
Susceptibility of Broiler Chickens to Coccidiosis When Fed Subclinical Doses of Deoxynivalenol and Fumonisins—Special Emphasis on the Immunological Response and the Mycotoxin Interaction

 , , and
, , and
Abstract
:
1. Introduction
2. Results
2.1. Experiment 1: Feeding Trial with Diets Contaminated with DON and FB, Alone or in Combination
2.1.1. Performance, Clinical Signs, Lesions, and Oocyst Identification
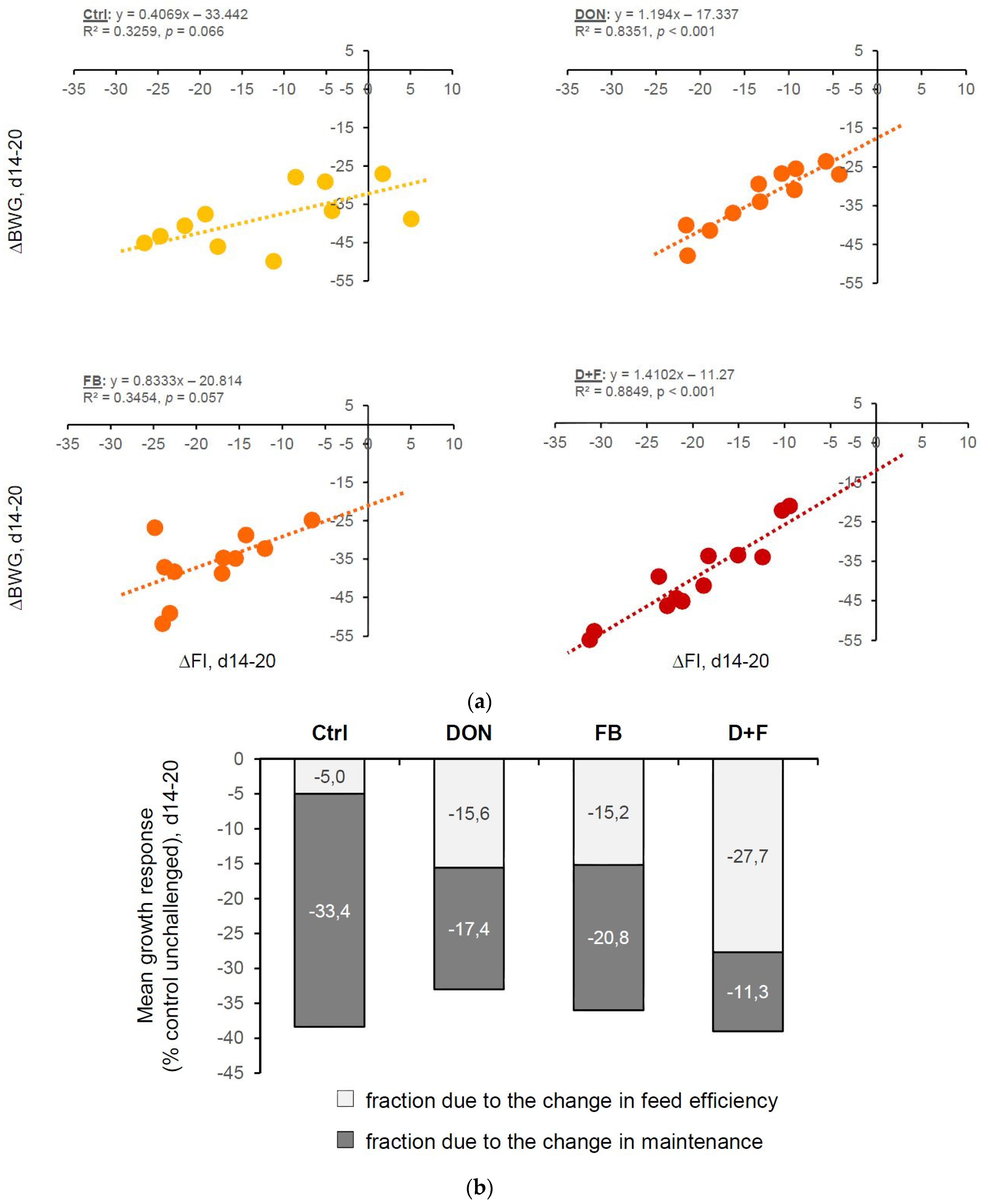
Performance
Clinical Signs
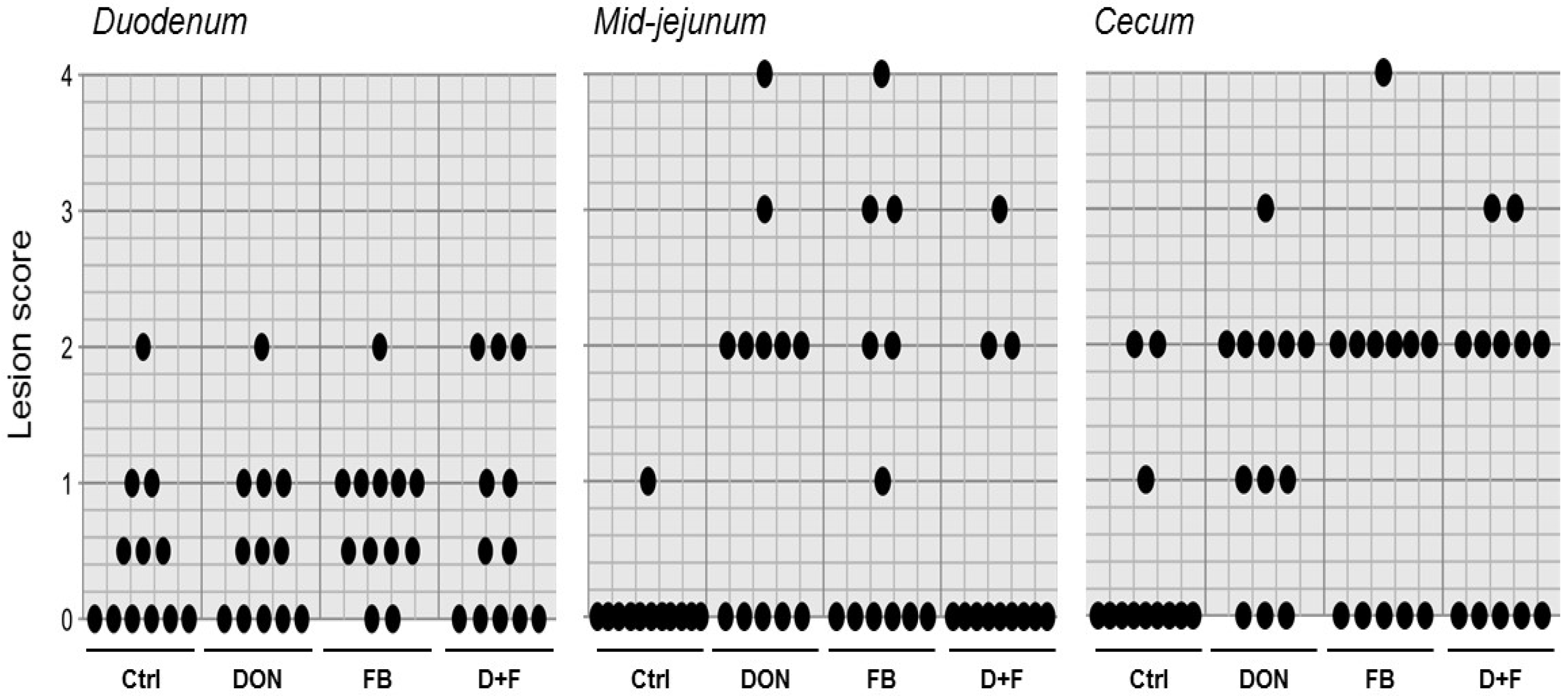
Gross Lesions
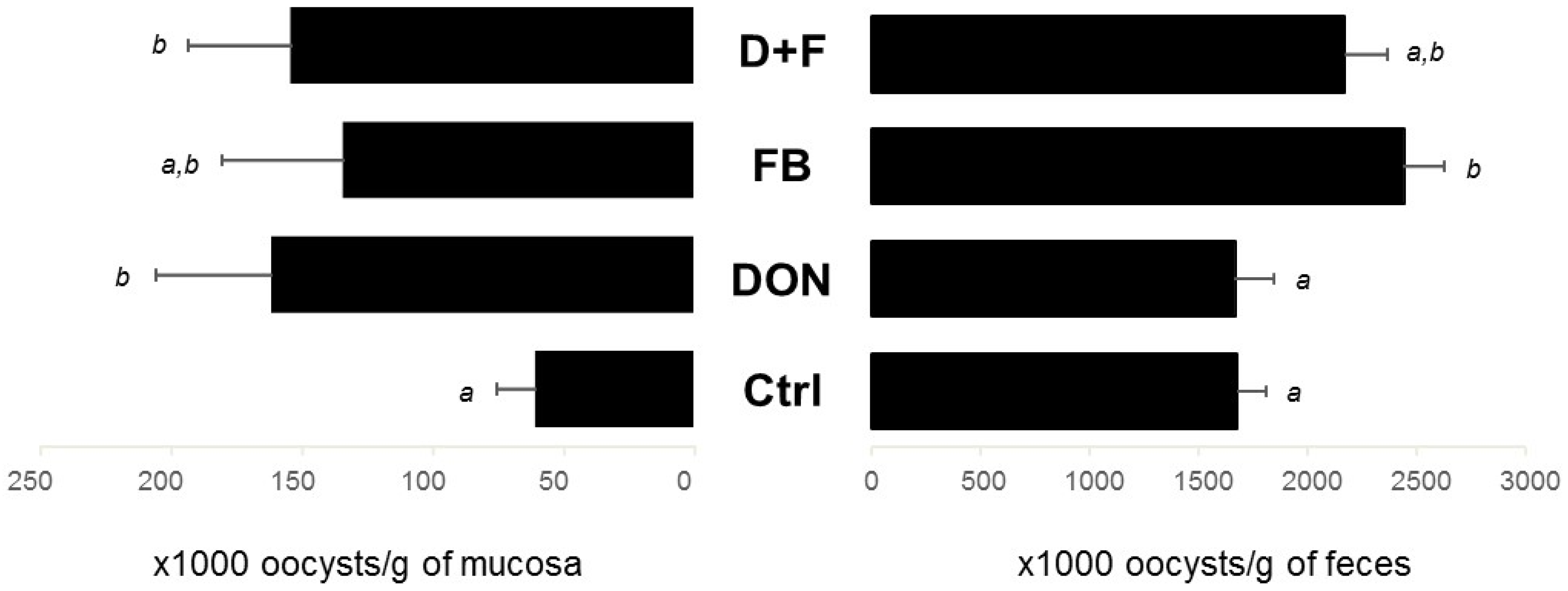
Oocyst Identification
2.1.2. Gut Integrity
Villi Height
Apparent digestibility
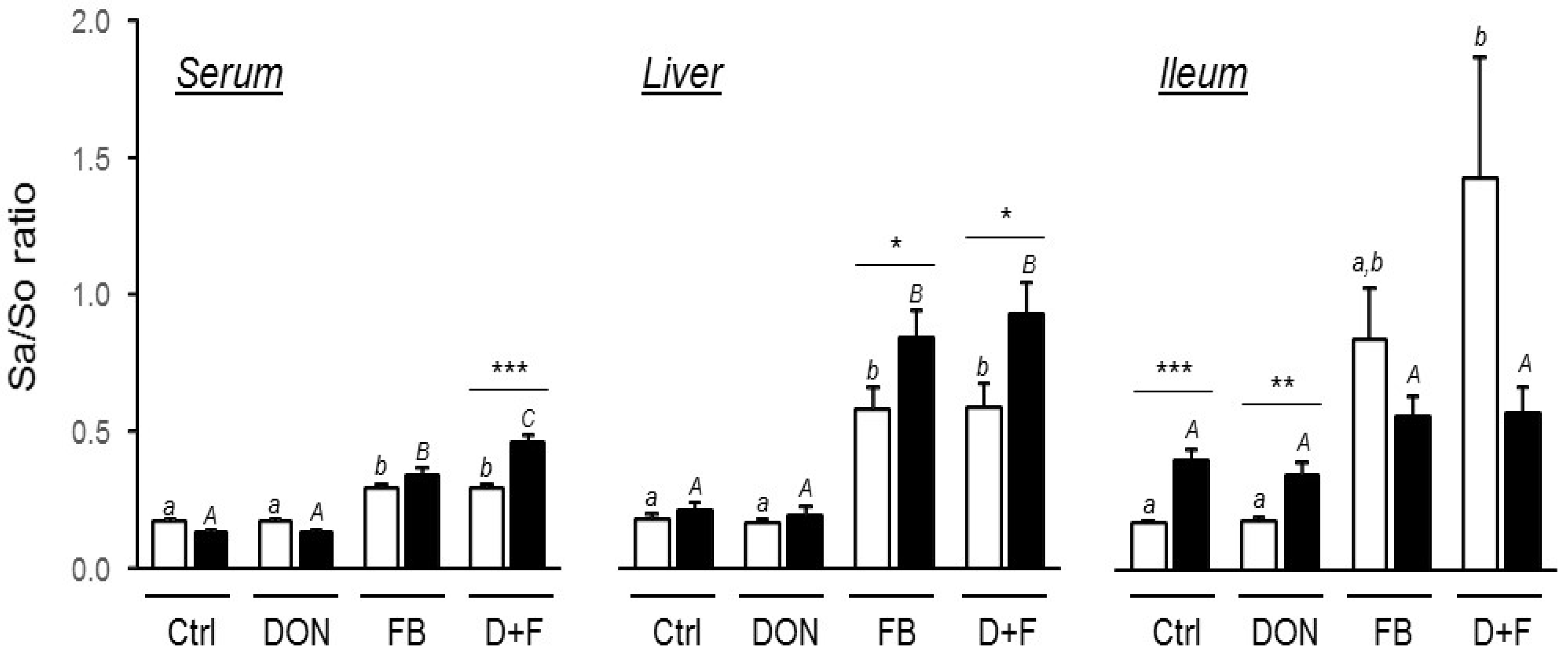
Fumonisin Exposure
Gene Expression
2.1.3. Gut Inflammation
Gene Expression
2.1.4. Interaction between Mycotoxins
Factorial ANOVA
2.2. Experiment 2: Feeding Trial with Diets Contaminated with DON
2.2.1. Gut Inflammation
Lymphocytes T Cells
Gene Expression
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experiment 1: Feeding Trial with Diets Contaminated with DON and FB, Alone or in Combination
5.1.1. Experimental Birds, Housing, Diet Formulation and Sampling
Diet Formulation
Birds and Housing
Challenge
Sampling
5.1.2. Experimental Parameter Measures
Digestibility and Retention Analysis
Lesion Score and Oocyst Counting
Sphingolipid Analysis
Gene Expression Analysis
Histological Analysis
5.1.3. Statistics
5.2. Experiment 2: Feeding Trial with Diets Contaminated with DON
Experimental Birds, Diet Formulation, Sampling and Experimental Parameter Measures
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DON | Deoxynivalenol |
| FB | Fumonisins |
| GIT | Gastrointestinal Tract |
| Sa | Sphinganine |
| So | Sphingosine |
References
- Oswald, I.P.; Marin, D.E.; Bouhet, S.; Pinton, P.; Taranu, I.; Accensi, F. Immunotoxicological risk of mycotoxins for domestic animals. Food Addit. Contam. 2005, 22, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on europe and asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Girgis, G.N.; Sharif, S.; Barta, J.R.; Boermans, H.J.; Smith, T.K. Immunomodulatory effects of feed-borne fusarium mycotoxins in chickens infected with coccidia. Exp. Biol. Med. 2008, 233, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Kana, J.R.; Teguia, A.; Tchoumboue, J. Effect of dietary plant charcoal from canarium schweinfurthii engl. And maize cob on aflatoxin b1 toxicosis in broiler chickens. Adv. Anim. Biosci. 2010, 1, 462–463. [Google Scholar] [CrossRef]
- Awad, W.A.; Vahjen, W.; Aschenbach, J.R.; Zentek, J. A diet naturally contaminated with the fusarium mycotoxin deoxynivalenol (don) downregulates gene expression of glucose transporters in the intestine of broiler chickens. Livest. Sci. 2011, 140, 72–79. [Google Scholar] [CrossRef]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Haesebrouck, F.; Timbermont, L.; Verlinden, M.; Janssens, G.P.J.; Eeckhaut, V.; Eeckhout, M.; et al. The mycotoxin deoxynivalenol predisposes for the development of clostridium perfringens-induced necrotic enteritis in broiler chickens. PLoS ONE 2014, 9, e108775. [Google Scholar]
- Grenier, B.; Schwartz-Zimmermann, H.; Caha, S.; Moll, W.; Schatzmayr, G.; Applegate, T. Dose-dependent effects on sphingoid bases and cytokines in chickens fed diets prepared with fusarium verticillioides culture material containing fumonisins. Toxins 2015, 7, 1253–1272. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.; De Baere, S.; Mountzouris, K.; Su, S.; Wong, A.; De Meulenaer, B.; et al. Mycotoxins deoxynivalenol and fumonisins alter the extrinsic component of intestinal barrier in broiler chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef] [PubMed]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin b1 in affecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins 2011, 3, 566–590. [Google Scholar] [CrossRef] [PubMed]
- Cheema, M.; Qureshi, M.; Havenstein, G. A comparison of the immune response of a 2001 commercial broiler with a 1957 randombred broiler strain when fed representative 1957 and 2001 broiler diets. Poult. Sci. 2003, 82, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.D. Milestones in avian coccidiosis research: A review. Poult. Sci. 2014, 93, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Dalloul, R.A.; Lillehoj, H.S. Poultry coccidiosis: Recent advancements in control measures and vaccine development. Expert Rev. Vaccines 2006, 5, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Q.; Kanu, S.; Xiao, S.M.; Xiang, F.Y. Responses of chickens vaccinated with a live attenuated multi-valent ionophore-tolerant eimeria vaccine. Vet. Parasitol. 2005, 129, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Farnell, M.B.; Donoghue, A.M.; de los Santos, F.S.; Blore, P.J.; Hargis, B.M.; Tellez, G.; Donoghue, D.J. Upregulation of oxidative burst and degranulation in chicken heterophils stimulated with probiotic bacteria. Poult. Sci. 2006, 85, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- Girgis, G.N.; Barta, J.R.; Brash, M.; Smith, T.K. Morphologic changes in the intestine of broiler breeder pullets fed diets naturally contaminated with fusarium mycotoxins with or without coccidial challenge. Avian Dis. 2010, 54, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Girgis, G.N.; Barta, J.R.; Girish, C.K.; Karrow, N.A.; Boermans, H.J.; Smith, T.K. Effects of feed-borne fusarium mycotoxins and an organic mycotoxin adsorbent on immune cell dynamics in the jejunum of chickens infected with eimeria maxima. Vet. Immunol. Immunopathol. 2010, 138, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Chou, T.-C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Kolf-Clauw, M.; Gauthier, T.; Abrami, R.; Abiola, F.A.; Oswald, I.P.; Puel, O. New insights into mycotoxin mixtures: The toxicity of low doses of type b trichothecenes on intestinal epithelial cells is synergistic. Toxicol. Appl. Pharmacol. 2013, 272, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Adedokun, S.A.; Ajuwon, K.M.; Romero, L.F.; Adeola, O. Ileal endogenous amino acid losses: Response of broiler chickens to fiber and mild coccidial vaccine challenge. Poult. Sci. 2012, 91, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Selvaraj, R.K. Regulatory t cell properties of chicken CD4+CD25+ cells. J. Immunol. 2011, 186, 1997–2002. [Google Scholar] [CrossRef] [PubMed]
- Pastorelli, H.; van Milgen, J.; Lovatto, P.; Montagne, L. Meta-analysis of feed intake and growth responses of growing pigs after a sanitary challenge. Animal 2012, 6, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Min, W.; Dalloul, R.A. Recent progress on the cytokine regulation of intestinal immune responses to eimeria. Poult. Sci. 2004, 83, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, G.R.; Ledoux, D.R.; Naehrer, K.; Berthiller, F.; Applegate, T.J.; Grenier, B.; Phillips, T.D.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.H.; Wyatt, R.D.; Fletchert, O.J. The toxicity of purified fumonisin b1 in broiler chicks. Poult. Sci. 2000, 79, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, G.S.; Pettersson, H. Toxicological evaluation of trichothecenes in animal feed. Anim. Feed Sci. Technol. 2004, 114, 205–239. [Google Scholar] [CrossRef]
- Broomhead, J.N.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Chronic effects of fumonisin b1 in broilers and turkeys fed dietary treatments to market age. Poult. Sci. 2002, 81, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Yunus, A.W.; Ghareeb, K.; Twaruzek, M.; Grajewski, J.; Böhm, J. Deoxynivalenol as a contaminant of broiler feed: Effects on bird performance and response to common vaccines. Poult. Sci. 2012, 91, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Stringfellow, K.; Caldwell, D.; Lee, J.; Mohnl, M.; Beltran, R.; Schatzmayr, G.; Fitz-Coy, S.; Broussard, C.; Farnell, M. Evaluation of probiotic administration on the immune response of coccidiosis-vaccinated broilers. Poult. Sci. 2011, 90, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Paris, N.E.; Wong, E.A. Expression of digestive enzymes and nutrient transporters in the intestine of eimeria maxima-infected chickens. Poult. Sci. 2013, 92, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Devreese, M.; Van Immerseel, F.; De Baere, S.; Hessenberger, S.; Martel, A.; Croubels, S. Chronic exposure to deoxynivalenol has no influence on the oral bioavailability of fumonisin b1 in broiler chickens. Toxins 2015, 7, 560–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, C.H.; Lillehoj, H.S.; Lillehoj, E.P. Intestinal immune responses to coccidiosis. Dev. Comp. Immunol. 2000, 24, 303–324. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Lillehoj, E.P.; Lee, S.H. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following eimeria maxima infection of chickens. Vet. Immunol. Immunopathol. 2006, 114, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Applegate, T.J.; Lossie, A.C. Cloning, annotation and developmental expression of the chicken intestinal muc2 gene. PLoS ONE 2013, 8, e53781. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Selvaraj, R.K. CD4+CD25+ regulatory t cell ontogeny and preferential migration to the cecal tonsils in chickens. PLoS ONE 2012, 7, e33970. [Google Scholar] [CrossRef] [PubMed]
- Kubena, L.; Edrington, T.; Harvey, R.; Buckley, S.; Phillips, T.; Rottinghaus, G.; Casper, H. Individual and combined effects of fumonisin b1 present in fusarium moniliforme culture material and t-2 toxin or deoxynivalenol in broiler chicks. Poult. Sci. 1997, 76, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Plattner, R.D.; Shackelford, D.D.; Leslie, J.F.; Nelson, P.E. Heritability of fumonisin b1 production in gibberella fujikuroi mating population a. Appl. Environ. Microbiol. 1992, 58, 2799–2805. [Google Scholar] [PubMed]
- Altpeter, F.; Posselt, U.K. Production of high quantities of 3-acetyldeoxynivalenol and deoxynivalenol. Appl. Microbiol. Biotechnol. 1994, 41, 384–387. [Google Scholar] [CrossRef]
- Hodgson, J.N. Coccidiosis: Oocyst counting technique for coccidiostat evaluation. Exp. Parasitol. 1970, 28, 99–102. [Google Scholar] [CrossRef]
- Hill, F.W.; Anderson, D.L. Comparison of metabolizable energy and productive energy determinations with growing chicks. J. Nutr. 1958, 64, 587–603. [Google Scholar] [PubMed]
- NRC. Nutrient Requirements of Poultry, 9th ed.; Natl. Acad. Press: Washington, DC, USA, 1994. [Google Scholar]
- Johnson, J.; Reid, W.M. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef]
- Conway, D.P.; McKenzie, M.E. Examination of lesions and lesion scoring. In Poultry Coccidiosis, 3rd ed.; Blackwell Publishing Professional: Ames, IA, USA, 2008; pp. 14–40. [Google Scholar]
- Peirson, S.N.; Butler, J.N.; Foster, R.G. Experimental validation of novel and conventional approaches to quantitative real-time pcr data analysis. Nucleic Acids Res. 2003, 31, e73. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BWG per Bird (g) | Unchallenged | Challenged | ||||||
|---|---|---|---|---|---|---|---|---|
| Ctrl | DON | FB | DON + FB | Ctrl | DON | FB | DON + FB | |
| 7 birds/pen | ||||||||
| d0–d14 | 334 ± 7 a,b | 357 ± 10 b | 343 ± 11 b | 303 ± 12 a | 334 ± 13 | 354 ± 14 | 340 ± 8 | 332 ± 8 |
| d14–d20 | 300 ± 20 | 347 ± 12 | 325 ± 6 | 312 ± 10 | 225 ± 15 | 207 ± 6 | 225 ± 8 | 202 ± 15 |
| 4 birds/pen | ||||||||
| d0–d14 | 337± 12 | 342 ± 13 | 352 ± 13 | 330 ± 20 | 301 ± 12 a | 379 ± 12 b | 337 ± 11 a,b | 333 ± 11 a |
| d14–d20 | 339 ± 18 | 332 ± 17 | 331 ± 15 | 322 ± 27 | 195 ± 10 a,b | 231 ± 12 b | 188 ± 12 a | 200 ± 15 a,b |
| d20–d34 * | 1014 ± 31 | 997 ± 46 | 984 ± 69 | 962 ± 62 | 981 ± 9 | 1022 ± 31 | 986 ± 21 | 1021 ± 12 |
| Groups | Apparent DM Digestibility (%) | Apparent N Digestibility (%) | Villus Height (μm/villus) |
|---|---|---|---|
| Unchallenged birds | |||
| Control | 63.7 ± 0.9 | 79.6 ± 0.9 | 862 ± 31 |
| DON | 67.1 ± 1.4 | 81.8 ± 0.7 | 892 ± 42 |
| FB | 63.5 ± 0.9 | 80.2 ± 0.5 | 829 ± 24 |
| DON + FB | 62.6 ± 1.6 | 80.5 ± 0.8 | 802 ± 27 |
| Challenged birds | |||
| Control | 32.7 ± 4.8 | 56.6 ± 2.1 a | ND |
| DON | 36.6 ± 5.4 | 56.3 ± 3.3 a,b | ND |
| FB | 42.2 ± 4.7 | 60.9 ± 2.5 a | ND |
| DON + FB | 32.1 ± 4.4 | 48.8 ± 3.2 b | ND |
| Markers Assessed | No Effect of DON, FB or DON + FB —No Interaction Determined * | Type of Interaction between DON and FB * | ||
|---|---|---|---|---|
| Antagonistic | Additive | Synergistic | ||
| markers of pathogenicity | - | Lesions jejunum (0.036) | Oocyst excreta (0.47) | - |
| - | - | Oocyst mucosa (0.21) | - | |
| - | - | Lesions duod. (0.17) | - | |
| - | - | Lesions ceca (0.47) | - | |
| markers of gut inflammation | IL-17 | IL-6 (0.036) | IL-8 (0.14) | - |
| IL-21 | IL-1β (0.095, tendency) | IL-10 (0.19) | - | |
| IFN-γ | SOCS1 (0.47) | - | ||
| markers of gut integrity | MUC2 | - | SGLT1 (0.71) | Digestibility N (0.047) |
| Digestibility DM | - | - | - | |
| marker of exposure/effect | Sa/So ileum | - | Sa/So liver (0.47) | Sa/So serum (0.007, potentiation) |
| number of endpoints | 6 | 3 | 9 | 2 |
| relative to total endpoints | 30% | 15% | 45% | 10% |
| Parameters | Unchallenged | Challenged | ||
|---|---|---|---|---|
| Control | DON | Control | DON | |
| Lymphocyte (%) | ||||
| CD4+CD25+ | 12 ± 3 a | 12 ± 4 a | 27 ± 3 b | 45 ± 7 c |
| CD4+ | 45 ± 3 a | 42 ± 2 a | 52 ± 2 a,b | 66 ± 4 b |
| CD8+ | 36 ± 2 a | 35 ± 5 a,b | 25 ± 3 b | 40 ± 5 a |
| Gene expression | ||||
| IL-1β | 1.00 ± 0.14 a | 1.01 ± 0.14 a | 1.79 ± 0.53 a,b | 3.19 ± 0.68 b |
| IL-6 | 1.00 ± 0.09 a | 0.59 ± 0.08 b | 4.73 ± 1.54 c | 4.57 ± 0.84 c |
| IL-17 | 1.00 ± 0.10 a | 0.85 ± 0.10 a | 0.56 ± 0.06 b | 0.50 ± 0.07 b |
| IFN-γ | 1.00 ± 0.11 a | 1.02 ± 0.16 a | 4.41 ± 0.65 b | 8.49 ± 1.41 c |
| IL-10 | 1.00 ± 0.16 a | 1.08 ± 0.20 a | 1.97 ± 0.36 b | 2.27 ± 0.22 b |
| SOCS1 | 1.00 ± 0.07 | 0.98 ± 0.15 | 1.32 ± 0.08 | 1.38 ± 0.19 |
| Item | Starter Diet (day 0–20) | Grower Diet (day 21–34) |
|---|---|---|
| Ingredient (% of diet) | ||
| Corn | 53.98 | 52.10 |
| Soybean meal (48% CP) | 38.05 | 39.09 |
| Soy oil | 3.52 | 5.00 |
| Sodium chloride | 0.48 | 0.46 |
| DL-Methionine | 0.25 | 0.24 |
| Threonine | 0.07 | - |
| L-Lysine, HCl | 0.10 | - |
| Limestone | 1.68 | 1.56 |
| Monocalcium phosphate | 1.52 | 1.20 |
| Vitamin and mineral premix 1 | 0.35 | 0.35 |
| Nutrient composition (calculated) | ||
| ME, kcal/kg | 3066 | 3151 |
| CP, % | 22.43 | 22.81 |
| Ca, % | 1.01 | 0.85 |
| Non-phytate phosphorus, % | 0.43 | 0.44 |
| Met, % | 0.59 | 0.59 |
| Thr, % | 0.92 | 0.89 |
| Lys, % | 1.34 | 1.29 |
| Analyzed composition | Deoxynivalenol (mg/kg) | Fumonisins 2 (mg/kg) |
| Control diet | - | 3.1 |
| DON diet | 1.6 | 2.9 |
| FB diet | - | 20.5 |
| DON + FB diet | 1.3 | 20.8 |
| Gene | Primer Sequence | Amplicon | Ensembl Access | Reference | |
|---|---|---|---|---|---|
| Size | Intron 1 | ||||
| Housekeeping genes | |||||
| GAPDH | F (300 nM) TCCTAGGATACACAGAGGACCA R (300 nM) CGGTTGCTATATCCAAACTCA | 151 bp | 2 (499) | ENSGALG00000014442 | [9] |
| RPL4 | F (300 nM) TTATGCCATCTGTTCTGCC R (300 nM) GCGATTCCTCATCTTACCCT | 235 bp | 2 (893) | ENSGALG00000007711 | [9] |
| Pro-inflammatory cytokines | |||||
| IL-1β | F (300 nM) GCATCAAGGGCTACAAGCTC R (300 nM) CAGGCGGTAGAAGATGAAGC | 131 bp | 1 (87) | ENSGALG00000000534 | [22] |
| IL-6 | F (300 nM) GAATGTTTTAGTTCGGGCACA R (300 nM) TTCCTAGAAGGAAATGAGAATGC | 130 bp | 0 | ENSGALG00000010915 | [9] |
| IL-8 | F (300 nM) GCGGCCCCCACTGCAAGAAT R (300 nM) TCACAGTGGTGCATCAGAATTGAGC | 146 bp | 2 (1210) | ENSGALG00000011670 | [9] |
| Treg signature | |||||
| IL-10 | F (300 nM) GCTGAGGGTGAAGTTTGAGG R (300 nM) AGACTGGCAGCCAAAGGTC | 121 bp | 2 (1127) | ENSGALG00000000892 | [9] |
| SOCS1 | F (300 nM) CAAGCGGATTTCAGTAGCATC R (300 nM) GGCTCAGACTTCAGCTTCTCA | 110 bp | no intron | ENSGALG00000007158 | [9] |
| Th17 & Th1 signature | |||||
| IL-17 | F (300 nM) TATCAGCAAACGCTCACTGG R (300 nM) AGTTCACGCACCTGGAATG | 110 bp | 1 (666) | ENSGALG00000016678 | [9] |
| IL-21 | F (300 nM) GCTTTCAAAGACAATTGACCATC R (300 nM) TACAGCTGTGAGCAGGCATC | 106 bp | 2 (3765) | ENSGALG00000011844 | [9] |
| IFN-γ | F (300 nM) AGCTGACGGTGGACCTATTATT R (300 nM) GGCTTTGCGCTGGATTC | 259 bp | 2 (998) | ENSGALG00000009903 | [37] |
| Gut integrity | |||||
| SGLT1 | F (300 nM) CATCTTCCGAGATGCTGTCA R (300 nM) AGGTATCCGCACATCACACA | 168 bp | 1 (979) | ENSGALG00000006728 | Present study |
| MUC2 | F (300 nM) CAGCACCAACTTCTCAGTTCC R (300 nM) TCTGCAGCCACACATTCTTT | 102 bp | 1 (1711) | ENSGALG00000006752 | Present study |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grenier, B.; Dohnal, I.; Shanmugasundaram, R.; Eicher, S.D.; Selvaraj, R.K.; Schatzmayr, G.; Applegate, T.J. Susceptibility of Broiler Chickens to Coccidiosis When Fed Subclinical Doses of Deoxynivalenol and Fumonisins—Special Emphasis on the Immunological Response and the Mycotoxin Interaction. Toxins 2016, 8, 231. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8080231
Grenier B, Dohnal I, Shanmugasundaram R, Eicher SD, Selvaraj RK, Schatzmayr G, Applegate TJ. Susceptibility of Broiler Chickens to Coccidiosis When Fed Subclinical Doses of Deoxynivalenol and Fumonisins—Special Emphasis on the Immunological Response and the Mycotoxin Interaction. Toxins. 2016; 8(8):231. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8080231
Chicago/Turabian StyleGrenier, Bertrand, Ilse Dohnal, Revathi Shanmugasundaram, Susan D. Eicher, Ramesh K. Selvaraj, Gerd Schatzmayr, and Todd J. Applegate. 2016. "Susceptibility of Broiler Chickens to Coccidiosis When Fed Subclinical Doses of Deoxynivalenol and Fumonisins—Special Emphasis on the Immunological Response and the Mycotoxin Interaction" Toxins 8, no. 8: 231. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins8080231