
Temperature Influences the Production and Transport of Saxitoxin and the Expression of sxt Genes in the Cyanobacterium Aphanizomenon gracile



Abstract
:1. Introduction
2. Results
2.1. Ecophysiology of Aphanizomenon Gracile
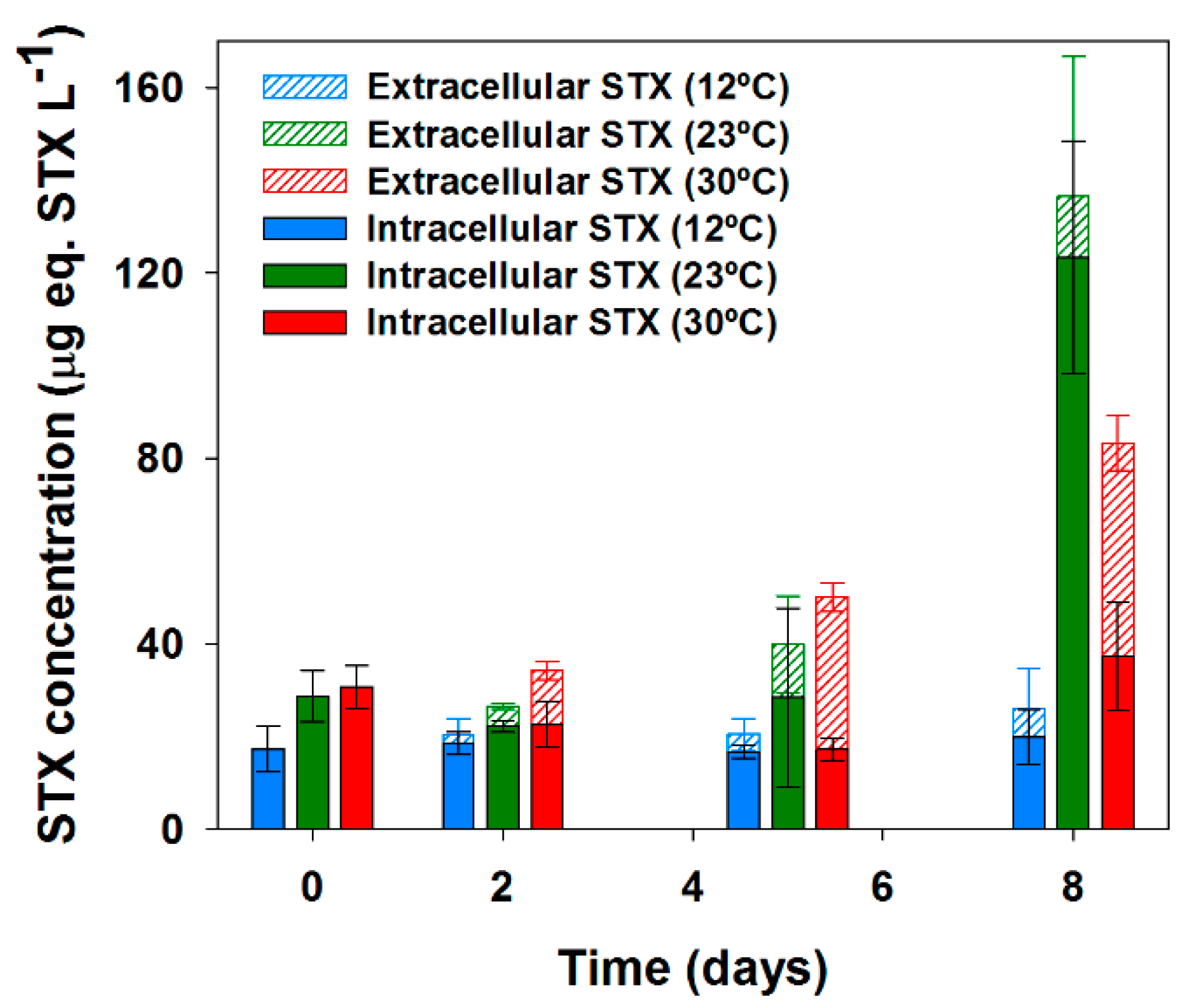
2.2. Effects of Temperature on Saxitoxin Production and Release
2.3. Effects of Temperature on the Expression of sxt Genes
2.3.1. Dynamics of STX-Biosynthesis Gene sxtA
2.3.2. Dynamics of STX-Transporter Genes sxtM and sxtPer
2.4. Effects of Nitrogen Depletion at 23 °C
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Culture Conditions
5.2. Growth Dynamics
5.3. Alterations in Membrane Integrity and Membrane Potential by Flow Cytometry
5.4. Analysis of the Expression of Genes sxtA, sxtM, and sxtPer by qPCR
5.4.1. Total RNA Extraction and cDNA Synthesis
5.4.2. Real Time qPCR
5.5. Quantification of Intracellular and Extracellular Saxitoxins
5.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef] [PubMed]
- Negri, A.P.; Jones, G.J. Bioaccumulation of paralytic shellfish poisoning (PSP) toxins from the cyanobacterium Anabaena circinalis by the freshwater mussel Alathyria condola. Toxicon 1995, 33, 667–678. [Google Scholar] [CrossRef]
- Berry, J.P.; Lind, O. First evidence of “paralytic shellfish toxins” and cylindrospermopsin in a mexican freshwater system, Lago Catemaco, and apparent bioaccumulation of the toxins in “tegogolo” snails (Pomacea patula catemacensis). Toxicon 2010, 55, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Dias, E.; Franca, S.; Pereira, E.; Carolino, M.; Vasconcelos, V. Accumulation and depuration of cyanobacterial paralytic shellfish toxins by the freshwater mussel Anodonta cygnea. Aquat. Toxicol. 2004, 68, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Kleinteich, J.; Wood, S.A.; Puddick, J.; Schleheck, D.; Küpper, F.C.; Dietrich, D. Potent toxins in Arctic environments–presence of saxitoxins and an unusual microcystin variant in Arctic freshwater ecosystems. Chem.-Biol. Interact. 2013, 206, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.M.J.; Wood, S.A.; van Ginkel, R.; Broady, P.A.; Gaw, S. First report of saxitoxin production by a species of the freshwater benthic cyanobacterium, Scytonema Agardh. Toxicon 2011, 57, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Cirés, S.; Ballot, A. A review of the phylogeny, ecology and toxin production of bloom-forming Aphanizomenon spp. and related species within the Nostocales (cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Cerasino, L.; Hostyeva, V.; Cirés, S. Variability in the sxt gene clusters of PSP toxin producing Aphanizomenon gracile strains from Norway, Spain, Germany and North America. PLoS ONE 2016, 11, e0167552. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.A.; Mihali, T.K.; Neilan, B.A. Extraordinary conservation, gene loss, and positive selection in the evolution of an ancient neurotoxin. Mol. Biol. Evol. 2011, 28, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Soto-Liebe, K.; López-Cortés, X.A.; Fuentes-Valdés, J.J.; Stucken, K.; González-Nilo, F.; Vasquez, M. In silico analysis of putative paralytic shellfish poisoning toxins export proteins in cyanobacteria. PLoS ONE 2013, 8, e55664. [Google Scholar] [CrossRef]
- Casero, M.C.; Ballot, A.; Agha, R.; Quesada, A.; Cirés, S. Characterization of saxitoxin production and release and phylogeny of sxt genes in paralytic shellfish poisoning toxin-producing Aphanizomenon gracile. Harmful Algae 2014, 37, 28–37. [Google Scholar] [CrossRef]
- Dias, E.; Pereira, P.; Franca, S. Production of paralytic shellfish toxins by Aphanizomenon sp LMECYA 31 (Cyanobacteria). J. Phycol. 2002, 38, 705–712. [Google Scholar] [CrossRef]
- Soto-Liebe, K.; Méndez, M.A.; Fuenzalida, L.; Krock, B.; Cembella, A.; Vásquez, M. PSP toxin release from the cyanobacterium Raphidiopsis brookii D9 (Nostocales) can be induced by sodium and potassium ions. Toxicon 2012, 60, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Ongley, S.E.; Pengelly, J.J.; Neilan, B.A. Elevated Na+ and pH influence the production and transport of saxitoxin in the cyanobacteria Anabaena circinalis AWQC131C and Cylindrospermopsis raciborskii T3. Environ. Microbiol. 2016, 18, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Stucken, K.; John, U.; Cembella, A.; Soto-Liebe, K.; Vásquez, M. Impact of nitrogen sources on gene expression and toxin production in the diazotroph Cylindrospermopsis raciborskii CS-505 and non-diazotroph Raphidiopsis brookii D9. Toxins 2014, 6, 1896–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukenik, A.; Hadas, O.; Kaplan, A.; Quesada, A. Invasion of Nostocales (Cyanobacteria) to subtropical and temperate freshwater lakes—Physiological, regional and global driving forces. Front. Microbiol. 2012, 3, 86. [Google Scholar] [CrossRef] [PubMed]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velázquez, D.; Casero, M.C.; Quesada, A. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing Nostocales cyanobacteria from Mediterranean Europe (Spain). Appl. Environ. Microbiol. 2014, 80, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Mehnert, G.; Leunert, F.; Cirés, S.; Jöhnk, K.D.; Rücker, J.; Nixdorf, B.; Wiedner, C. Competitiveness of invasive and native cyanobacteria from temperate freshwaters under various light and temperature conditions. J. Plankton Res. 2010, 32, 1009–1021. [Google Scholar] [CrossRef]
- Perini, F.; Galluzzi, L.; Dell’Aversano, C.; Iacovo, E.D.; Tartaglione, L.; Ricci, F.; Forino, M.; Ciminiello, P.; Penna, A. SxtA and sxtG gene expression and toxin production in the mediterranean Alexandrium minutum (Dinophyceae). Mar. Drugs 2014, 12, 5258–5276. [Google Scholar] [CrossRef] [PubMed]
- Barón-Sola, Á.; Gutiérrez-Villanueva, M.A.; del Campo, F.F.; Sanz-Alférez, S. Characterization of aphanizomenon ovalisporum amidinotransferase involved in cylindrospermopsin synthesis. Microbiol. Open 2013, 2, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Muenchhoff, J.; Siddiqui, K.S.; Poljak, A.; Raftery, M.J.; Barrow, K.D.; Neilan, B.A. A novel prokaryotic L-arginine: Glycine amidinotransferase is involved in cylindrospermopsin biosynthesis. FEBS J. 2010, 277, 3844–3860. [Google Scholar] [CrossRef] [PubMed]
- Agha, R.; Cirés, S.; Wörmer, L.; Domínguez, J.A.; Quesada, A. Multi-scale strategies for the monitoring of freshwater cyanobacteria: Reducing the sources of uncertainty. Water Res. 2012, 46, 3043–3053. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Hisbergues, M.; Börner, T.; Dittmann, E.; Neilan, B.A. Inactivation of an ABC transporter gene, mcyH, results in loss of microcystin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Appl. Environ. Microbiol. 2004, 70, 6370–6378. [Google Scholar] [CrossRef] [PubMed]
- Burse, A.; Weingart, H.; Ullrich, M.S. Norm, an Erwinia amylovora multidrug efflux pump involved in in vitro competition with other epiphytic bacteria. Appl. Environ. Microbiol. 2004, 70, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Bengoechea, J.A.; Skurnik, M. Temperature-regulated efflux pump/potassium antiporter system mediates resistance to cationic antimicrobial peptides in Yersinia. Mol. Microbiol. 2000, 37, 67–80. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, P.M.; Moffitt, M.C.; Neilan, B.A. Current knowledge of paralytic shellfish toxin biosynthesis, molecular detection and evolution. In Toxins and Biologically Active Compounds from Microalgae; Rossini, G.P., Ed.; CRC Press: Boca Raton, FL, USA, 2014; Volume 1, pp. 251–280. [Google Scholar]
- Zhang, Y.; Zhang, S.-F.; Lin, L.; Wang, D.-Z. Whole transcriptomic analysis provides insights into molecular mechanisms for toxin biosynthesis in a toxic dinoflagellate Alexandrium catenella (ACHK-T). Toxins 2017, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, P.M.; Song, X.; Neilan, B.A.; Moffitt, M.C. Comparative proteomics reveals that a saxitoxin-producing and a nontoxic strain of Anabaena circinalis are two different ecotypes. J. Proteome Res. 2014, 13, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Velzeboer, R.M.; Baker, P.D.; Rositano, J. Saxitoxins associated with the growth of the cyanobacterium Anabaena circinalis (Nostocales, Cyanophyta) under varying sources and concentrations of nitrogen. Phycologia 2001, 40, 305–312. [Google Scholar] [CrossRef]
- Yunes, J.S.; De La Rocha, S.; Giroldo, D.; Silveira, S.B.D.; Comin, R.; Bicho, M.D.S.; Melcher, S.S.; Sant’Anna, C.L.; Vieira, A.A.H. Release of carbohydrates and proteins by a subtropical strain of Raphidiopsis brookii (Cyanobacteria) able to produce saxitoxin at three nitrate concentrations. J. Phycol. 2009, 45, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Marker, A.F.; Nush, E.A.; Rai, H.; Riemann, B. The measurement of photosynthetic pigments in Δfreshwaters and standardization of methods: Conclusions and recommendations. Arch. Hydrobiol. 1980, 14, 91–106. [Google Scholar]
- Prado, R.; Rioboo, C.; Herrero, C.; Cid, Á. Screening acute cytotoxicity biomarkers using a microalga as test organism. Ecotoxicol. Environ. Safe 2012, 86, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, S.; Llaneza, V.; Pulido-Reyes, G.; Fernández-Piñas, F.; Bonzongo, J.C.; Leganes, F.; Rosal, R.; García-Calvo, E.; Rodea-Palomares, I. A colloidal singularity reveals the crucial role of colloidal stability for nanomaterials in vitro toxicity testing: nZVI-microalgae colloidal system as a case study. PLoS ONE 2014, 9, e109645. [Google Scholar] [CrossRef] [PubMed]
- Rodea-Palomares, I.; Makowski, M.; Gonzalo, S.; González-Pleiter, M.; Leganés, F.; Fernández-Piñas, F. Effect of PFOA/PFOS pre-exposure on the toxicity of the herbicides 2, 4-D, Atrazine, Diuron and Paraquat to a model aquatic photosynthetic microorganism. Chemosphere 2015, 139, 65–72. [Google Scholar] [CrossRef] [PubMed]
- NCBI Genbank. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genbank/ (accessed on 20 April 2017).
- European Nucleotide Archive. Available online: http://www.ebi.ac.uk/ena (accessed on 20 April 2017).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2−ΔΔct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Applied Biosystems. Guide to Performing Relative Quantitation of Gene Expression Using Real-Time Quantitative PCR; Applied Biosystems: Foster City, CA, USA, 2004. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Growth Rate (day−1) | Chl a Content (mg g−1 DW) |
|---|---|---|
| 12 | 0.10 ± 0.02 a | 8.6 ± 1.6 a |
| 23 | 0.17 ± 0.03 b | 13.6 ± 2.4 b |
| 30 | 0.07 ± 0.01 a | 9.9 ± 3.1 a |
| Temperature (°C) | Total STX Content (µg mg−1 DW) | Total STX Content (µg mg−1 Chl a) | Extracellular STX (% of Total STX) |
|---|---|---|---|
| 12 | 0.08 ± 0.03 a | 8.7 ± 2.2 a | 16.9 ± 10.2 a |
| 23 | 0.20 ± 0.08 b | 14.0 ± 4.3 a | 11.8 ± 6.1 a |
| 30 | 0.25 ± 0.07 b | 28.6 ± 13.7 b | 53.8 ± 15.5 b |
| Culture Medium | Ecophysiology | Toxin Production/Release | Relative Expression | ||||
|---|---|---|---|---|---|---|---|
| Growth Rate (day−1) | Chl a Content (mg g−1 DW) | Total STX Content (µg mg−1 DW) | Extracellular STX (%) | sxtA | sxtM | sxtPer | |
| BG11 | 0.17 ± 0.03 | 13.6 ± 2.4 | 0.20 ± 0.08 | 11.8 ± 6.1 | 1.0 ± 0.01 | 1.0 ± 0.02 | 1.0 ± 0.01 |
| BG110 | 0.05 ± 0.01 * | 7.6 ± 1.9 * | 0.15 ± 0.04 | 51.2 ± 20.2 * | 1.5 ± 0.5 | 1.2 ± 0.5 | 1.1 ± 0.3 |
| Organism | Factor | Effect on Extracellular STX Release | Effect on the Expression of STX Transporters | Reference | |
|---|---|---|---|---|---|
| sxtM | sxtPer | ||||
| Aphanizomenon gracile | Temperature (12°C and 30°C vs. 23 °C) | 4-fold higher at 30 °C than at 23 °C | Upregulation (2.9–4.3X) at 30 °C Upregulation (3.6–3.8X) at 12 °C | Upregulation (2.1X) at 30 °C | This study |
| Nitrate (absence—BG110- vs. presence—BG11-) | 4-fold higher without nitrate (BG110) | NS | NS | This study | |
| Anabaena circinalis | Ph (9 vs. 7) | Higher at pH 9 (exact value not provided) | Downregulation at pH 9 (59X) vs. pH 7 | NP | [18] |
| Na+ (10 mM vs. 1.3 mM 1) | Higher at 10 mM (exact value not provided) | Downregulation at 10 mM (2.7X) vs. 1.3 mM | NP | [18] | |
| Cylindrospermopsis raciborskii | pH (9 vs. 7) | Higher at pH 9 (exact value not provided) | Upregulation at pH = 9 (24X) vs. pH 7 | NP | [18] |
| Na+ (10 mM vs. 1.3 mM 1) | Higher at 10 mM (exact value not provided) | Upregulation at 10 mM (2.7X) vs. 1.3 mM | NP | [18] | |
| Gene | Primer | Sequence (5’–3’) | Amplicon Size (bp) | Slope; R2 | Efficiency (%) 1 | Source |
|---|---|---|---|---|---|---|
| 16S rRNA | q16grF q16grR | GAGAGACTGCCGGTGACAAA TGCCCTTTGTCCGTAGCATT | 106 | −3.24 0.995 | 103 | This study |
| sxtA | jrtPKSF jrtPKSR | GGAGTGGATTTCAACACCAGAA GTTTCCCAGACTCGTTTCAGG | 147 | −3.38 0.999 | 98 | [18] |
| sxtM | qMgrF qMgrR | GAAGCACGAGTCAGCCTACA CAAAGCACCACCAGCCAAAA | 129 | −3.29 0.998 | 101 | This study |
| sxtPer | qPERgrF qPERgrR | CTGGGCGAGACATTTGAGA GCACAGAGACAGGCGAACTA | 116 | −3.37 0.993 | 98 | This study |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cirés, S.; Delgado, A.; González-Pleiter, M.; Quesada, A. Temperature Influences the Production and Transport of Saxitoxin and the Expression of sxt Genes in the Cyanobacterium Aphanizomenon gracile. Toxins 2017, 9, 322. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9100322
Cirés S, Delgado A, González-Pleiter M, Quesada A. Temperature Influences the Production and Transport of Saxitoxin and the Expression of sxt Genes in the Cyanobacterium Aphanizomenon gracile. Toxins. 2017; 9(10):322. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9100322
Chicago/Turabian StyleCirés, Samuel, Adrián Delgado, Miguel González-Pleiter, and Antonio Quesada. 2017. "Temperature Influences the Production and Transport of Saxitoxin and the Expression of sxt Genes in the Cyanobacterium Aphanizomenon gracile" Toxins 9, no. 10: 322. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9100322