A complementation analysis of the restriction and modification of DNA in Escherichia coli. J. Mol. Biol. 1969, 41, 459–472. [Google Scholar] [CrossRef]
- Hall, I.C.; Scott, J.P. Bacillus sordellii, a cause of malignant edema in man. J. Infect. Dis. 1927, 41, 329–335. [Google Scholar] [CrossRef]
- Fagan, R.P.; Fairweather, N.F. Clostridium difficile has two parallel and essential sec secretion systems. J. Biol. Chem. 2011, 286, 27483–27493. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.K.; Keyburn, A.L.; Carter, G.P.; Lanckriet, A.L.; Van Immerseel, F.; Moore, R.J.; Rood, J.I. The virsr two-component signal transduction system regulates netb toxin production in Clostridium perfringens. Infect. Immun. 2010, 78, 3064–3072. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.R.; Johnson, S.; Gerding, D.N. Clostridium difficile infection caused by the epidemic BI/NAP1/027 strain. Gastroenterology 2009, 136, 1913–1924. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. Signalp 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Nancy, Y.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. Psortb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar]
- Unger-Torroledo, L.; Straub, R.; Lehmann, A.D.; Graber, F.; Stahl, C.; Frey, J.; Gerber, V.; Hoppeler, H.; Baum, O. Lethal toxin of Clostridium sordellii is associated with fatal equine atypical myopathy. Vet. Microbiol. 2010, 144, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Rimoldi, G.; Uzal, F.; Chin, R.P.; Palombo, E.A.; Awad, M.; Lyras, D.; Shivaprasad, H.L. Necrotic enteritis in chickens associated with Clostridium sordellii. Avian Dis. 2015, 59, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Crespo, R.; Franca, M.; Shivaprasad, H.L. Ulcerative enteritis-like disease associated with Clostridium sordellii in quail. Avian Dis. 2013, 57, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Aspholm, M.; Olfat, F.O.; Norden, J.; Sonden, B.; Lundberg, C.; Sjostrom, R.; Altraja, S.; Odenbreit, S.; Haas, R.; Wadstrom, T.; et al. Saba is the H. pylori hemagglutinin and is polymorphic in binding to sialylated glycans. PLoS Pathog. 2006, 2, e110. [Google Scholar] [CrossRef] [PubMed]
- Dalia, A.B.; Standish, A.J.; Weiser, J.N. Three surface exoglycosidases from Streptococcus pneumoniae, NanA, BgaA, and StrH, promote resistance to opsonophagocytic killing by human neutrophils. Infect. Immun. 2010, 78, 2108–2116. [Google Scholar] [CrossRef] [PubMed]
- Nardy, A.F.; Freire-de-Lima, C.G.; Perez, A.R.; Morrot, A. Role of Trypanosoma cruzi trans-sialidase on the escape from host immune surveillance. Front. Microbiol. 2016, 7, 348. [Google Scholar] [CrossRef] [PubMed]
- Vimr, E.; Lichtensteiger, C. To sialylate, or not to sialylate: That is the question. Trends Microbiol. 2002, 10, 254–257. [Google Scholar] [CrossRef]
- Crost, E.H.; Tailford, L.E.; Le Gall, G.; Fons, M.; Henrissat, B.; Juge, N. Utilisation of mucin glycans by the human gut symbiont ruminococcus gnavus is strain-dependent. PLoS ONE 2013, 8, e76341. [Google Scholar] [CrossRef] [PubMed]
- Cribby, S.; Taylor, M.; Reid, G. Vaginal microbiota and the use of probiotics. Interdiscip. Perspect. Infect. Dis. 2008, 2008, 256490. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; Hedges, S.R.; Mordechai, E.; Adelson, M.E.; Trama, J.P.; Gygax, S.E.; Kaunitz, A.M.; Hilbert, D.W. Cervical and vaginal flora specimens are highly concordant with respect to bacterial vaginosis-associated organisms and commensal lactobacillus species in women of reproductive age. J. Clin. Microbiol. 2014, 52, 3078–3081. [Google Scholar] [CrossRef] [PubMed]
- Borgdorff, H.; Tsivtsivadze, E.; Verhelst, R.; Marzorati, M.; Jurriaans, S.; Ndayisaba, G.F.; Schuren, F.H.; van de Wijgert, J.H. Lactobacillus-dominated cervicovaginal microbiota associated with reduced HIV/STI prevalence and genital HIV viral load in african women. ISME J. 2014, 8, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Juge, N.; Tailford, L.; Owen, C.D. Sialidases from gut bacteria: A mini-review. Biochem. Soc. Trans. 2016, 44, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Perez-Lopez, A.; Behnsen, J.; Nuccio, S.P.; Raffatellu, M. Mucosal immunity to pathogenic intestinal bacteria. Nat. Rev. Immunol. 2016, 16, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.M.; Ferreyra, J.A.; Higginbottom, S.K.; Lynch, J.B.; Kashyap, P.C.; Gopinath, S.; Naidu, N.; Choudhury, B.; Weimer, B.C.; Monack, D.M.; et al. Microbiota-liberated host sugars facilitate post-antibiotic expansion of enteric pathogens. Nature 2013, 502, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, R.; Hicks, S.J.; Soothill, P.W.; Millar, M.R.; Corfield, A.P. Mucinases and sialidases: Their role in the pathogenesis of sexually transmitted infections in the female genital tract. Sex. Transm. Infect. 2001, 77, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Meites, E.; Zane, S.; Gould, C. Fatal Clostridium sordellii infections after medical abortions. N. Engl. J. Med. 2010, 363, 1382–1383. [Google Scholar] [CrossRef] [PubMed]
- Soong, G.; Muir, A.; Gomez, M.I.; Waks, J.; Reddy, B.; Planet, P.; Singh, P.K.; Kaneko, Y.; Wolfgang, M.C.; Hsiao, Y.S.; et al. Bacterial neuraminidase facilitates mucosal infection by participating in biofilm production. J. Clin. Investig. 2006, 116, 2297–2305. [Google Scholar] [CrossRef] [PubMed]
- Riedewald, S.; Kreutzmann, I.M.; Heinze, T.; Saling, E. Vaginal and cervical pH in normal pregnancy and pregnancy complicated by preterm labor. J. Perinat. Med. 1990, 18, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Aronoff, D.M.; Hao, Y.; Chung, J.; Coleman, N.; Lewis, C.; Peres, C.M.; Serezani, C.H.; Chen, G.H.; Flamand, N.; Brock, T.G.; et al. Misoprostol impairs female reproductive tract innate immunity against Clostridium sordellii. J. Immunol. 2008, 180, 8222–8230. [Google Scholar] [CrossRef] [PubMed]
- Rorbye, C.; Petersen, I.S.; Nilas, L. Postpartum Clostridium sordellii infection associated with fatal toxic shock syndrome. Acta Obstet. Gynecol. Scand. 2000, 79, 1134–1135. [Google Scholar] [PubMed]
- Qa’Dan, M.; Spyres, L.M.; Ballard, J.D. pH-enhanced cytopathic effects of Clostridium sordellii lethal toxin. Infect. Immun. 2001, 69, 5487–5493. [Google Scholar] [CrossRef] [PubMed]
- Voth, D.E.; Qa’Dan, M.; Hamm, E.E.; Pelfrey, J.M.; Ballard, J.D. Clostridium sordellii lethal toxin is maintained in a multimeric protein complex. Infect. Immun. 2004, 72, 3366–3372. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Characteristics | Reference |
|---|---|---|
| Strains | ||
| E. coli | ||
| DH5α | F− ϕ80dlacZΔM15 Δ(lacZYA argF)U169 endA1 recA1 hsdR17 (rK− mK−) deoR thi-1 supE44 gyrA96 relA1 | Life Technologies |
| One-Shot TOP10 | F− mcrA (mrr-hsdRMS-mcrBC) 80lacZ M15 lacX74 nupG recA1 araD139 (araleu) 7697 galU galK rpsL(Strr) endA1 λ _ | Invitrogen |
| HB101 | thi-1 hsdS20(rB− mB−) supE44 recAB ara-14 leuB5 proA2 lacY1 galK rpsL20 (Smr) xyl-5 mtl-1 | [29] |
| C. sordellii | ||
| ATCC 9714 | Type strain; TcsL+ TcsH− Nan+ SDL− PLC+ | [30] |
| DLL5034 | ATCC 9714 nanSΩtargetron; Emr | This study |
| DLL5101 | DLL5034(pDLL197) | This study |
| DLL5002 | ATCC 9714 tcsLΩtargetron; Emr | [8] |
| Plasmids | ||
| pVS520 | Tra+ Mob+ RP4 derivative; Tcr | [27] |
| pRPF185 | E. coli–C. difficile shuttle vector, Ptet::gusA cat CD6ori RP4oriT-traJ pMB1ori | [31] |
| pJIR3566 | Clostridial mobilizable targetron vector; with pCB102 replication region, lacZ, oriT; Cmr | [32] |
| pJIR3566 | Group II intron of pJIR3566 retargeted to the 304/305s site of the nanS gene | This study |
| pDLL197 | pRPF185 (SacI/BamHI) DLP202/DLP233 nanS PCR product (SacI/BamHI; 1654 bp); Cmr | This study |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awad, M.M.; Singleton, J.; Lyras, D. The Sialidase NanS Enhances Non-TcsL Mediated Cytotoxicity of Clostridium sordellii. Toxins 2016, 8, 189. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8060189
Awad MM, Singleton J, Lyras D. The Sialidase NanS Enhances Non-TcsL Mediated Cytotoxicity of Clostridium sordellii. Toxins. 2016; 8(6):189. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8060189
Chicago/Turabian StyleAwad, Milena M., Julie Singleton, and Dena Lyras. 2016. "The Sialidase NanS Enhances Non-TcsL Mediated Cytotoxicity of Clostridium sordellii" Toxins 8, no. 6: 189. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8060189