Frequency of EBV LMP-1 Promoter and Coding Variations in Burkitt Lymphoma Samples in Africa and South America and Peripheral Blood in Uganda


Abstract
:1. Introduction
2. Results
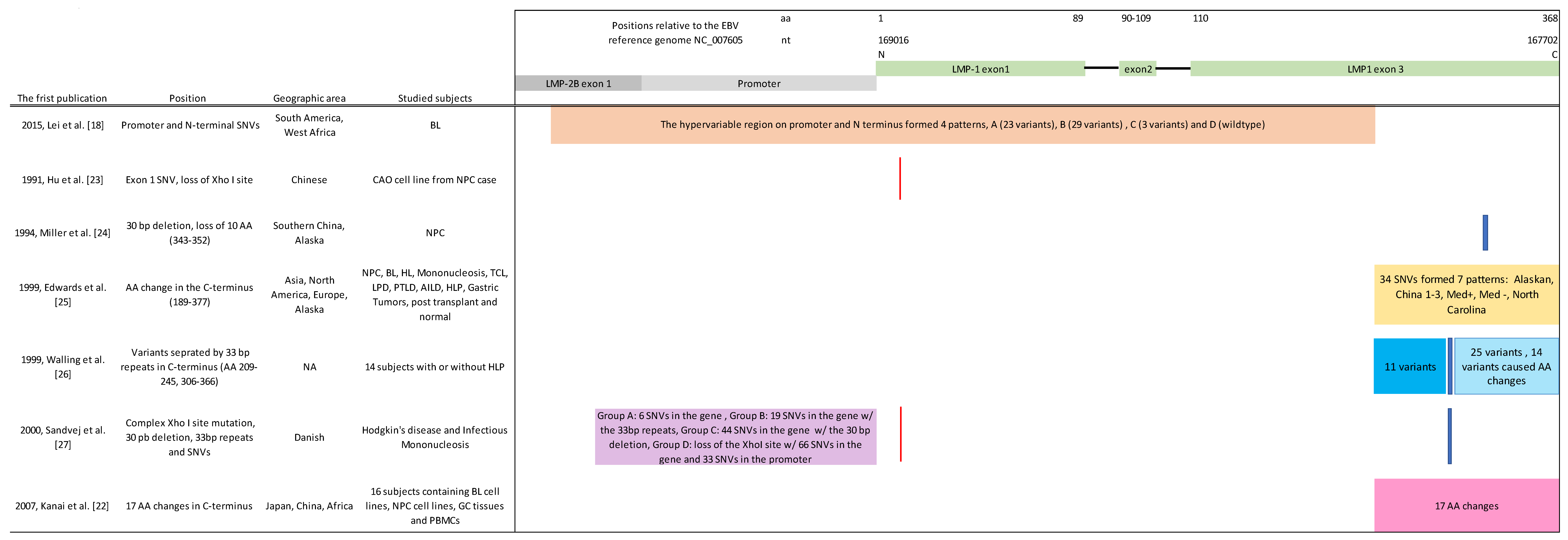
2.1. Comparison of EBV LMP-1 Genetic Diversity in 114 Published Genomes Classified by Seven Techniques
2.2. Discovery of Novel Pattern A-Like Variations in Samples Studied by PCR-Sanger Sequencing
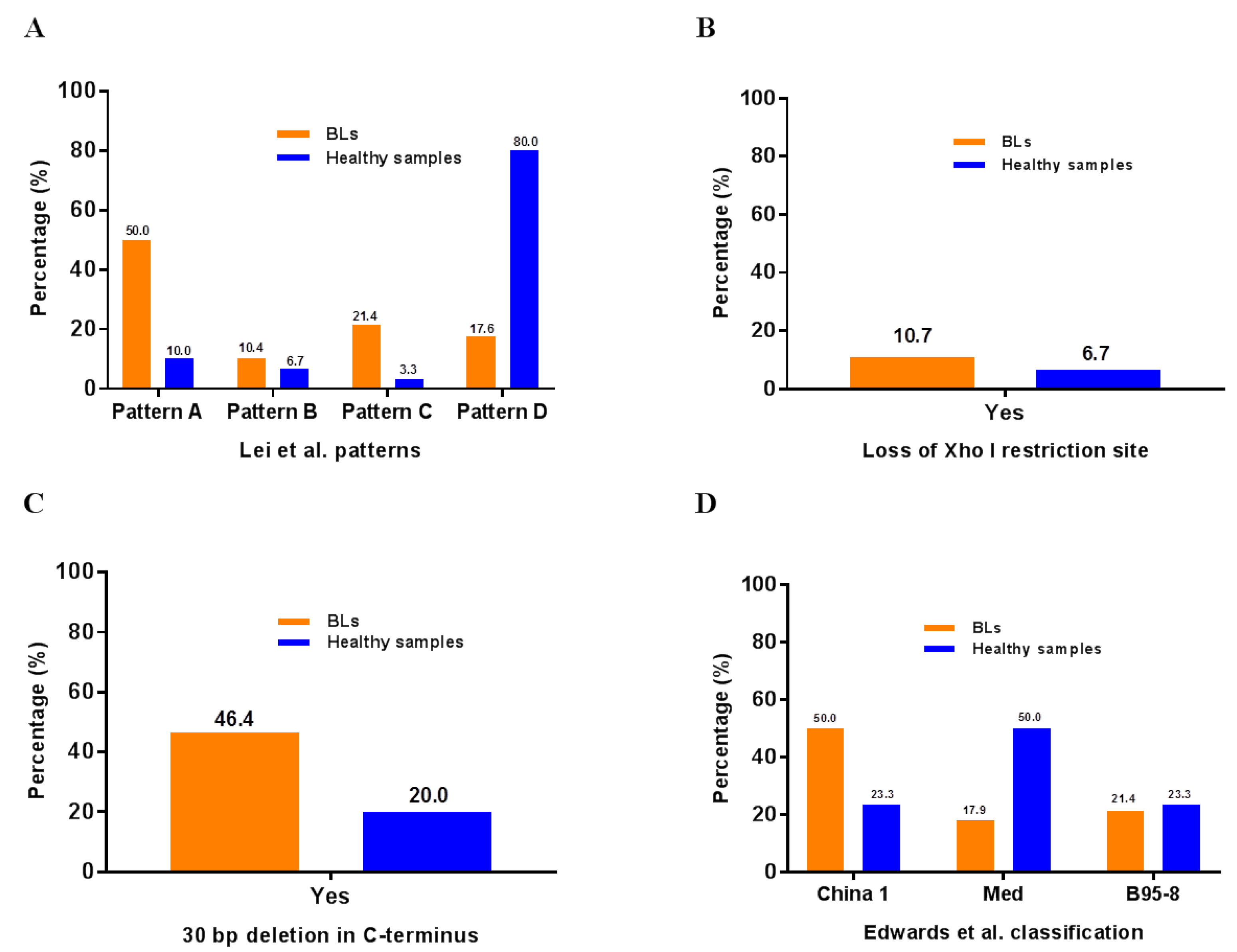
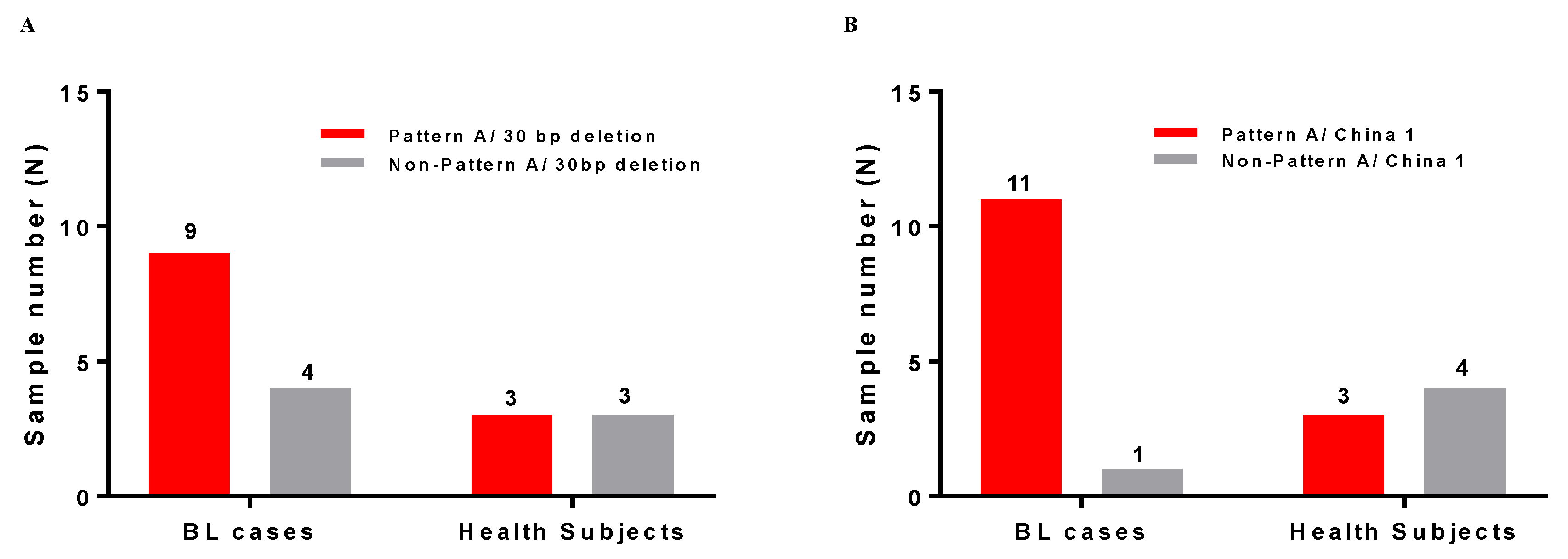
2.3. Frequency of Pattern A in 64 BL Tumors from Ghana, Argentina, and Brazil
2.4. Frequency of Pattern A in Peripheral Blood of 113 Children with- and 115 Children without-BL in Uganda
3. Discussion
3.1. LMP-1 Classification Using the Lei et al. Technique Applicable to HTS Data and Has Parsimony of Variants
3.2. Pattern A Variations Frequently Detected in BL Tumors or Peripheral Blood of eBL Cases
3.3. Novel Pattern A-Like Variations Suggest a Much Broader Undiscovered LMP-1 Genetic Diversity
3.4. Pattern A and D Heterozygosity in Tumors Suggests Multiple EBV Infections in Single Pre-Cancer B Cells
3.5. Implications of Detecting a High Frequency of Pattern A in BL Tumors or Peripheral Blood of eBL Cases
4. Materials and Methods
4.1. Subjects and IRB
4.2. DNA Extraction
4.3. PCR Amplification of the Targeted Hypervariable Region of LMP-1
4.4. Sanger Sequencing and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EBV | Epstein-Barr virus |
| LMP-1 | Latent membrane protein-1 |
| EBNA-1 | Epstein–Barr nuclear antigens-1 |
| BZLF-1 | Trans-activator protein BZLF1 |
| HTS | High throughput sequencing |
| SNV | Single nucleotide polymorphism |
| PCR | Polymerase chain reaction |
| BL | Burkitt lymphoma |
| eBL | endemic Burkitt lymphoma |
| NPC | Nasopharyngeal carcinoma |
| PTLD | Post-transplant lymphoproliferative disease |
| HL | Hodgkin Lymphoma |
| GC | EBV-positive gastric carcinoma |
| IM | Infectious mononucleosis |
| EMBLEM | Epidemiology of Burkitt’s Lymphoma in East-African Children and Minors |
| sLCL | Spontaneous transformed lymphoblastoid cell line |
| CTAR-1, 2, 3 | Carboxyl-terminal activator regions 1, -2, -3 |
| IL-6, -8, -10 | Interleukin-6, -8, -10 |
| SSCP | Single strand conformational polymorphism |
References
- Zur Hausen, H.; Schulte-Holthausen, H.; Klein, G.; Henle, W.; Henle, G.; Clifford, P.; Santesson, L. EBV DNA in biopsies of Burkitt tumours and anaplastic carcinomas of the nasopharynx. Nature 1970, 228, 1056–1058. [Google Scholar] [CrossRef] [PubMed]
- De Thé, G.; Ablashi, D.V.; Liabeuf, A.; Mourali, N. Nasopharyngeal carcinoma (NPC). VI. Presence of an EBV nuclear antigen in fresh tumour biopsies. Preliminary results. Biomedicine 1973, 19, 349–352. [Google Scholar] [PubMed]
- Proceedings of the IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Epstein-Barr Virus and Kaposi’s Sarcoma Herpesvirus/Human Herpesvirus 8. Lyon, France, 17–24 June 1997. IARC Monogr. Eval. Carcinog. Risks Hum. 1997, 70, 1–492.
- Harabuchi, Y.; Imai, S.; Wakashima, J.; Hirao, M.; Kataura, A.; Osato, T.; Kon, S. Nasal T-cell lymphoma causally associated with Epstein-Barr virus: Clinicopathologic, phenotypic, and genotypic studies. Cancer 1996, 77, 2137–2149. [Google Scholar] [CrossRef] [Green Version]
- Pittaluga, S.; Loke, S.L.; So, K.C.; Cheung, K.N.; Ma, L. Clonal Epstein-Barr virus in lymphoepithelioma-like carcinoma of the stomach: Demonstration of viral genome by in situ hybridization and southern blot analysis. Mod. Pathol. 1992, 5, 661–664. [Google Scholar] [PubMed]
- Shibata, D.; Hawes, D.; Stemmermann, G.N.; Weiss, L.M. Epstein-Barr virus-associated gastric adenocarcinoma among Japanese Americans in Hawaii. Cancer Epidemiol. Biomarkers Prev. 1993, 2, 213–217. [Google Scholar] [PubMed]
- Parkin, D.M.; Sitas, F.; Chirenje, M.; Stein, L.; Abratt, R.; Wabinga, H. Part I: Cancer in Indigenous Africans—burden, distribution, and trends. Lancet Oncol. 2008, 9, 683–692. [Google Scholar] [CrossRef]
- Thorley-Lawson, D.A. Epstein-Barr virus: Exploiting the immune system. Nat. Rev. Immunol. 2001, 1, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.; de Martel, C.; Vignat, J.; Ferlay, J.; Bray, F.; Franceschi, S. Global burden of cancers attributable to infections in 2012: A synthetic analysis. Lancet Glob. Health 2016, 4, e609–e616. [Google Scholar] [CrossRef]
- Chang, C.M.; Yu, K.J.; Mbulaiteye, S.M.; Hildesheim, A.; Bhatia, K. The extent of genetic diversity of Epstein-Barr virus and its geographic and disease patterns: A need for reappraisal. Virus Res. 2009, 143, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, J.L. Burkitt’s lymphoma. N. Engl. J. Med. 1981, 305, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, K.; Raj, A.; Guitierrez, M.I.; Judde, J.G.; Spangler, G.; Venkatesh, H.; Magrath, I.T. Variation in the sequence of Epstein Barr virus nuclear antigen 1 in normal peripheral blood lymphocytes and in Burkitt’s lymphomas. Oncogene 1996, 13, 177–181. [Google Scholar] [PubMed]
- Sample, J.; Young, L.; Martin, B.; Chatman, T.; Kieff, E.; Rickinson, A.; Kieff, E. Epstein-Barr virus types 1 and 2 differ in their EBNA-3A, EBNA-3B, and EBNA-3C genes. J. Virol. 1990, 64, 4084–4092. [Google Scholar] [PubMed]
- Tzellos, S.; Farrell, P.J. Epstein-Barr virus sequence variation-biology and disease. Pathogens 2012, 1, 156–174. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Palser, A.; Elgueta Karstegl, C.; Middeldorp, J.M.; Ramayanti, O.; Cohen, J.I.; Hildesheim, A.; Fellner, M.D.; Wiels, J.; White, R.E.; et al. Natural variation of Epstein-Barr virus genes, proteins, and primary microrna. J. Virol. 2017, 91, e00375-17. [Google Scholar] [CrossRef] [PubMed]
- Bornkamm, G.W. Epstein-Barr virus and the pathogenesis of Burkitt’s lymphoma: More questions than answers. Int. J. Cancer 2009, 124, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Kwok, H.; Chiang, A.K. From conventional to next generation sequencing of Epstein-Barr virus genomes. Viruses 2016, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Li, T.; Li, B.; Tsai, S.; Biggar, R.J.; Nkrumah, F.; Neequaye, J.; Gutierrez, M.; Epelman, S.; Mbulaiteye, S.M.; et al. Epstein-Barr virus from Burkitt lymphoma biopsies from Africa and South America share novel LMP-1 promoter and gene variations. Sci. Rep. 2015, 5, 16706. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I.; Fauci, A.S.; Varmus, H.; Nabel, G.J. Epstein-Barr virus: An important vaccine target for cancer prevention. Sci. Transl. Med. 2011, 3, 107fs7. [Google Scholar] [CrossRef] [PubMed]
- Legason, I.D.; Pfeiffer, R.M.; Udquim, K.I.; Bergen, A.W.; Gouveia, M.H.; Kirimunda, S.; Otim, I.; Karlins, E.; Kerchan, P.; Nabalende, H.; et al. Evaluating the causal link between malaria infection and endemic Burkitt lymphoma in Northern Uganda: A Mendelian Randomization study. EBio Medicine 2017, 25, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Maziarz, M.; Kinyera, T.; Otim, I.; Kagwa, P.; Nabalende, H.; Legason, I.D.; Ogwang, M.D.; Kirimunda, S.; Emmanuel, B.; Reynolds, S.J.; et al. Age and geographic patterns of plasmodium falciparum malaria infection in a representative sample of children living in Burkitt lymphoma-endemic areas of Northern Uganda. Malar. J. 2017, 16, 124. [Google Scholar] [CrossRef] [PubMed]
- Kanai, K.; Satoh, Y.; Yamanaka, H.; Kawaguchi, A.; Horie, K.; Sugata, K.; Hoshikawa, Y.; Sata, T.; Sairenji, T. The vIL-10 gene of the Epstein-Barr virus (EBV) is conserved in a stable manner except for a few point mutations in various EBV isolates. Virus Genes 2007, 35, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.F.; Zabarovsky, E.R.; Chen, F.; Cao, S.L.; Ernberg, I.; Klein, G.; Winberg, G. Isolation and sequencing of the Epstein-Barr virus BNLF-1 gene (LMP1) from a Chinese nasopharyngeal carcinoma. J. Gen. Virol. 1991, 72 Pt 10, 2399–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.E.; Edwards, R.H.; Walling, D.M.; Raab-Traub, N. Sequence variation in the Epstein-Barr virus latent membrane protein 1. J. Gen. Virol. 1994, 75 Pt 10, 2729–2740. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.H.; Seillier-Moiseiwitsch, F.; Raab-Traub, N. Signature amino acid changes in latent membrane protein 1 distinguish Epstein-Barr virus strains. Virology 1999, 261, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Walling, D.M.; Shebib, N.; Weaver, S.C.; Nichols, C.M.; Flaitz, C.M.; Webster-Cyriaque, J. The molecular epidemiology and evolution of Epstein-Barr virus: Sequence variation and genetic recombination in the latent membrane protein-1 gene. J. Infect. Dis. 1999, 179, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Sandvej, K.; Andresen, B.S.; Zhou, X.G.; Gregersen, N.; Hamilton-Dutoit, S. Analysis of the Epstein-Barr virus (EBV) latent membrane protein 1 (LMP-1) gene and promoter in hodgkin’s disease isolates: Selection against EBV variants with mutations in the LMP-1 promoter ATF-1/CREB-1 binding site. Mol. Pathol. 2000, 53, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.I.; Bhatia, K.; Cherney, B.; Capello, D.; Gaidano, G.; Magrath, I. Intraclonal molecular heterogeneity suggests a hierarchy of pathogenetic events in Burkitt’s lymphoma. Ann. Oncol. 1997, 8, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Mbulaiteye, S.M.; Walters, M.; Engels, E.A.; Bakaki, P.M.; Ndugwa, C.M.; Owor, A.M.; Goedert, J.J.; Whitby, D.; Biggar, R.J. High levels of Epstein-Barr virus DNA in saliva and peripheral blood from Ugandan mother-child pairs. J. Infect. Dis. 2006, 193, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Biggar, R.J.; Henle, W.; Fleisher, G.; Bocker, J.; Lennette, E.T.; Henle, G. Primary Epstein-Barr virus infections in African infants. I. Decline of maternal antibodies and time of infection. Int. J. Cancer 1978, 22, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Matrajt, L.; Gantt, S.; Mayer, B.T.; Krantz, E.M.; Orem, J.; Wald, A.; Corey, L.; Schiffer, J.T.; Casper, C. Virus and host-specific differences in oral human herpesvirus shedding kinetics among Ugandan women and children. Sci. Rep. 2017, 7, 13105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, A.S.; Mueller, N.E. Viruses and cancer. Causal associations. Ann. Epidemiol. 1990, 1, 71–92. [Google Scholar] [CrossRef]
- Munoz, N.; Bosch, F.X.; de Sanjose, S.; Herrero, R.; Castellsague, X.; Shah, K.V.; Snijders, P.J.; Meijer, C.J.; International Agency for Research on Cancer Multicenter Cervical Cancer Study Group. Epidemiologic classification of human papillomavirus types associated with cervical cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.M.; Syed, N.; Whittle, H.; Crawford, D.H. Circulating Epstein-Barr virus-carrying B cells in acute malaria. Lancet 1991, 337, 876–878. [Google Scholar] [CrossRef]
- Smatti, M.K.; Yassine, H.M.; AbuOdeh, R.; AlMarawani, A.; Taleb, S.A.; Althani, A.A.; Nasrallah, G.K. Prevalence and molecular profiling of Epstein Barr virus (EBV) among healthy blood donors from different nationalities in Qatar. PLoS ONE 2017, 12, e0189033. [Google Scholar] [CrossRef] [PubMed]
- Gantuz, M.; Lorenzetti, M.A.; Chabay, P.A.; Preciado, M.V. A novel recombinant variant of latent membrane protein 1 from Epstein Barr virus in Argentina denotes phylogeographical association. PLoS ONE 2017, 12, e0174221. [Google Scholar] [CrossRef] [PubMed]
- Kehdy, F.S.; Gouveia, M.H.; Machado, M.; Magalhaes, W.C.; Horimoto, A.R.; Horta, B.L.; Moreira, R.G.; Leal, T.P.; Scliar, M.O.; Soares-Souza, G.B.; et al. Origin and dynamics of admixture in Brazilians and its effect on the pattern of deleterious mutations. Proc. Natl. Acad. Sci. USA 2015, 112, 8696–8701. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.Y.; Tierney, R.J.; Croom-Carter, D.; Dukers, D.; Cooper, G.M.; Ellis, C.J.; Rowe, M.; Rickinson, A.B. Frequency of multiple Epstein-Barr virus infections in T-cell-immunocompromised individuals. J. Virol. 1996, 70, 4884–4894. [Google Scholar] [PubMed]
- Walling, D.M.; Brown, A.L.; Etienne, W.; Keitel, W.A.; Ling, P.D. Multiple Epstein-Barr virus infections in healthy individuals. J. Virol. 2003, 77, 6546–6550. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.I.; Spangler, G.; Kingma, D.; Raffeld, M.; Guerrero, I.; Misad, O.; Jaffe, E.S.; Magrath, I.T.; Bhatia, K. Epstein-Barr virus in nasal lymphomas contains multiple ongoing mutations in the EBNA-1 gene. Blood 1998, 92, 600–606. [Google Scholar] [PubMed]
- Thorley-Lawson, D.A.; Gross, A. Persistence of the Epstein-Barr virus and the origins of associated lymphomas. N. Engl. J. Med. 2004, 350, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Rastelli, J.; Homig-Holzel, C.; Seagal, J.; Muller, W.; Hermann, A.C.; Rajewsky, K.; Zimber-Strobl, U. LMP1 signaling can replace CD40 signaling in B cells in vivo and has unique features of inducing class-switch recombination to IgG1. Blood 2008, 111, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Uchida, J.; Yasui, T.; Takaoka-Shichijo, Y.; Muraoka, M.; Kulwichit, W.; Raab-Traub, N.; Kikutani, H. Mimicry of CD40 signals by Epstein-Barr virus LMP1 in B lymphocyte responses. Science 1999, 286, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Kaye, K.M.; Izumi, K.M.; Kieff, E. Epstein-Barr virus latent membrane protein 1 is essential for B-lymphocyte growth transformation. Proc. Natl. Acad. Sci. USA 1993, 90, 9150–9154. [Google Scholar] [CrossRef] [PubMed]
- Izumi, K.M.; Kaye, K.M.; Kieff, E.D. The Epstein-Barr virus lmp1 amino acid sequence that engages tumor necrosis factor receptor associated factors is critical for primary B lymphocyte growth transformation. Proc. Natl. Acad. Sci. USA 1997, 94, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Izumi, K.M.; Kieff, E.D. The Epstein-Barr virus oncogene product latent membrane protein 1 engages the tumor necrosis factor receptor-associated death domain protein to mediate B lymphocyte growth transformation and activate NF-kappaB. Proc. Natl. Acad. Sci. USA 1997, 94, 12592–12597. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.C.; Lin, S.J.; Lin, C.J.; Chou, Y.C.; Lin, J.H.; Yeh, T.H.; Chen, M.R.; Huang, L.M.; Lu, M.Y.; Huang, Y.C.; et al. Autocrine CCL3 and CCL4 induced by the oncoprotein LMP1 promote Epstein-Barr virus-triggered B cell proliferation. J. Virol. 2013, 87, 9041–9052. [Google Scholar] [CrossRef] [PubMed]
- Pratt, Z.L.; Zhang, J.; Sugden, B. The latent membrane protein 1 (LMP1) oncogene of Epstein-Barr virus can simultaneously induce and inhibit apoptosis in B cells. J. Virol. 2012, 86, 4380–4393. [Google Scholar] [CrossRef] [PubMed]
- Middeldorp, J.M.; Pegtel, D.M. Multiple roles of LMP1 in Epstein-Barr virus induced immune escape. Semin. Cancer Biol. 2008, 18, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.I.; Bhatia, K.; Barriga, F.; Diez, B.; Muriel, F.S.; de Andreas, M.L.; Epelman, S.; Risueno, C.; Magrath, I.T. Molecular epidemiology of Burkitt’s lymphoma from South America: Differences in breakpoint location and Epstein-Barr virus association from tumors in other world regions. Blood 1992, 79, 3261–3266. [Google Scholar] [PubMed]
- Nkrumah, F.K. Changes in the presentation of Burkitt’s lymphoma in Ghana over a 15-year period (1969–1982). IARC Sci. Publ. 1984, 665–674. [Google Scholar]
- Nkrumah, F.; Henle, W.; Henle, G.; Herberman, R.; Perkins, V.; Depue, R. Burkitt’s lymphoma: Its clinical course in relation to immunologic reactivities to Epstein-Barr virus and tumor-related antigens. J. Natl. Cancer Inst. 1976, 57, 1051–1056. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | LMP-2B Exon1 | LMP-1 Promoter | LMP-1 Exon1 | LMP-1 Exon3 | |||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| nt position | 1. −426 | 2. −412 | 3. −410 | 4. −367 | −372 | −356 | 5. −354 | −329 | −328 | −315 | −286 | −284 | −240 | −238 | −234 | −233 | 6. −227 | −207 | −199 | 7. −184 | 8. −172 | −163 | −136 | −70 | 9. −50 | −44 | −43 | −37 | −34 | 10. −39 | −17 | 11. −12 | −6 | −4 | −3 | +11 | +12 | 12. +18 | +26 | +32 | +41 | 13. E2D | 14.15. H3L | R17L | 16. V43I | 17. S57A | 18.19. I63V | 20. I124V | 21. I152L | 22. H213N | 23. E214Q |
| nt change | G>A | T>G | C>A | G>A | G>A | C>A | A>G | C>T | G>A | C>T | A>G | G>T | G>A | A>G | G>T | G>A | G>A | C>T | C>T | A>T | T>C | C>T | A>T | A>C | T>A | G>T | A>T | C>G | C>G | A>C | A>T | G>A | C>T | C>A | C>G | A>G | C>T | T>G | A>C | G>A | G>C | GAA>GAC | CAC>CTG | CGA>CTA | GTT>ATT | TCC>GCC | ATA>GTG | ATC>GTC | ATC>CTC | CAT>AAT | GAA>CAA |
| Pattern A (23 variants) | A | G | A | A | G | C | G | C | G | C | A | G | G | A | G | C | A | C | C | T | C | C | A | A | A | C | A | C | C | C | A | A | C | C | C | A | C | G | A | G | C | C | TG | G | A | G | GG | G | C | A | C |
| Pattern B (29 variants) | G | T | C | G | A | A | A | T | A | T | G | T | A | G | T | A | G | T | T | A | T | T | T | C | T | C | T | G | G | A | T | G | T | A | G | G | T | T | C | A | G | A | AC | T | G | T | AA | A | A | C | G |
| Pattern C (3 variants) | G | T | C | G | G | C | A | C | G | C | A | G | G | A | G | G | G | C | C | A | T | C | A | A | T | T | A | C | C | A | A | G | C | C | C | A | C | T | A | G | C | C | AC | G | G | T | AA | A | A | C | G |
| Pattern D (wild-type) | G | T | C | G | G | C | A | C | G | C | A | G | G | A | G | G | G | C | C | A | T | C | A | A | T | G | A | C | C | A | A | G | C | C | C | A | C | T | A | G | G | A | AC | G | G | T | AA | A | A | C | G |
| EBV Genome | Lei et al. [18] Patterns | Source | Origin | EBV Type | Hu et al. [23] Loss of Xho I Site | Miller et al. [24] 30 bp Deletion | 33 bp Repeats (# of RP Unit) | † Edwards et al. [25] C-Terminal Variants | # Sandvej et al. [27] Group |
|---|---|---|---|---|---|---|---|---|---|
| KP968263**(H058015C) | A | BL biopsy | Ghana | Type 1 | No | Yes | 4 | China 1 | ND |
| KP968262**(H018436D) | A | BL biopsy | Ghana | Type 1 | No | Yes | 4 | China 1 | ND |
| KP968264**(H002213) | A | BL biopsy | Ghana | Type 1 | No | Yes | 4 | China 1 | ND |
| KP968261**(HU11393) | A | BL biopsy | Ghana | Type 1 | No | Yes | 4 | ND | ND |
| KR063342**(H03753A) | A | BL biopsy | Ghana | Type 1 | No | No | 4 | China 1 | ND |
| KR063345**(FNR) | A | BL biopsy | Brazil | Type 1 | No | Yes | 4 | China 1 | ND |
| KR063344**(RPF) | A | BL biopsy | Brazil | Type 1 | No | Yes | 4 | China 1 | ND |
| KP968258**(MP) | A | BL biopsy | Brazil | Type 1 | No | Yes | 4 | China 1 | ND |
| KP968257**(CCH) | A | BL biopsy | Brazil | Type 1 | No | Yes | 4 | China 1 | ND |
| KR063343**(CV-ARG) | A | BL biopsy | Argentina | Type 1 | No | Yes | 4 | China 1 | ND |
| KT001102 (VA) | A | BL biopsy | Argentina | N/A | Gap | Gap | 4 | China 1 | ND |
| KT001103 (SG) | A | BL biopsy | Argentina | Type 1 | No | Gap | 4 | China 1 | ND |
| KP968259**(SCL) | A | BL biopsy | Brazil | Type 1 | No | No | 4 | B95–8 | ND |
| LN827554(LCL-AFB1) | A | LCL | Unknown | Type 2 | No | Yes | 5 | China 1 | ND |
| LN824206(pLCL-TRL1-post) | A | sLCL. PTLD (post) | USA | Type 1 | No | Yes | 5 | China 1 | ND |
| LN824207(pLCL-TRL1-pre) | A | sLCL. PTLD (pre) | USA | Type 1 | No | Yes | 5 | China 1 | ND |
| LN827591(sLCL-2.15) | A | sLCL | Kenya | Type 2 | No | Yes | 5 | China 1 | ND |
| LN827594(sLCL-IS1.07) | A | sLCL. PTLD | Australia | Type 1 | No | Yes | 5 | China 1 | ND |
| LN827559(pLCL-TRL595) | A | sLCL. PTLD | USA | Type 1 | No | Yes | 5 | China 1 | ND |
| LN827563(sLCL-1.18) | A | sLCL | Kenya | Type 1 | No | Yes | 5 | China 1 | ND |
| KF717093**(Raji) | A | BL | Nigeria | Type 1 | No | No | 4 | B95-8 | ND |
| KP968260**(VGO) | B | BL biopsy | Brazil | Type 1 | Yes | No | 5 | ND | ND |
| KC207813**(Akata) | B | BL | Japan | Type 1 | Yes | Yes | 4 | China 1 | ND |
| LN824208**(Akata) | B | BL | Japan | Type 1 | Yes | Yes | 4 | China 1 | ND |
| KC617875(C666-1) | B | NPC | Asia | Type 1 | Yes | Yes | 4 | China 1 | ND |
| KJ411974(C666-1) | B | NPC | Asia | Type 1 | Yes | Yes | 1 | China 1 | ND |
| KC617875(C666-1) | B | NPC | Asia | Type 1 | Yes | Yes | 4 | China 1 | ND |
| AY961628(GD1) | B | NPC | China | Type 1 | Yes | Yes | 4 | China 1 | ND |
| HQ020558(GD2) | B | NPC | China | Type 1 | Yes | Yes | 3 | China 1 | ND |
| KF373730(M81) | B | NPC | Asia | Type 1 | Yes | Yes | 4 | China 1 | ND |
| LN824142(Saliva) | B | Healthy saliva | UK | Type 1 | Yes | Yes | 2 | China 1 | ND |
| LN827562(sLCL-1.19) | B | sLCL | Kenya | Type 1 | Yes | No | 4 | ND | ND |
| LN827561(YCCEL1) | B | GC cell line | South Korea | Type 1 | Yes | Yes | 5 | China 1 | ND |
| LN824209(HKN14) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 5 | China 1 | ND |
| LN827547(HKN15) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 4 | China 1 | ND |
| LN824224(HKN19) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 4 | China 1 | ND |
| LN827549(D3201.2) | B | NPC | China | Type 1 | Yes | Yes | 6 | China 1 | ND |
| JQ009376(HKNPC1) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 5 | China 1 | ND |
| KF992564(HKNPC2) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 1 | China 1 | ND |
| KF992565(HKNPC3) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 4 | China 1 | ND |
| KF992566(HKNPC4) | B | NPC | Hong Kong | Type 1 | Gap | Gap | Gap | Gap | ND |
| KF992567(HKNPC5) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 1 | China 1 | ND |
| KF992568(HKNPC6) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 1 | China 1 | ND |
| KF992569(HKNPC7) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 1 | China 1 | ND |
| KF992570(HKNPC8) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 2 | China 1 | ND |
| KF992571(HKNPC9) | B | NPC | Hong Kong | Type 1 | Yes | Yes | 1 | China 1 | ND |
| LN827523(L591) | B | HL cell line | Germany | Type 1 | Yes | No | 4 | NC | ND |
| LN827799(sLCL-IM1.16) | B | sLCL. IM. | Australia | Type 1 | Yes | No | 4 | NC | ND |
| LN827578(sLCL-IS1.13) | B | sLCL. PTLD | Australia | Type 1 | Yes | No | 4 | NC | ND |
| LN827586(sLCL-IS1.15) | B | sLCL. PTLD | Australia | Type 1 | Yes | No | 4 | NC | ND |
| LN827800**(Jijoye) | C | BL | Nigeria | Type 2 | No | No | 4 | B95-8 | ND |
| LN827548**(P3HR1_c16) | C | BL | Nigeria | Type 2 | No | No | 4 | B95-8 | ND |
| LN827557**(BL36) | C | BL | N. Africa | Type 1 | No | No | 4 | B95-8 | ND |
| LN827545**(Daudi) | C | BL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827551**(Makau) | C | BL | Kenya | Type 1 | No | No | 5 | Med | ND |
| LN824205(sLCL-1.12) | C | sLCL | Kenya | Type 1 | No | No | 5 | Med | ND |
| LN824203**(Mak1) | C | BL | Kenya | Type 1 | No | No | 5 | Med | ND |
| LN827544**(Wewak1) | D | BL | PNG | Type 2 | No | No | 4 | ND | ND |
| LN827556**(Cheptages) | D | BL | Kenya | Type 2 | No | No | 4 | B95-8 | ND |
| LN827526**(BL37) | D | BL | Africa | Type 1 | No | Yes | 6 | Med | ND |
| KC207814**(Mutu) | D | BL | Kenya | Type 1 | No | No | 4 | Med | ND |
| NC_009334**(AG876) | D | BL | Ghana | Type 2 | No | Yes | 4 | China 1 | ND |
| NC_007605 (WT-EBV) | D | B98-8 | USA | type 1 | No | No | 4 | B95-8 | ND |
| AJ507799(WT-EBV) | D | B98-8 | USA | Type 1 | No | No | 4 | B95-8 | ND |
| V01555(WT-EBV) | D | B95-8 | USA | Type 1 | No | No | 4 | B95-8 | ND |
| KC440851(K4123-Mi) | D | healthy donor | USA | Type 1 | No | No | 4 | Med | ND |
| KC440852(K4413-Mi) | D | healthy donor | USA | Type 1 | No | No | 4 | B95-8 | A |
| NA19114 | D | healthy donor | Yoruba | Type 1 | No | No | 4 | B95-8 | ND |
| NA19315 | D | healthy donor | Kenya | Type 1 | No | No | 4 | B95-8 | ND |
| NA19384 | D | healthy donor | Kenya | Type 1 | Gap | No | 4 | China 1 | ND |
| LN827739(LCL_B958) | D | LCL, B95-8 | USA | Type 1 | No | No | 4 | B95-8 | ND |
| LN827597(sLCL-IS1.04) | D | sLCL. PTLD | Australia | Type 1 | No | No | 5 | B95-8 | ND |
| LN827596(sLCL-IM1.02) | D | sLCL. IM. | Australia | Type 1 | No | Yes | 6 | China 1 | ND |
| LN827595(sLCL-IS1.03) | D | sLCL. PTLD | Australia | Type 1 | No | No | 5 | Med | ND |
| LN827593(sLCL-IS1.12) | D | sLCL. PTLD | Australia | Type 1 | No | No | 4 | B95-8 | A |
| LN827592(sLCL-IS1.10) | D | sLCL. PTLD | Australia | Type 1 | No | No | 5 | Med | ND |
| LN827590(sLCL-IM1.05) | D | sLCL. IM. | Australia | Type 1 | No | Yes | 6 | China 1 | ND |
| LN827589(sLCL-IS2.01) | D | sLCL. PTLD | Australia | Type 2 | No | Yes | 4 | China 1 | ND |
| LN827588(sLCL-IS1.19) | D | sLCL. PTLD | Australia | Type 1 | No | No | 5 | B95-8 | A |
| LN827587(sLCL-2.21) | D | sLCL | Kenya | Type 2 | No | Yes | 4 | China 1 | ND |
| LN827585(sLCL-1.04) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827584(sLCL-IS1.06) | D | sLCL. PTLD | Australia | Type 1 | No | No | 4 | Med | ND |
| LN827583(sLCL-IM1.17) | D | sLCL. IM. | Australia | Type 1 | No | No | 4 | B95-8 | ND |
| LN827582(sLCL-BL1.03) | D | sLCL | Kenya | Type 1 | No | No | 4 | B95-8 | ND |
| LN827581(sLCL-1.05) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827580(sLCL-2.16) | D | sLCL | Kenya | Type 2 | No | No | 5 | B95-8 | ND |
| LN827579(sLCL-1.13) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827577(sLCL-1.17) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | A |
| LN827576(sLCL-IS1.20) | D | sLCL. PTLD | Australia | Type 1 | No | Yes | 7 | China 1 | ND |
| LN827575(sLCL-IS1.14) | D | sLCL. PTLD | Australia | Type 1 | No | No | 4 | B95-8 | A |
| LN827574(sLCL-1.09) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | A |
| LN827573(sLCL-1.10) | D | sLCL | Kenya | Type 1 | No | No | 3 | Med | ND |
| LN827572(sLCL-IS1.18) | D | sLCL. PTLD | Australia | Type 1 | No | No | 4 | B95-8 | ND |
| LN827571(sLCL-BL1.20) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827570(sLCL-IS1.01) | D | sLCL. PTLD | Australia | Type 1 | No | Yes | 4 | China 1 | ND |
| LN827569(sLCL-IS1.11) | D | sLCL. PTLD | Australia | Type 1 | No | No | 5 | Med | ND |
| LN827568(sLCL-1.24) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827567(sLCL-IM1.09) | D | sLCL. IM. | Australia | Type 1 | No | No | 4 | B95-8 | ND |
| LN827566(sLCL-1.06) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827565(sLCL-1.07) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827564(HL04) | D | HL | UK | Type 1 | No | Yes | 3 | China 1 | ND |
| LN827560(sLCL-2.14) | D | sLCL | Kenya | Type 2 | No | Yes | 4 | China 1 | ND |
| LN827558(sLCL-1.02) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827555(X50-7) | D | LCL | USA | Type 1 | No | No | 4 | B95-8 | ND |
| LN827553(sLCL-IS1.08) | D | sLCL. PTLD | Australia | Type 1 | No | No | 5 | Med | ND |
| LN827552(sLCL-1.08) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827550(sLCL-1.11) | D | sLCL | Kenya | Type 1 | No | No | 4 | Med | ND |
| LN827546(HL02) | D | HL | UK | Type 1 | No | Yes | 3 | China 1 | ND |
| LN827527(M-ABA) | D | LCL, NPC virus | N. Africa | Type 1 | No | No | 4 | B95-8 | A |
| LN827524(HL11) | D | HL | UK | Type 1 | No | No | 5 | Med | ND |
| LN827522(HL09) | D | HL | UK | Type 1 | No | Yes | 6 | China 1 | ND |
| LN824226(HL01) | D | HL | UK | Type 1 | No | Yes | 3 | China 1 | ND |
| LN824225(HL08) | D | HL | UK | Type 1 | No | Yes | 5 | China 1 | ND |
| LN824204(HL05) | D | HL | UK | Type 1 | No | Yes | 5 | China 1 | ND |
| Gene Region | LMP-2B Exon1 | LMP-1 Promoter | LMP-1 Exon1 | LMP-1 Exon3 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Primer pair | Lei-F3/Lei-R3 | Lei-F1/Lei-R1 | Lei-F2/Lei-R2 | ||||||||||||||||||||||
| Amplicon size | 396 bp | 435 bp | 428 bp | ||||||||||||||||||||||
| FT and a.a. affected | AML1 | LBF2 | LBF4 | CREB | E2D | H3L | S24A (new) | V43I | S57A | I63V | I124V | I152L | H213N | E214Q | |||||||||||
| Variation | 1. G-426A | 2. T-412G | 3. C-410A | 4. G-376A | 5. A-354G | 6. G-227A | 7. A-184T | 8. T-172C | 9. T-50A | 10. A-39C | 11. G-12A | 12. T+18G | 13. GAA>GAC | 14. CAC>CTG | 15. CAC>CTG | TCC>GCC | 16. GTT>ATT | 17. TCC>GCC | 18. ATA>GTG | 19. ATA>GTG | 20. ATC>GTC | 21. ATC>CTC | 22. CAT>AAT | 23. GAA>CAA | |
| Pattern | Example | ||||||||||||||||||||||||
| A | BLS000073 | A | G | A | A | G | A | T | C | A | C | A | G | C | T | G | T | A | G | G | G | G | C | A | C |
| Mid-Length A | BLS000207 | G | T | C | G | A | G | A | T | T | A | A | G | C | T | G | T | A | G | G | G | G | C | A | C |
| 4nt sub-pattern A | BLS000292 | G | T | C | G | A | G | A | T | T | C | G | T | C | T | G | G | G | T | A | A | A | A | C | G |
| 3nt sub-pattern A | BLS000020 | G | T | C | G | A | G | A | T | T | A | G | T | C | T | G | G | G | T | A | A | A | A | C | G |
| Partial A sub-pattern | BLS001943 | A | G | A | A | G | A | T | C | A | C | A | G | C | T | G | T | G | T | A | A | A | A | C | C |
| Geographical Region | PCR Positive * | PCR Negative * | Pattern A | Mid Length-A | Pattern D | Pattern B | Pattern C | Pattern A/D |
|---|---|---|---|---|---|---|---|---|
| Argentina | 15 | 3 | 8 (66.7%) | 2 (16.7%) | 2 (16.7%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Brazil | 11 | 1 | 8 (80.0%) | 0 (0%) | 1 (10.0%) | 1 (10.0%) | 0 (0%) | 0 (0%) |
| Ghana | 45 | 3 | 23 (54.8%) | 5 (11.9%) | 11 (26.2%) | 0 (0%) | 1 (2.4%) | 2 (4.8%) |
| Total (N) | 71 | 7 | 39 (60.9%) | 7 (10.9%) | 14 (21.9%) | 1 (1.6%) | 1 (1.6%) | 2 (3.1%) |
| Sample Batch | Group | Number of PBMCs | PCR Positive * | PCR Negative | Pattern A | Mid-Length A sub-Pattern | Partial A sub-Pattern | 4nt sub-Pattern A | 3nt sub-Pattern A | Pattern D | N.D. # |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Batch 1 | BL cases | 13 | 13 (100%) | 0 (0%) | 2 (15.4%) | 0 (0%) | 0 (0%) | 1 (7.7%) | 1 (7.7%) | 9 (69.2%) | 0 (0%) |
| Controls | 15 | 15 (100%) | 0 (0%) | 0 (0%) | 4 (26.7%) | 0 (0%) | 4 (26.7%) | 0 (0%) | 7 (46.7%) | 0 (0%) | |
| Batch 2 | BL cases | 100 | 86 (86%) | 14 (14%) | 10 (10.0%) | 7 (7.0%) | 3 (3.5%) | 8 (9.3%) | 3 (3.5%) | 55 (55.0%) | 0 (0%) |
| Controls | 100 | 29 (86%) | 71 (71%) | 3 (3.0%) | 4(4.0%) | 0 (0%) | 0 (0%) | 0 (0%) | 19 (19.0%) | 3 (3.0%) | |
| Batch 1 and 2 | BL cases | 113 | 99 (87.6%) | 14 (12.4%) | 12 (10.6%) | 7 (6.2%) | 3 (2.6%) | 9 (8.0%) | 4 (3.5%) | 64 (56.4%) | 0 (0%) |
| Controls | 115 | 41 (35.6%) | 71 (64.4%) | 3 (2.6%) | 8 (7.0%) | 0 (0%) | 4 (3.5%) | 0 (0%) | 26 (22.6%) | 3 (2.6%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, H.-M.; Liu, H.; Lei, H.; Li, B.; Chin, P.-J.; Tsai, S.; Bhatia, K.; Gutierrez, M.; Epelman, S.; Biggar, R.J.; et al. Frequency of EBV LMP-1 Promoter and Coding Variations in Burkitt Lymphoma Samples in Africa and South America and Peripheral Blood in Uganda. Cancers 2018, 10, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10060177
Liao H-M, Liu H, Lei H, Li B, Chin P-J, Tsai S, Bhatia K, Gutierrez M, Epelman S, Biggar RJ, et al. Frequency of EBV LMP-1 Promoter and Coding Variations in Burkitt Lymphoma Samples in Africa and South America and Peripheral Blood in Uganda. Cancers. 2018; 10(6):177. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10060177
Chicago/Turabian StyleLiao, Hsiao-Mei, Hebing Liu, Heiyan Lei, Bingjie Li, Pei-Ju Chin, Shien Tsai, Kishor Bhatia, Marina Gutierrez, Sidnei Epelman, Robert J. Biggar, and et al. 2018. "Frequency of EBV LMP-1 Promoter and Coding Variations in Burkitt Lymphoma Samples in Africa and South America and Peripheral Blood in Uganda" Cancers 10, no. 6: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10060177