
Effects of Adding Clostridium sp. WJ06 on Intestinal Morphology and Microbial Diversity of Growing Pigs Fed with Natural Deoxynivalenol Contaminated Wheat

Abstract
:1. Introduction
2. Results
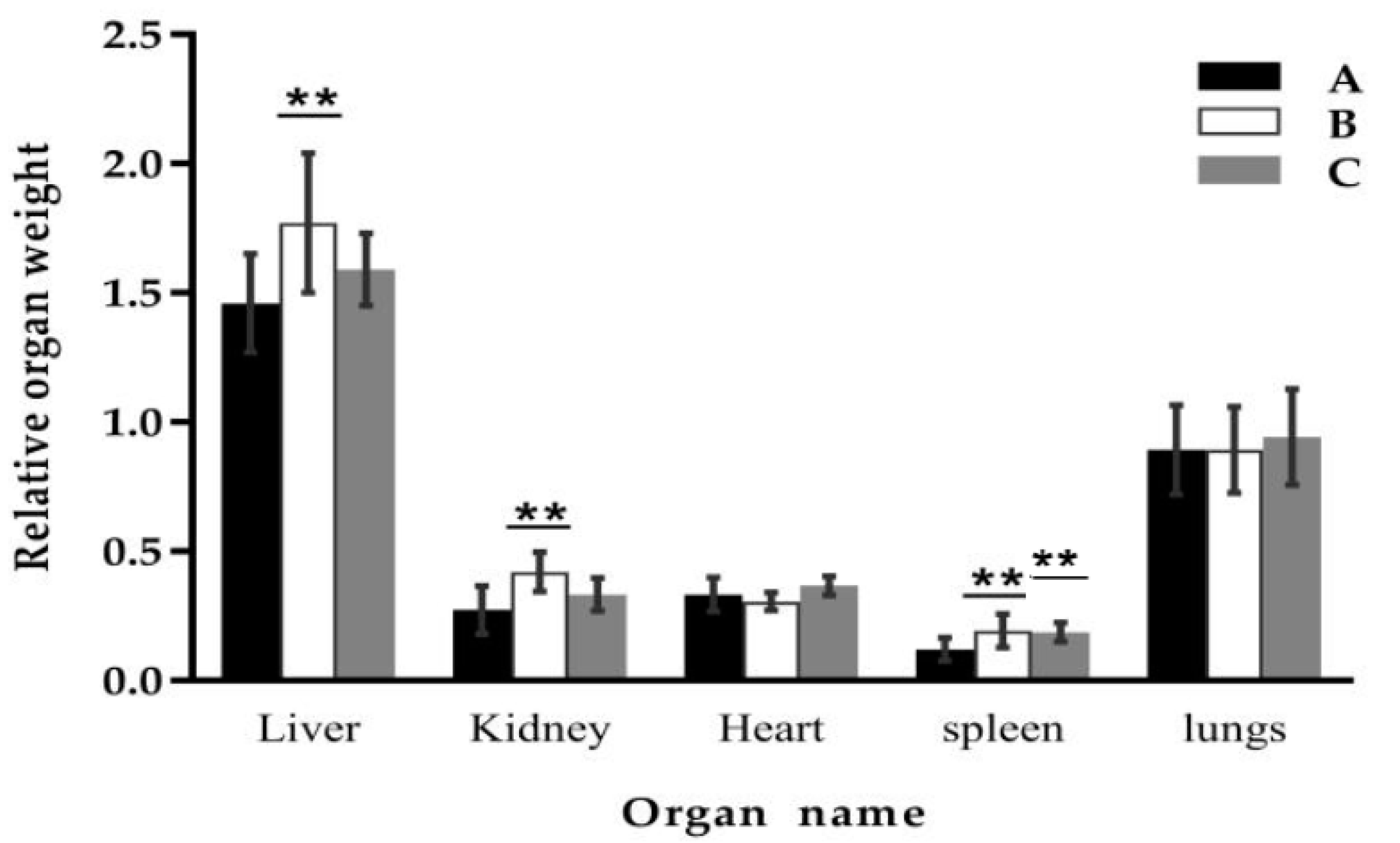
2.1. Effect of Clostridium sp. WJ06 on Growth Performance and Relative Organ Weight of Growing Pigs
2.2. Effects of Clostridium sp. WJ06 on Intestinal Morphology of Growing Pigs
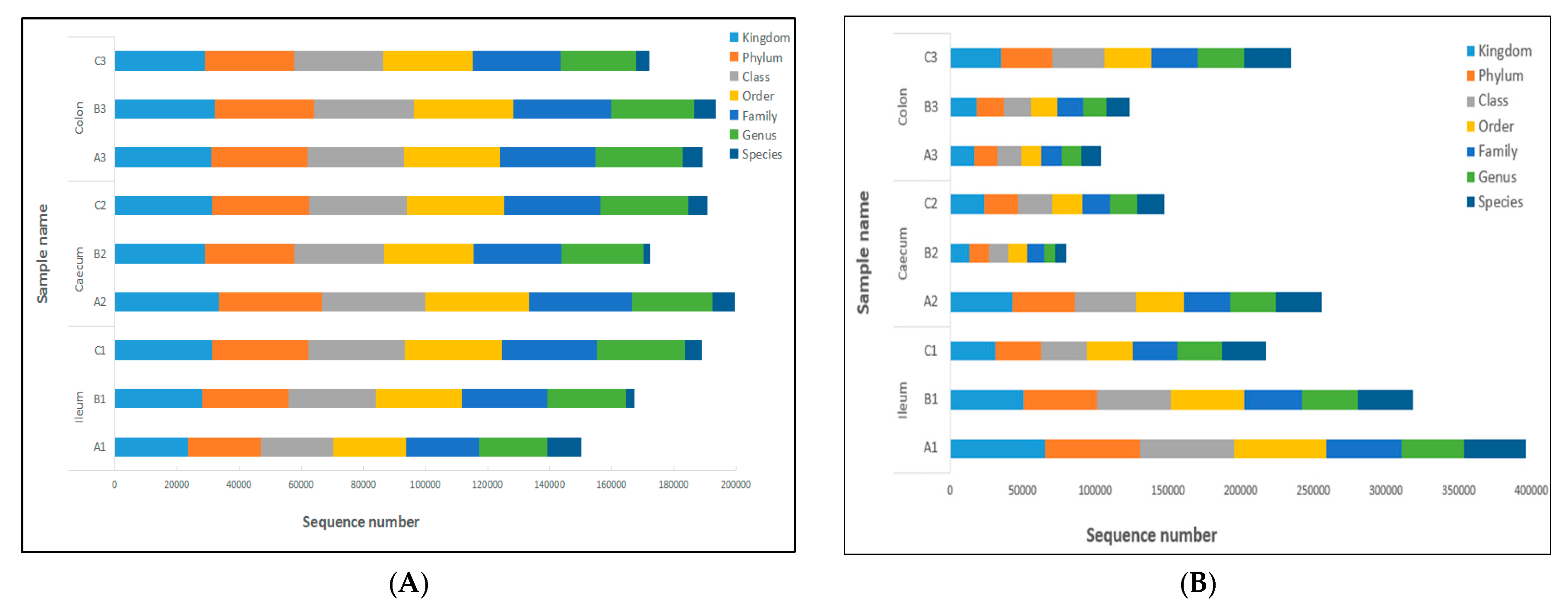
2.3. Analysis of Operational Taxonomic Units (OTUs) and the Alpha Diversity of Bacteria Flora and Fungal Flora in all Samples
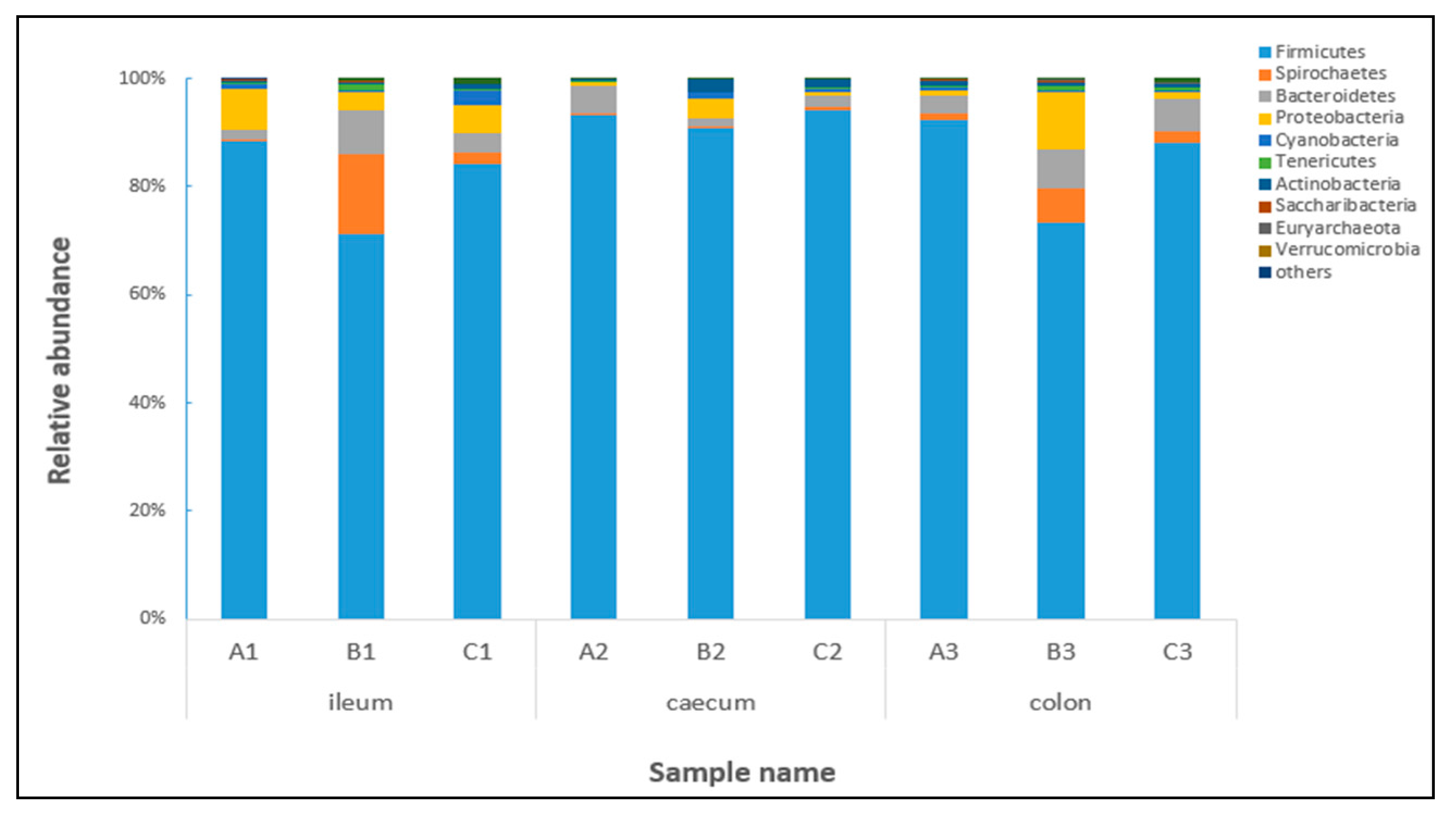
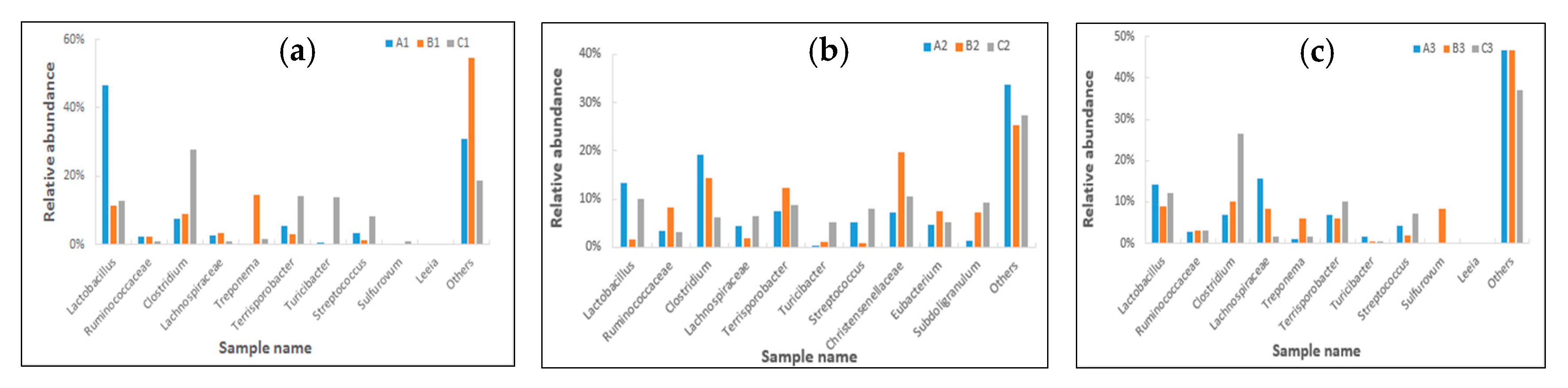
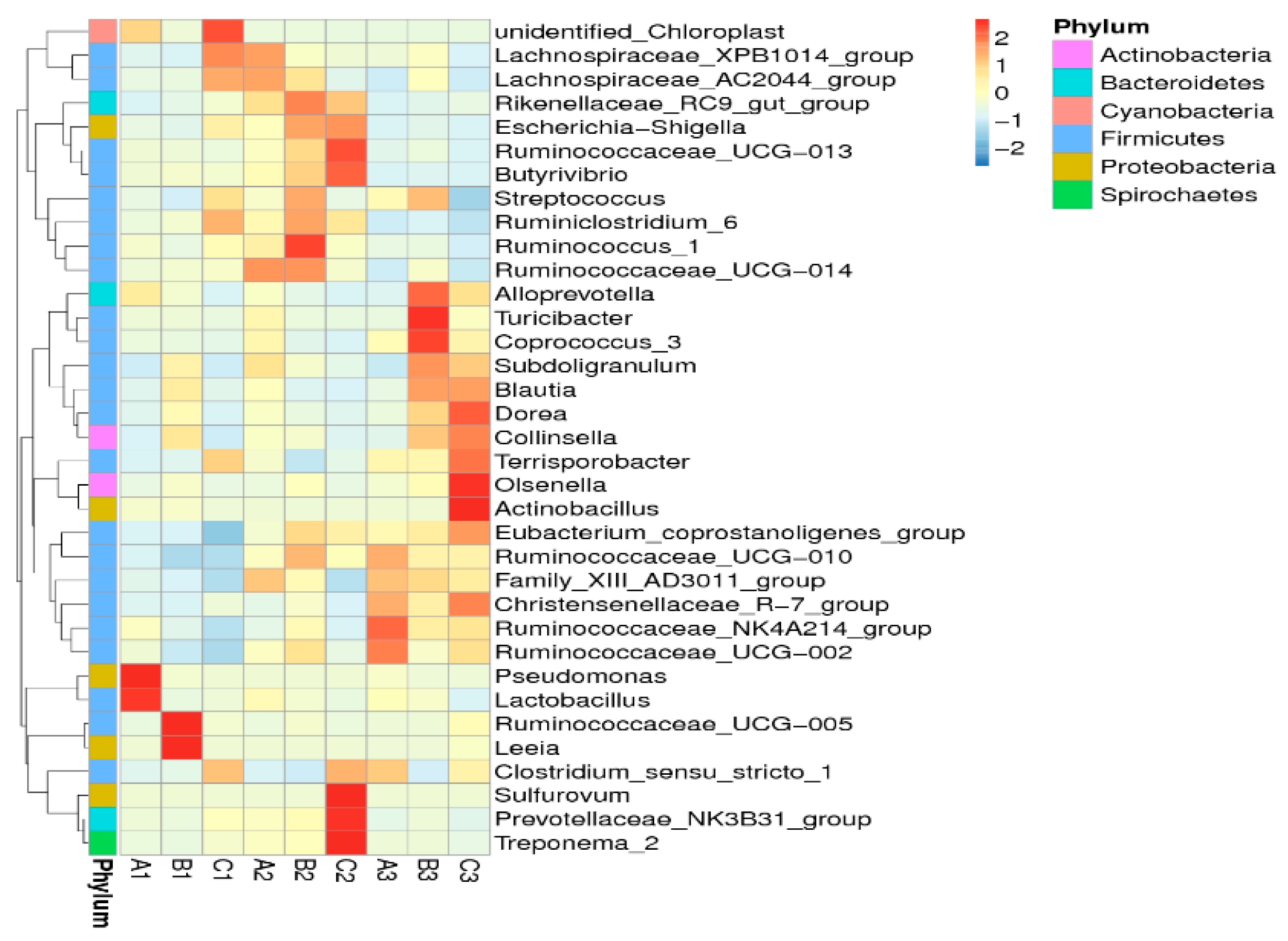
2.4. Effects of Clostridium sp. WJ06 on Intestinal Bacterial Flora of Pigs via 16S rRNA Gene Sequencing
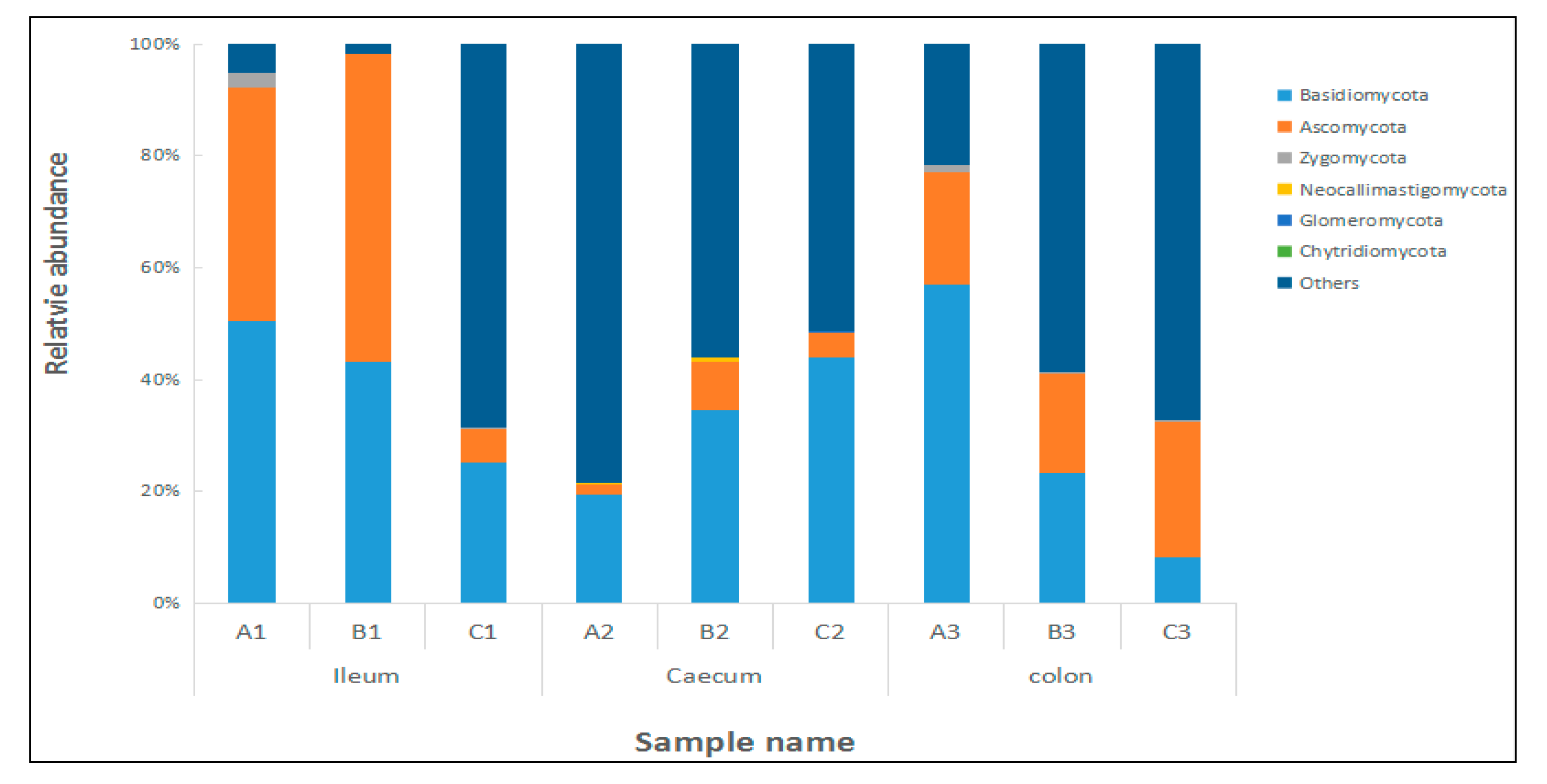
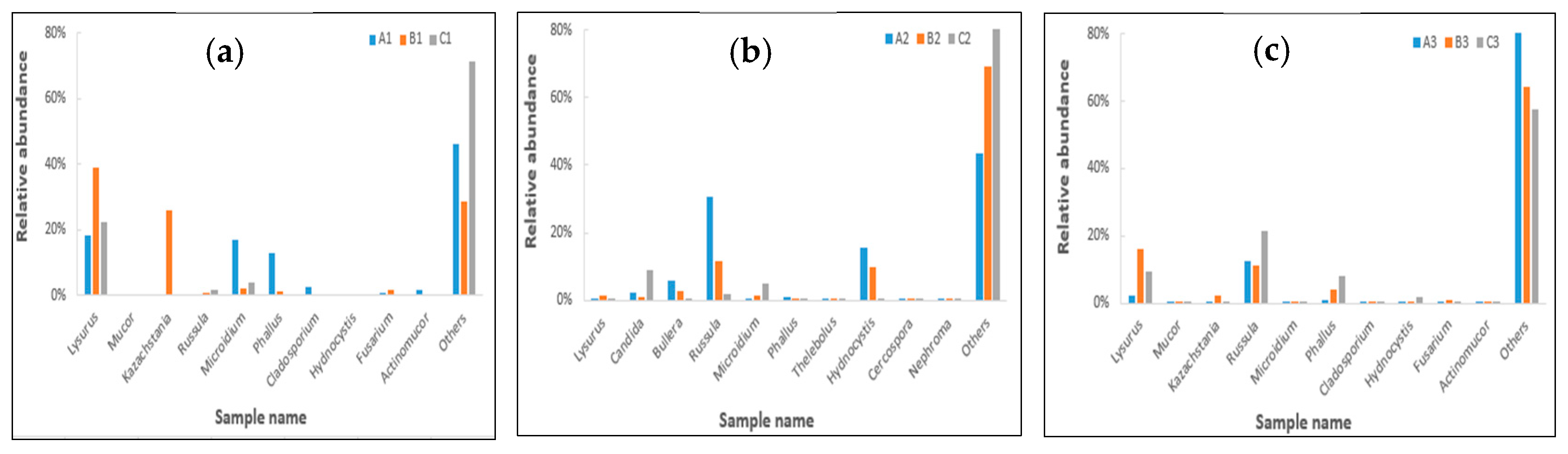
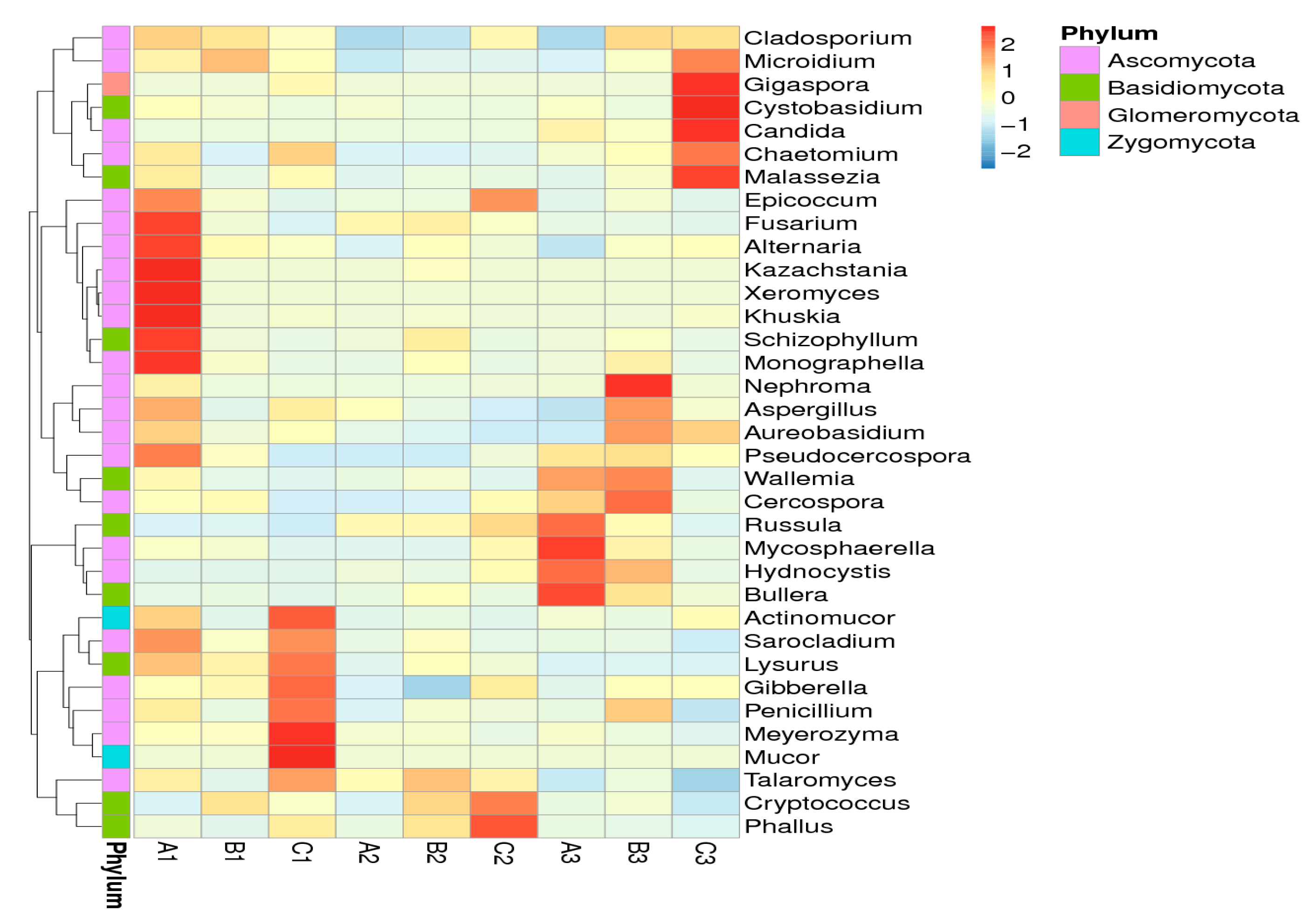
2.5. Effects of Clostridium sp. WJ06 on Intestinal Bacterial Flora of Pigs via ITS1 Gene Sequence
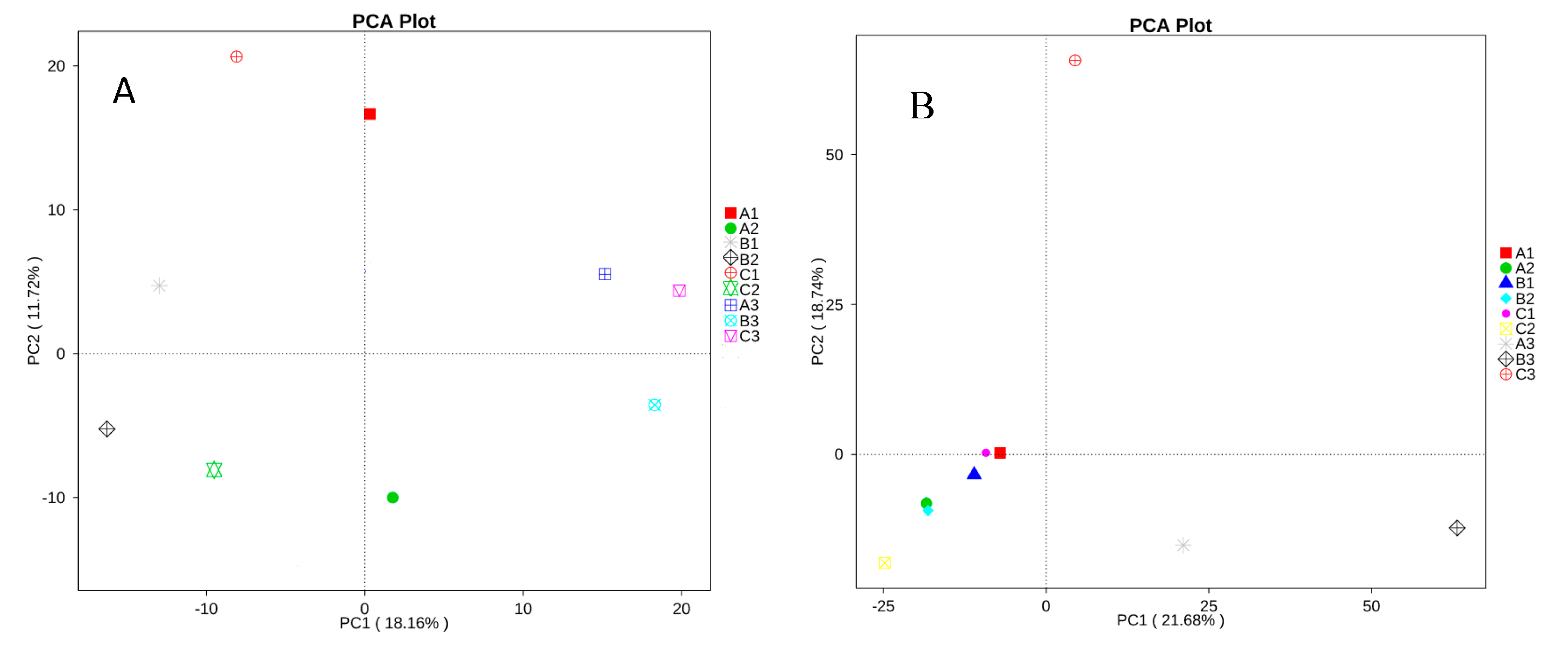
2.6. Analysis of Beta Diversity of all Samples
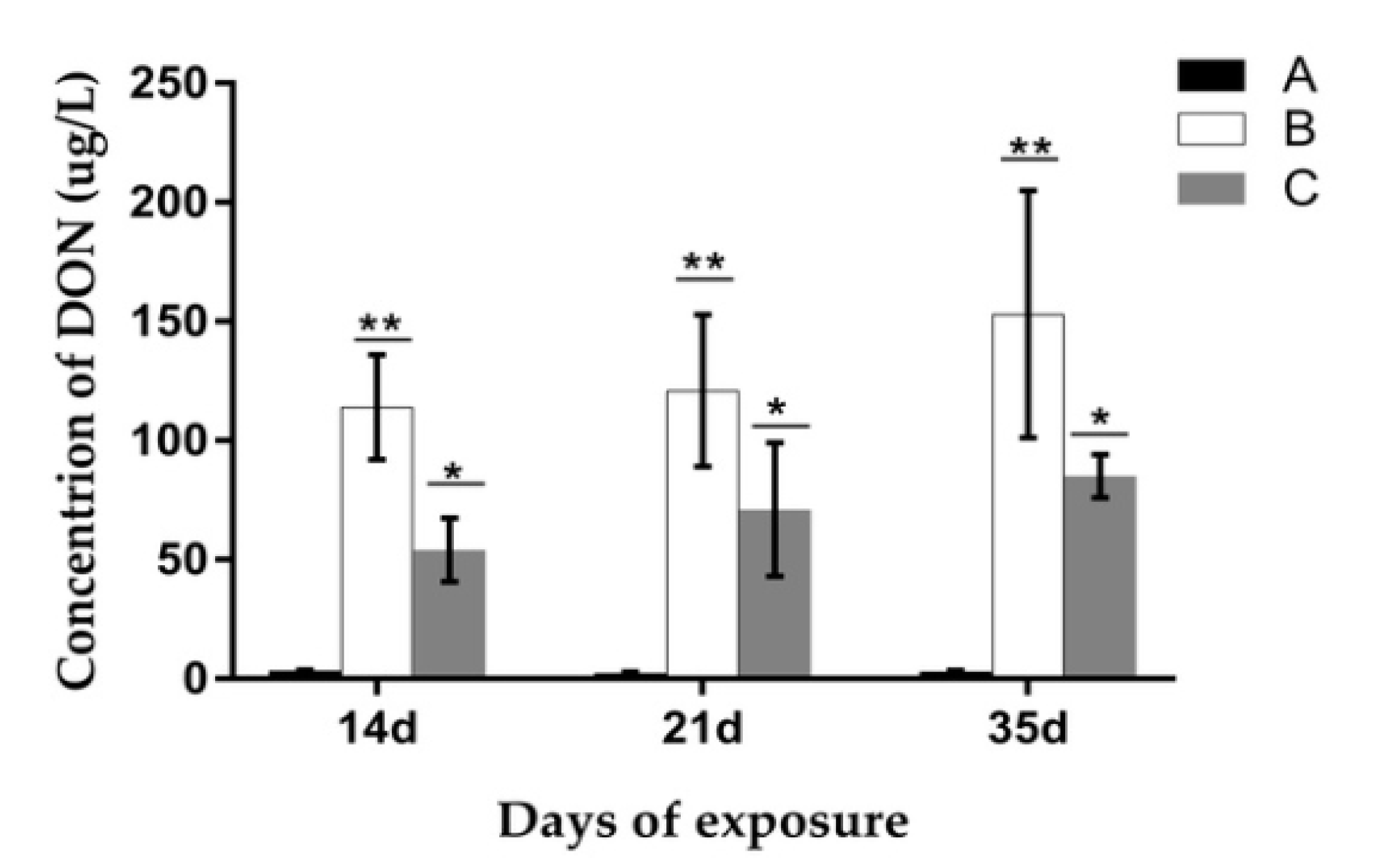
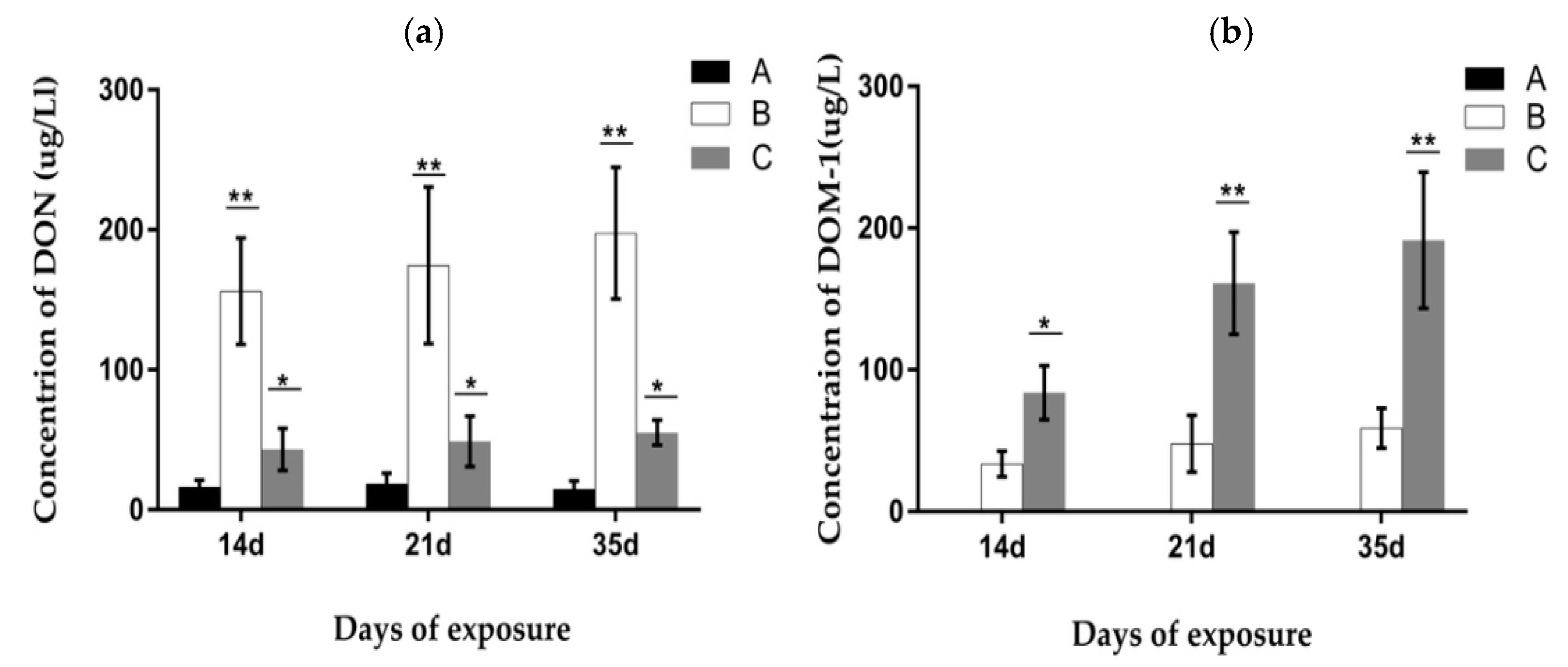
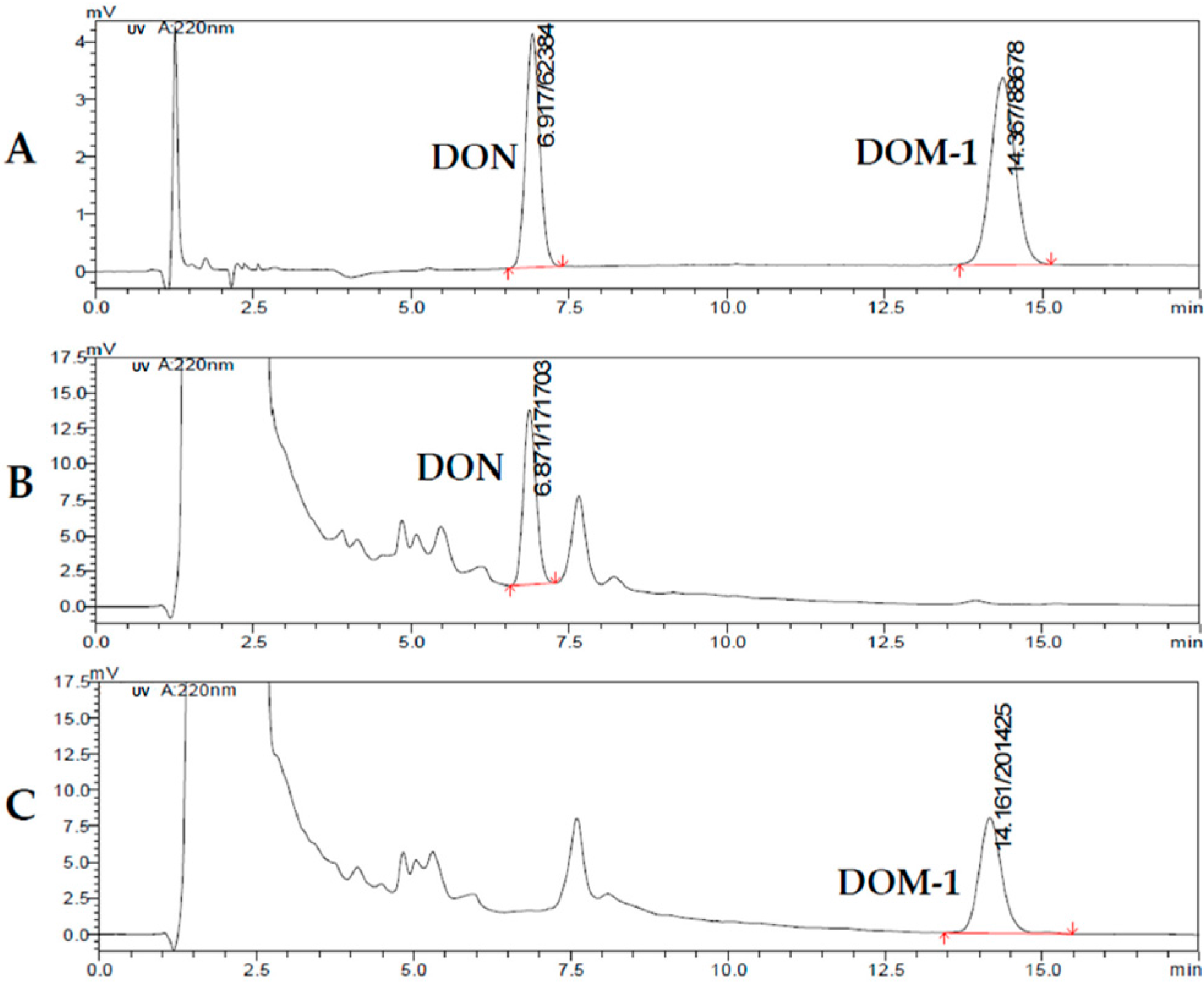
2.7. Analysis of the Concentration of DON and De-Epoxy-DON (DOM-1) in Urine and Feces
3. Discussion
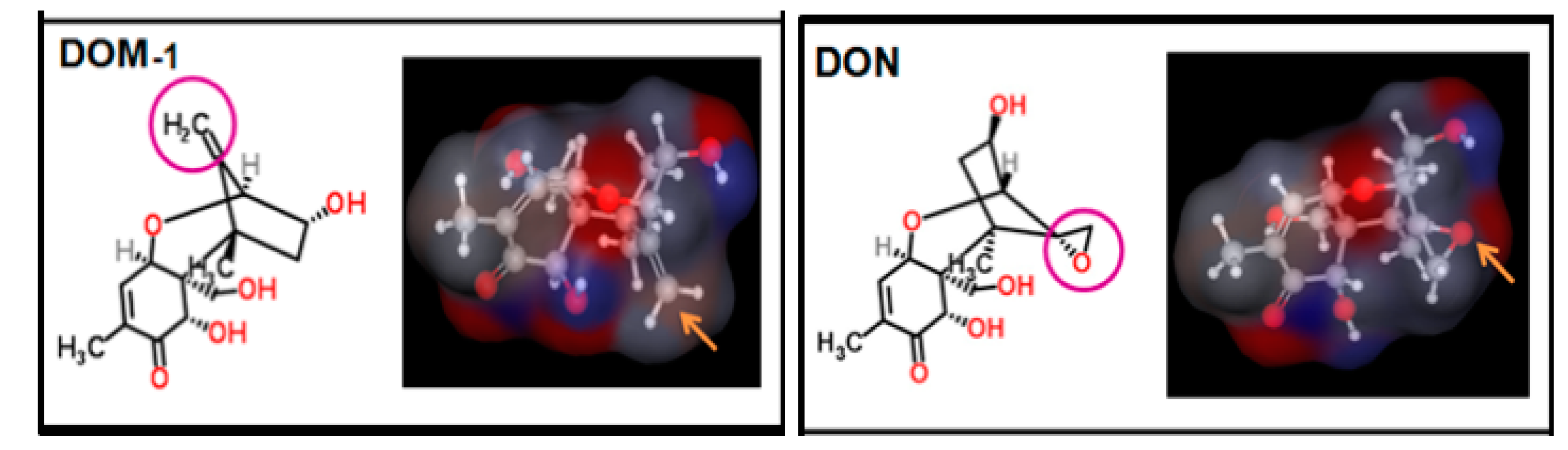
3.1. DON Metabolism and the Toxic Effects of DON
3.2. Intestinal Morphological Structural Integrity and Toxic Effects of DON
3.3. Effects of Clostridium sp. WJ06 on Intestinal Microflora Diversity
4. Materials and Methods
4.1. Ethics Statement
4.2. Bacteria Culture
4.3. Diets, Animals, and Experimental Design
4.4. DNA Extraction
4.5. MiSeq Sequencing of 16S rRNA Gene and ITS1 Gene
4.6. Bioinformatics Analysis
4.7. Analysis of the Concentration of DON and DOM-1 in Urine and Feces
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rohweder, D.; Kersten, S.; Valenta, H.; Sondermann, S.; Schollenberger, M.; Drochner, W.; Dänicke, S. Bioavailability of the Fusarium toxin deoxynivalenol (DON) from wheat straw and chaff in pigs. Arch. Anim. Nutr. 2013, 67, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Garda-Buffon, J.; Kupsk, L.; Badiale-Furlong, E. Deoxynivalenol (DON) degradation and peroxidase enzyme activity in submerged fermentation. Ciênc. Tecnol. Aliment. 2011, 31, 198–203. [Google Scholar] [CrossRef]
- Fodor, J.S. Individual and Combined Effects of Subchronic Exposure of Three Fusarium Toxins (Fumonisin B, Deoxynivalenol and Zearalenone) in Rabbit Bucks. J. Toxicol. Clin. Toxicol. 2015, 5, 2–11. [Google Scholar] [CrossRef]
- Kachlek, M.J.; Szabó-Fodor, A.; Szabó, I.; Bors, C.; Celia, Z.; Gerencsér, Z.; Matics, Z.; Szendrő, T.; Tuboly, E.; Balogh-Zándoki, R.; et al. Subchronic exposure to deoxynivalenol exerts slight effect on the immune system and liver morphology of growing rabbits. Acta Vet. Brno 2017, 86, 37–44. [Google Scholar] [CrossRef]
- Pestka, J.J. Mechanisms of Deoxynivalenol-Induced Gene Expression and Apoptosis. Food Addit. Contam. 2008, 25, 1128–1140. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.A.; Böhm, J.Q.; Zebeli, K.G. Impacts of the feed contaminant deoxynivalenol on the intestine of monogastric animals: Poultry and swine. J. Appl. Toxicol. 2015, 35, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Lewczuk, B.B.; Przybylska-Gornowicz, M.; Gajecka, K.; Targonska, N.; Ziolkowska, M.; Gajecki, M. Histological structure of duodenum in gilts receiving low doses of zearalenone and deoxynivalenol in feed. Exp. Toxicol. Pathol. 2016, 68, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.A.; Ramos, I.; Castellano, E.; Martinez, V.; Martinez-Larranaga, M.; Anadon, M.R.; Martinez, M.A. Mycotoxins modify the barrier function of Caco-2 cells through differential gene expression of specific claudin isoforms: Protective effect of illite mineral clay. Toxicology 2016, 353, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.L.; Chen, Z.; Peng, A.K.; Nussler, Q.; Wu, L.; Liu, W.Y. Mechanism of deoxynivalenol effects on the reproductive system and fetus malformation: Current status and future challenges. Toxicol. In Vitro 2017, 41, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of two mycotoxins deoxynivalenol and fumonisin on pig intestinal health. Porc. Health Manag. 2016, 2, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Du, K.; Wang, C.; Liu, P.; Li, Y.; Ma, X. Effects of Dietary Mycotoxins on Gut Microbiome. Protein Pept. Lett. 2017, 24, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Burel, C.; Tanguy, M.; Guerre, P.; Boilletot, E.; Cariolet, R.; Queguiner, M.; Postollec, G.; Pinton, P.; Salvat, G.; Oswald, I.P.; et al. Effect of low dose of fumonisins on pig health: Immune status, intestinal microbiota and sensitivity to Salmonella. Toxins 2013, 5, 841–864. [Google Scholar] [CrossRef] [PubMed]
- Wache, Y.J.; Valat, C.; Postollec, G.; Bougeard, S.; Burel, C.; Oswald, I.P.; Fravalo, P. Impact of deoxynivalenol on the intestinal microflora of pigs. Int. J. Mol. Sci. 2009, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hathout, A.S.; Aly, S.E. Biological detoxification of mycotoxins: A review. Ann. Microbiol. 2014, 64, 905–919. [Google Scholar] [CrossRef]
- Repečkienė, J.L.; Levinskaitė, A.; Paškevičius, V.R. Toxin-producing fungi on feed grains and application of yeasts for their detoxification. Pol. J. Vet. Sci. 2013, 16, 391–393. [Google Scholar] [CrossRef]
- Oliveira, P.B.; Brosnan, F.; Jacob, A.; Furey, A.; Coffey, E.; Zannini, E.; Arendt, K. Lactic acid bacteria bioprotection applied to the malting process. Part II: Substrate impact and mycotoxin reduction. Food Control 2015, 51, 444–452. [Google Scholar] [CrossRef]
- Chang, C.; Zhou, S.N.; Wang, X.D.; Wu, J.E. Protective Effect of Saccharomyces boulardii on Deoxynivalenol-Induced Injury of Porcine Macrophage via Attenuating p38 MAPK Signal Pathway. Appl. Biochem. Biotechnol. 2017, 182, 411–427. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Valenta, H.; Döll, S. On the toxicokinetics and the metabolism of deoxynivalenol (DON) in the pig. Arch. Anim. Nutr. 2004, 58, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Thanner, S.; Czeglédi, L.; Schwartz-Zimmermann, H.E.; Berthiller, A.F.; Gutzwiller, A. Urinary deoxynivalenol (DON) and zearalenone (ZEA) as biomarkers of DON and ZEA exposure of pigs. Mycotoxin Res. 2016, 32, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, W.; Huang, R.; Cui, Z.; He, L.; Yin, J.; Duan, L.; Li, T.; Wang, J.Q. Deoxynivalenol residues in edible tissue of infested pig. J. Food Agric. Environ. 2013, 11, 1129–1133. [Google Scholar]
- Danicke, S.; Brezina, U. Kinetics and metabolism of the Fusarium toxin deoxynivalenol in farm animals: Consequences for diagnosis of exposure and intoxication and carry over. Food Chem. Toxicol. 2013, 60, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Warth, B.S.; Fruhmann, M.; Mikula, P.; Berthiller, H.; Schuhmacher, F.; Hametner, R.; Abia, C.; Adam, W.A.; Frohlich, G.; Krska, J.R. Development and validation of a rapid multi-biomarker liquid chromatography/tandem mass spectrometry method to assess human exposure to mycotoxins. Rapid Commun. Mass Spectrom. 2012, 26, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol (Review). Toxins 2013, 23, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Diesing, A.K.N.; Panther, C.; Walk, P.; Post, N.; Kluess, A.; Kreutzmann, J.; Danicke, P.; Rothkotter, S.H.; Kahlert, J.S. Mycotoxin deoxynivalenol (DON) mediates biphasic cellular response in intestinal porcine epithelial cell lines IPEC-1 and IPEC-J2. Toxicol. Lett. 2011, 200, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.P.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Akbari, P.B.; Gremmels, S.; Koelink, H.; Verheijden, J.; Garssen, K.A.; Fink-Gremmels, J. Deoxynivalenol: A trigger for intestinal integrity breakdown. FASEB J. 2014, 28, 2414–2429. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, N.D.; Demeyere, M.; Berthiller, K.; Michlmayr, F.; Varga, H.; Adam, E.; Meyer, G.; Croubels, E.S. Comparative in vitro cytotoxicity of modified deoxynivalenol on porcine intestinal epithelial cells. Food Chem. Toxicol. 2016, 95, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Barnett, A.M.; Roy, N.C.; McNabb, W.C.; Cookson, A.L. The interactions between endogenous bacteria, dietary components and the mucus layer of the large bowel. Food Funct. 2012, 3, 690–699. [Google Scholar] [CrossRef] [PubMed]
- Osselaere, A.M.; Devreese, J.; Goossens, V.; Vandenbroucke, S.; De Baere, P.; Croubels, S. Toxicokinetic study and absolute oral bioavailability of deoxynivalenol, T-2 toxin and zearalenone in broiler chickens. Food Chem. Toxicol. 2013, 51, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Frey, J.C.; Pell, A.N.; Berthiaume, R.; Lapierre, H.S.; Lee, J.K.; Ha, J.E.; Angert, E.R. Comparative studies of microbial populations in the rumen, duodenum, ileum and faeces of lactating dairy cows. J. Appl. Microbiol. 2010, 108, 1982–1993. [Google Scholar] [CrossRef] [PubMed]
- Rotter, B.A.; Prelusky, D.B.; Pestks, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, G.S.; Pettersson, H.; Johnsen, K.; Lindberg, J.E. Transformation of trichothecenes in ileal digesta and faeces from pigs. Arch. Anim. Nutr. 2002, 56, 263–274. [Google Scholar] [CrossRef]
- Prelusky, D.B.; Veira, D.M.; Trenholm, H.L. Plasma pharmacokinetics of the mycotoxin deoxynivalenol following oral and intravenous administration to sheep. J. Environ. Sci. Health B 1985, 20, 603–624. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.C.; Savard, K.; Deschene, K.; Lauzon, V.A.; Pinilla, C.A.; Gagnon, J.; Lapointe, F.; Chorfi, Y. Impact of deoxynivalenol (DON) contaminated feed on intestinal integrity and immune response in swine. Food Chem. Toxicol. 2015, 80, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Zoumpopoulou, G.; Folign, B.; Alexandraki, V.; Kazou, M.; Pot, B.; Tsakalidou, E. Discovering probiotic microorganisms: In vitro, in vivo, genetic and omics approaches. Front. Microbiol. 2015, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Frantz, N.; Khoo, C.; Gibson, G.R.; Rastall, R.A.; McCartney, A.L. Investigation of the faecal microbiota associated with canine chronic diarrhoea. FEMS Microbiol. Ecol. 2010, 71, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Arqués, J.L.; Rodríguez, E.; Langa, S.; Landete, J.M.; Medina, M. Antimicrobial activity of lactic acid bacteria in dairy products and gut: Effect on pathogens. Biomed. Res. Int. 2015, 22, 584183. [Google Scholar] [CrossRef] [PubMed]
- Goyal, N.; Rishi, P.; Shukla, G. Lactobacillus rhamnosus GG antagonizes Giardia intestinalis induced oxidative stress and intestinal disaccharidases: An experimental study. World J. Microbiol. Biotechnol. 2013, 29, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, C.; Li, F. Impact of dietary fiber/starch ratio in shaping caecal microbiota in rabbits. Can. J. Microbiol. 2015, 61, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Michelland, R.J.S.; Combes, V.; Monteils, L.; Cauquil, T.; Fortun-Lamothe, L. Rapid adaptation of the bacterial community in the growing rabbit caecum after a change in dietary fibre supply. Animal 2011, 5, 1761–1768. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhu, Y.L.; Huang, L.; Li, F.C. The Effect of Lactobacillus isolates on growth performance, immune response, intestinal bacterial community composition of growing Rex Rabbits. J. Anim. Physiol. Anim. Nutr. 2017, 101, e1–e13. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.R.; Geng, L.L.; Li, Y.H.; Wang, Q.; Diao, Q.Y.; Zhou, T.; Dai, P.L. The effects of Bt Cry1Ie toxin on bacterial diversity in the midgut of Apis mellifera ligustica (Hymenoptera: Apidae). Sci. Rep. 2016, 6, 24664. [Google Scholar] [CrossRef] [PubMed]
- McGuire, K.L.; Payne, S.G.; Palmer, M.I.; Gillikin, C.M.; Keefe, D.S.; Kim, J.S.; Gedallovich, M.J.; Discenza, R.; Rangamannar, J.A.; Koshner, A.L.; et al. Digging the New York City Skyline: Soil fungal communities in green roofs and city parks. PLoS ONE 2013, 8, e58020. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–999. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Kersten, S.; Valenta, H.; Hüther, L.; Engelhardt, U.; Dänicke, S. Simultaneous determination of zearalenone, deoxynivalenol and their metabolites in bovine urine as biomarkers of exposure. World Mycotoxin J. 2014, 8, 63–74. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment * | Initial Weight (kg) | Final Weight (kg) | ADG (g) # | ADFI (kg) # | F/G # |
|---|---|---|---|---|---|
| A | 54.38 ± 1.21 | 78.3 ± 1.78 | 885.93±72 | 2.14±0.38 | 2.41 ± 0.32 |
| B | 56.44 ± 1.63 | 71.33 ± 2.98 a | 551.48±46 a | 1.66±0.33 a | 3.01 ± 0.41 a |
| C | 56.06 ± 1.78 | 76.07 ± 3.15 | 741.16 ± 62 | 1.98 ± 0.27 | 2.68 ± 0.36 |
| Sample Name * | Raw PE # | Qualified # | Base (nt) # | AvgLen (nt) # | Effective% # | Chao Ave 2 | Observed Species | Shannon Ave 2 |
|---|---|---|---|---|---|---|---|---|
| A1 | 64,782 | 34,392 | 10,231,113 | 390 | 40.5 | 564.27 | 426 | 4.93 |
| B1 | 69,691 | 41,156 | 14,360,310 | 388 | 53.05 | 496.63 | 380 | 4.66 |
| C1 | 68,531 | 40,201 | 13,531,583 | 384 | 51.41 | 538.44 | 443 | 5.44 |
| A2 | 72,117 | 47,250 | 15,233,409 | 389 | 54.37 | 434.19 | 365 | 5.21 |
| B2 | 64,769 | 45,377 | 14,399,002 | 389 | 57.1 | 436.26 | 389 | 5.90 |
| C2 | 69,357 | 45,332 | 14,537,019 | 385 | 54.4 | 379.02 | 329 | 5.53 |
| A3 | 75,203 | 47,673 | 14,834,644 | 392 | 50.28 | 489.40 | 406 | 4.85 |
| B3 | 68,288 | 41,553 | 14,224,802 | 396 | 52.59 | 468.35 | 382 | 5.05 |
| C3 | 70,715 | 45,665 | 14,628,909 | 386 | 53.59 | 586.57 | 485 | 6.00 |
| Total | 409,247 | 253,708 | 82,292,436 | 2325 | 311 | 4393 | 3605 | 48 |
| Average | 69,272 | 43,177 | 13,997,865 | 389 | 51.92 | 488.13 | 401 | 5.29 |
| Sample Name * | Raw PE # | Qualified # | Base (nt) # | AvgLen (nt)# | Effective% # | Chao Ave 2 | Observed Species | Shannon Ave 2 |
|---|---|---|---|---|---|---|---|---|
| A1 | 123,899 | 72,335 | 18,813,512 | 267 | 56.92 | 798.77 | 611 | 5.18 |
| B1 | 70,194 | 54,056 | 12,893,920 | 246 | 74.80 | 389.73 | 287 | 2.92 |
| C1 | 121,183 | 102,600 | 22,150,221 | 218 | 83.97 | 656.80 | 447 | 3.46 |
| A2 | 103,737 | 79,891 | 17,630,954 | 221 | 76.95 | 843.35 | 570 | 5.07 |
| B2 | 72,542 | 46,157 | 10,630,776 | 234 | 62.49 | 992.45 | 667 | 5.71 |
| C2 | 95,418 | 78,657 | 16,739,208 | 213 | 82.33 | 656.80 | 665 | 3.46 |
| A3 | 78,988 | 64,817 | 12,797,508 | 198 | 81.95 | 959.79 | 748 | 6.87 |
| B3 | 83,591 | 62,692 | 13,180,068 | 211 | 74.72 | 1195.65 | 918 | 6.52 |
| C3 | 72,307 | 44,728 | 10,577,687 | 238 | 61.34 | 886.02 | 769 | 5.96 |
| Total | 821,859 | 605,933 | 135,413,854 | 2046 | 655.47 | 7379.36 | 5681 | 45.14 |
| Average | 91,318 | 67,326 | 15,045,984 | 227 | 72.83 | 819.93 | 631 | 5.02 |
| Basal Diet | Contaminated Diet | |
|---|---|---|
| Ingredients (%) | ||
| Corn | 60.0 | 32.7 |
| Soybean Meal | 23.0 | 18.0 |
| Extruded soybean | 2.0 | 1.0 |
| Contaminated wheat | 0 | 33.3 |
| Concentrate feed # | 15 | 15 |
| Total | 100 | 100 |
| Calculated composition (%) | ||
| Dry matter | 85.4 | 86.62 |
| Crude ash | 4.79 | 4.88 |
| Calcium | 0.63 | 0.58 |
| Available phosphorus | 0.57 | 0.62 |
| Crude fibre | 53.75 | 52.55 |
| Crude protein | 16.67 | 16.70 |
| Crude fat | 3.78 | 2.72 |
| Digestible energy (MJ/kg) | 17.17 | 16.69 |
| Content of mycotoxin (ug/kg) 2 | ||
| DON | 371.2 | 1904.44 |
| Aflatoxin B1 (AFB1) | 1.56 | 1.59 |
| Fumonisin B1 (FB1) | 256.3 | 325.7 |
| Zearalenone (ZEA) | 145.16 | 96.08 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Wang, J.; Huang, L.; Chen, H.; Wang, C. Effects of Adding Clostridium sp. WJ06 on Intestinal Morphology and Microbial Diversity of Growing Pigs Fed with Natural Deoxynivalenol Contaminated Wheat. Toxins 2017, 9, 383. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9120383
Li F, Wang J, Huang L, Chen H, Wang C. Effects of Adding Clostridium sp. WJ06 on Intestinal Morphology and Microbial Diversity of Growing Pigs Fed with Natural Deoxynivalenol Contaminated Wheat. Toxins. 2017; 9(12):383. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9120383
Chicago/Turabian StyleLi, FuChang, JinQuan Wang, LiBo Huang, HongJu Chen, and ChunYang Wang. 2017. "Effects of Adding Clostridium sp. WJ06 on Intestinal Morphology and Microbial Diversity of Growing Pigs Fed with Natural Deoxynivalenol Contaminated Wheat" Toxins 9, no. 12: 383. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9120383