
Distinct Expression of Immunoglobulin-Binding Proteins in Shiga Toxin-Producing Escherichia coli Implicates High Protein Stability and a Characteristic Phenotype


Abstract
:1. Introduction
2. Results
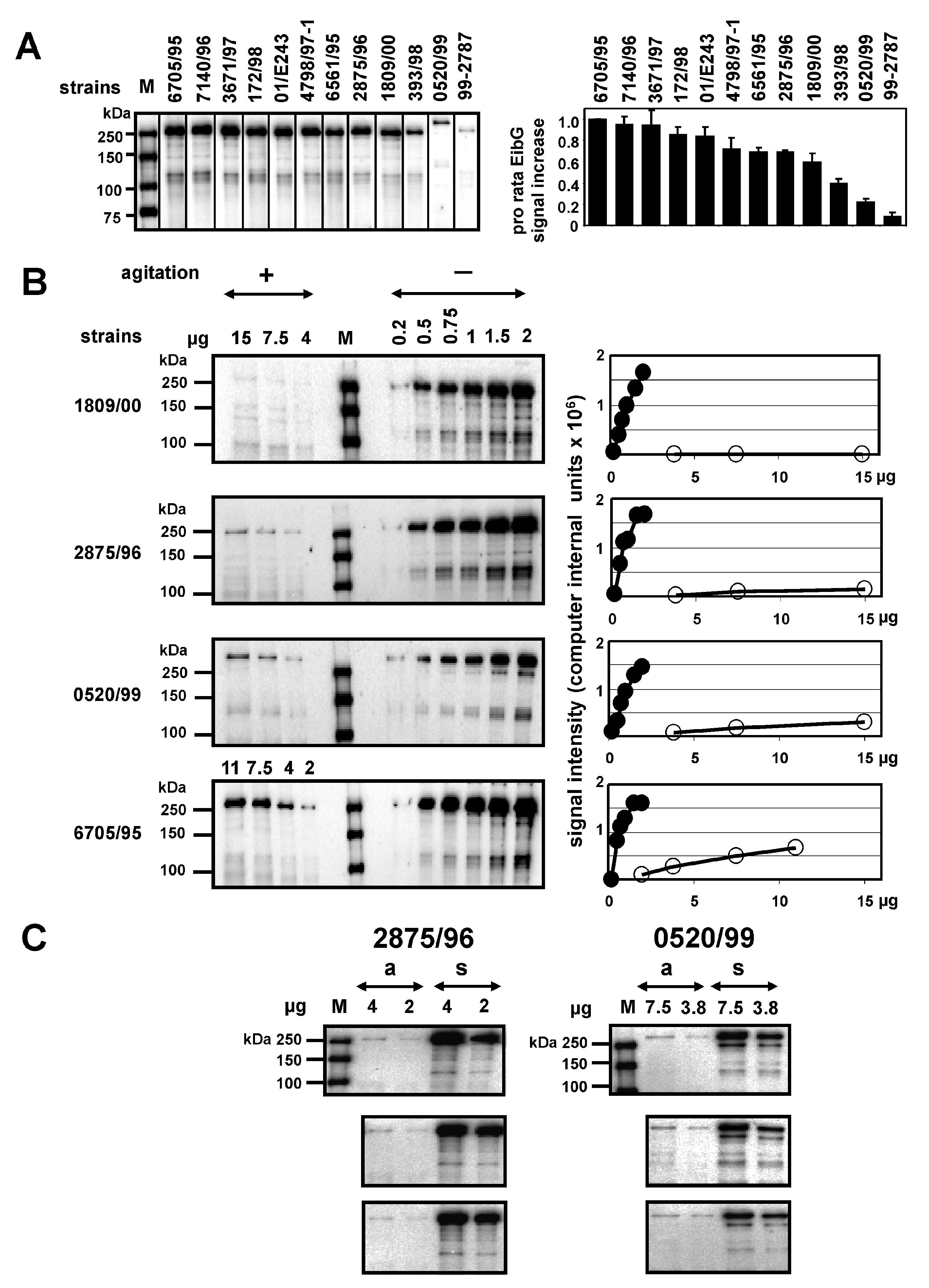
2.1. EibG Is Highly Upregulated Under Static Incubation, yet the Protein Is Differentially Expressed under Shaking and Non-Shaking Conditions among Isolates
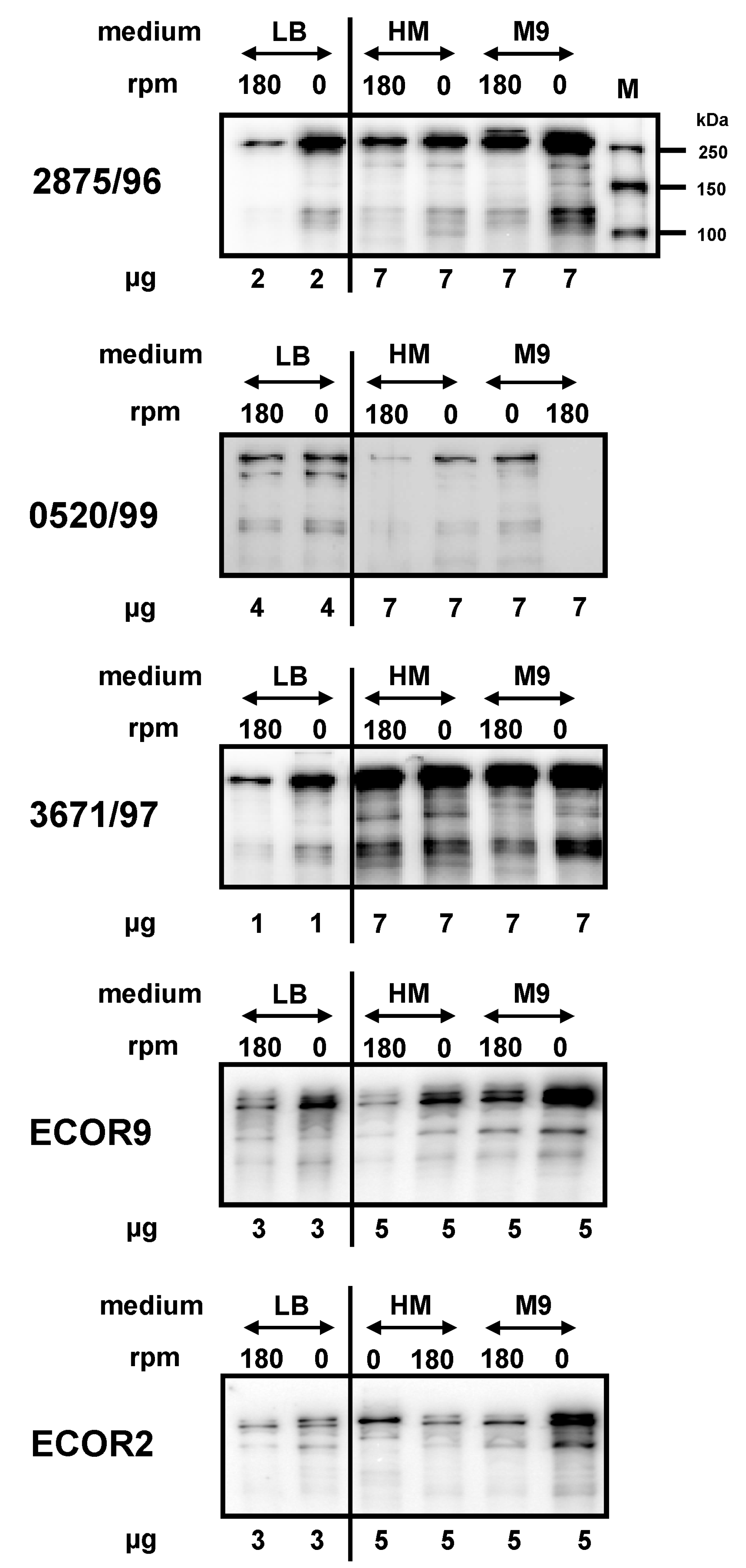
2.2. Impact of Medium Composition on EibG Expression in STEC and Other Eib Proteins in E. coli
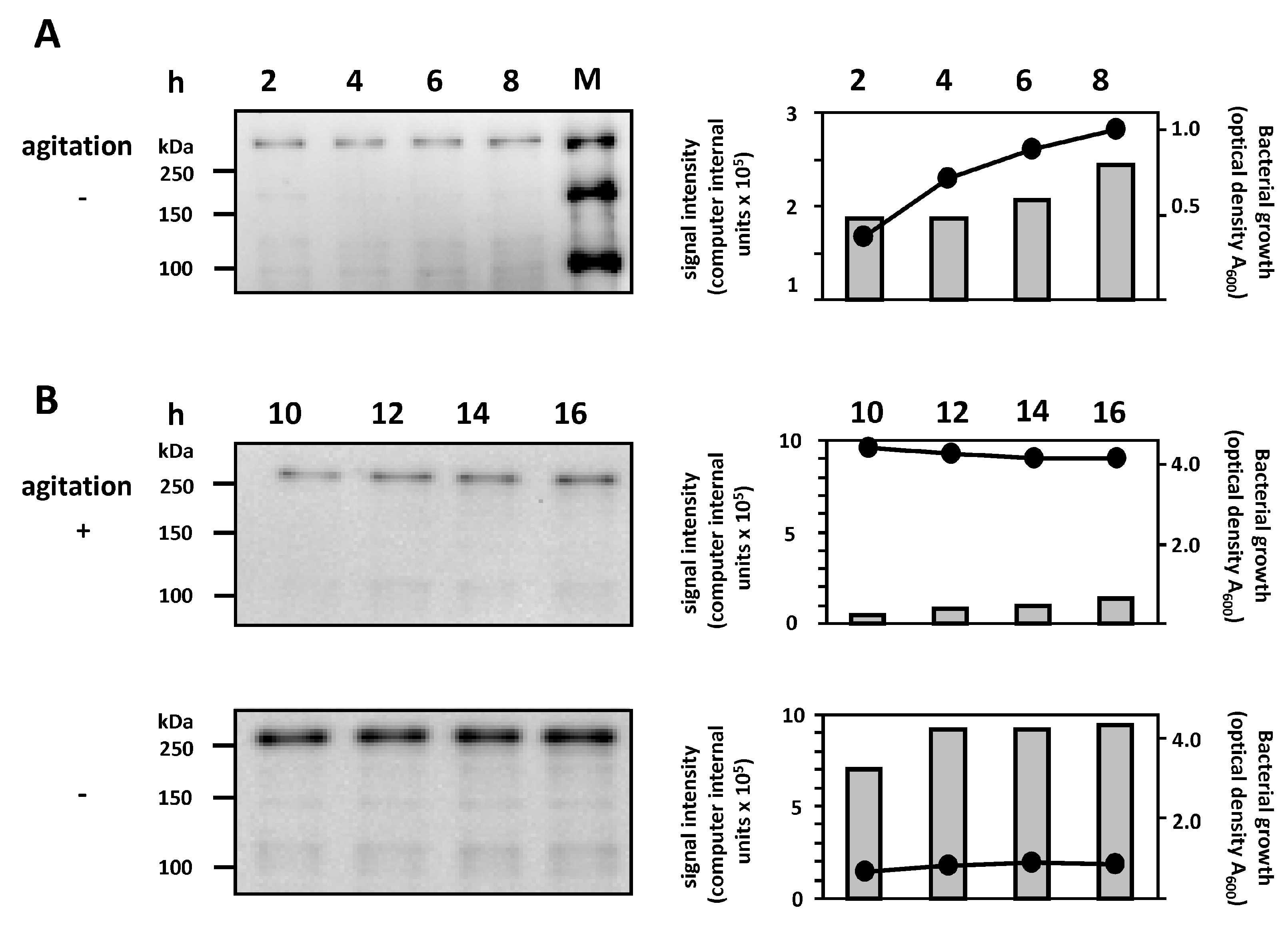
2.3. Correlation between Bacterial Growth and EibG Expression in STEC under Different Growth Conditions
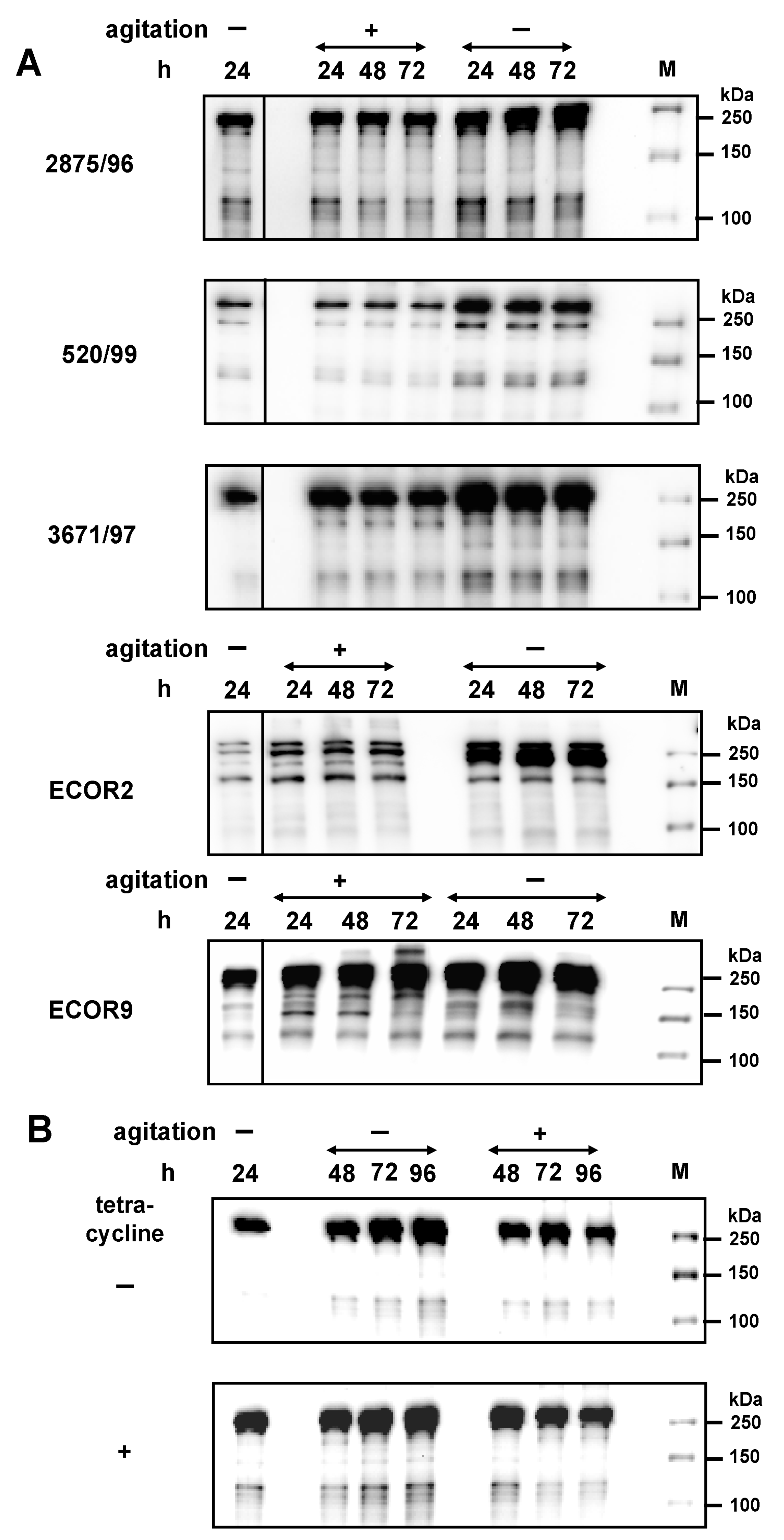
2.4. Regulation and Stability of EibG Proteins in STEC
3. Discussion
4. Experimental Section
4.1. Strains and Culture Conditions
4.2. Immunoblot Analysis and Signal Quantification
4.3. Phenotypic Characterization
4.4. Peptide Mass Fingerprinting
Author Contributions
Conflicts of Interest
References
- Bouvet, J.P. Immunoglobulin Fab fragment-binding proteins. Int. J. Immunopharmacol. 1994, 16, 419–424. [Google Scholar] [CrossRef]
- Sandt, C.H.; Wang, Y.-D.; Wilson, R.A.; Hill, C.W. Escherichia coli strains with nonimmune immunoglobulin-binding activity. Infect. Immun. 1997, 65, 4572–4579. [Google Scholar] [PubMed]
- Sandt, C.H.; Hill, C.W. Four different genes responsible for nonimmune immunoglobulin-binding activities within a single strain of Escherichia coli. Infect. Immun. 2000, 68, 2205–2214. [Google Scholar] [CrossRef] [PubMed]
- Ochman, H.; Selander, R.K. Standard reference strains of Escherichia coli from natural populations. J. Bacteriol. 1984, 157, 690–693. [Google Scholar] [PubMed]
- Sandt, C.H.; Hill, C.W. Nonimmune binding of human immunoglobulin A (IgA) and IgG Fc by distinct sequence segments of the EibF cell surface protein of Escherichia coli. Infect. Immun. 2001, 69, 7293–7303. [Google Scholar] [CrossRef] [PubMed]
- Bardiau, M.; Grégoire, F.; Muylaert, A.; Nahayo, A.; Duprez, J.N.; Mainil, J.; Linden, A. Enteropathogenic (EPEC), enterohemorrhagic (EHEC) and verotoxigenic (VTEC) Escherichia coli in wild cervids. J. Appl. Microbiol. 2010, 109, 2214–2222. [Google Scholar] [CrossRef] [PubMed]
- Merkel, V.; Ohder, B.; Bielaszewska, M.; Zhang, W.; Fruth, A.; Menge, C.; Borrmann, E.; Middendorf, B.; Müthing, J.; Karch, H.; et al. Distribution and phylogeny of immunoglobulin-binding protein G in Shiga toxin-producing Escherichia coli and its association with adherence phenotypes. Infect. Immun. 2010, 78, 3625–3636. [Google Scholar] [CrossRef] [PubMed]
- Kuczius, T.; Zhang, W.; Merkel, V.; Mellmann, A.; Tarr, P.I.; Karch, H. Agitation down-regulates immunoglobulin binding protein EibG expression in Shiga toxin-producing Escherichia coli (STEC). PLoS ONE 2015, 10, e0119583. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 4, 822–880. [Google Scholar] [CrossRef] [PubMed]
- Karch, H.; Meyer, T. Evaluation of oligonucleotide probes for identification of Shiga-like-toxin-producing Escherichia coli. J. Clin. Microbiol. 1989, 27, 1180–1186. [Google Scholar] [PubMed]
- Bauwens, A.; Betz, J.; Meisen, I.; Kemper, B.; Karch, H.; Müthing, J. Facing glycosphingolipid–Shiga toxin interaction: Dire straits for endothelial cells of the human vasculature. Cell. Mol. Life Sci. 2013, 70, 425–457. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Koo, S.; Jeong, D.G.; Tesh, V.L. Shiga toxins as multi-functional proteins: Induction of host cellular stress responses, role in pathogenesis and therapeutic applications. Toxins 2016, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Betz, J.; Dorn, I.; Kouzel, I.U.; Bauwens, A.; Meisen, I.; Kemper, B.; Bielaszewska, M.; Mormann, M.; Weymann, L.; Sibrowski, W.; et al. Shiga toxin of enterohaemorrhagic Escherichia coli directly injures developing human erythrocytes. Cell. Microbiol. 2016, 10, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Sinha, B.; Kuczius, T.; Karch, H. Cytolethal distending toxin from Shiga toxin-producing Escherichia coli O157 causes irreversible G2/M arrest, inhibition of proliferation, and death of human endothelial cells. Infect. Immun. 2005, 73, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Stoewe, F.; Fruth, A.; Zhang, W.; Prager, R.; Brockmeyer, J.; Mellmann, A.; Karch, H.; Friedrich, A.W. Shiga toxin, cytolethal distending toxin, and hemolysin repertoires in clinical Escherichia coli O91 isolates. J. Clin. Microbiol. 2009, 47, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Karch, H.; Tarr, P.I.; Bielaszewska, M. Enterohaemorrhagic Escherichia coli in human medicine. Int. J. Med. Microbiol. 2005, 295, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Brunder, W.; Schmidt, H.; Karch, H. KatP, a novel catalase-peroxidase encoded by the large plasmid of enterohaemorrhagic Escherichia coli O157:H7. Microbiology 1996, 142, 3305–3315. [Google Scholar] [CrossRef] [PubMed]
- Brunder, W.; Schmidt, H.; Karch, H. EspP, a novel extracellular serine protease of enterohaemorrhagic Escherichia coli O157:H7 cleaves human coagulation factor V. Mol. Microbiol. 1997, 24, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Brockmeyer, J.; Spelten, S.; Kuczius, T.; Bielaszewska, M.; Karch, H. Structure and function relationship of the autotransport and proteolytic activity of EspP from Shiga toxin-producing Escherichia coli. PLoS ONE 2009, 4, e6100. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Aldick, T.; Bauwens, A.; Karch, H. Hemolysin of enterohemorrhagic Escherichia coli: Structure, transport, biological activity and putative role in virulence. Int. J. Med. Microbiol. 2014, 304, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Rüter, C.; Kunsmann, L.; Greune, L.; Bauwens, A.; Zhang, W.; Kuczius, T.; Kim, K.S.; Mellmann, A.; Schmidt, M.A.; et al. Enterohemorrhagic Escherichia coli hemolysin employs outer membrane vesicles to target mitochondria and cause endothelial and epithelial apoptosis. PLoS Pathog. 2013, 9, e1003797. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Iyoda, S.; Satou, H.; Satou, H.; Itoh, K.; Saitoh, T.; Watanabe, H. A new immunoglobulin-binding protein, EibG, is responsible for the chain-like adhesion phenotype of locus of enterocyte effacement-negative, Shiga toxin-producing Escherichia coli. Infect. Immun. 2006, 74, 5747–5755. [Google Scholar] [CrossRef] [PubMed]
- Werber, D.; Beutin, L.; Pichner, R.; Stark, K.; Fruth, A. Shiga toxin-producing Escherichia coli-serogroups in food and patients, Germany. Emerg. Infect. Dis. 2008, 14, 1803–1806. [Google Scholar] [CrossRef] [PubMed]
- Kendall, M.M.; Gruber, C.C.; Parker, C.T.; Sperandio, V. Ethanolamine controls expression of genes encoding components involved in interkingdom signaling and virulence in enterohemorrhagic Escherichia coli O157:H7. MBio 2012, 3, e00050-12. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Milflores-Flores, L.; Garcia-Gallegos, J.G.; Patel, S.D.; Best, A.; La Ragione, R.M.; Martinez-Laguna, Y.; Woodward, M.J. Environmental regulation and colonization attributes of the long polar fimbriae (LPF) of Escherichia coli O157:H7. Int. J. Med. Microbiol. 2007, 297, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Leo, J.C.; Lyskowski, A.; Hattula, K.; Hartmann, M.D.; Schwarz, H.; Butcher, S.J.; Linke, D.; Lupas, A.N.; Goldman, A. The structure of E. coli IgG-binding protein D suggests a general model for bending and binding in trimeric autotransporter adhesins. Structure 2011, 19, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Munn, C.B.; Ishiguro, E.E.; Kay, W.W.; Trust, T.J. Role of surface components in serum resistance of virulent Aeromonas salmonicida. Infect. Immun. 1982, 36, 1069–1075. [Google Scholar] [PubMed]
- Raeder, R.; Boyle, M.D. Properties of IgG-binding proteins expressed by Streptococcus pyogenes isolates are predictive of invasive potential. J. Infect. Dis. 1996, 173, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Beachey, E.H. Bacterial adherence: Adhesin-receptor interactions mediating the attachment of bacteria to mucosal surface. J. Infect. Dis. 1981, 143, 325–345. [Google Scholar] [CrossRef] [PubMed]
- Sokurenko, E.V.; Courtney, H.S.; Abraham, S.N.; Klemm, P.; Hasty, D.L. Functional heterogeneity of type 1 fimbriae of Escherichia coli. Infect. Immun. 1992, 60, 4709–4719. [Google Scholar] [PubMed]
- Markwell, M.A.; Haas, S.M.; Bieber, L.L.; Tolbert, N.E. A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Kuczius, T.; Koch, R.; Keyvani, K.; Karch, H.; Grassi, J.; Groschup, M.H. Regional and phenotype heterogeneity of cellular prion proteins in the human brain. Eur. J. Neurosci. 2007, 25, 2649–2655. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain No. | Serotype a | eib Genes | eib Subtype |
|---|---|---|---|
| 4798/97-1 | O146:H21 | eibG | α |
| 7140/96 | O91:H− [H14] | eibG | α |
| 2875/96 | O91:H14 [H14] | eibG | α |
| 393/98 | O91:H− [H14] | eibG | α |
| 1809/00 | O91:H14 [H14] | eibG | α |
| 01/E243 | O91:H− [H14] | eibG | α |
| 6705/95 | OR:H14 [H14] | eibG | α |
| 99-02787 | OR:H10 | eibG | α |
| 0520/99 | Ont:H30 | eibG | γ |
| 3671/97 | O91:H− [H14] | eibC, eibG | α |
| 172/98 | OR:H− | eibC, eibG | α |
| 6561/95 | Ont:H− [H14] | eibC, eibG | α |
| ECOR9 | Ont:H− | eibA, eibC eibD, eibE | - |
| ECOR2 | Ont:H32 | eibF | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubin, D.; Zhang, W.; Karch, H.; Kuczius, T. Distinct Expression of Immunoglobulin-Binding Proteins in Shiga Toxin-Producing Escherichia coli Implicates High Protein Stability and a Characteristic Phenotype. Toxins 2017, 9, 153. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9050153
Rubin D, Zhang W, Karch H, Kuczius T. Distinct Expression of Immunoglobulin-Binding Proteins in Shiga Toxin-Producing Escherichia coli Implicates High Protein Stability and a Characteristic Phenotype. Toxins. 2017; 9(5):153. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9050153
Chicago/Turabian StyleRubin, Dennis, Wenlan Zhang, Helge Karch, and Thorsten Kuczius. 2017. "Distinct Expression of Immunoglobulin-Binding Proteins in Shiga Toxin-Producing Escherichia coli Implicates High Protein Stability and a Characteristic Phenotype" Toxins 9, no. 5: 153. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins9050153