Immunosenescence and Immunotherapy in Elderly Acute Myeloid Leukemia Patients: Time for a Biology-Driven Approach

Abstract
:1. Introduction
2. Immunosenescence and Immunoaging (Biological Background)
3. Immunotherapy for Elderly Patients with AML: Different Strategies to Harness the Immune System against Leukemic Clones
3.1. Vaccines
3.2. Natural Killer Cell Therapy
4. Chimeric Antigen Receptor (CAR) T Cells
- (1)
- CD123, which is over-expressed in AML compared with normal hematopoietic cells [34,35]. CD123 expression is mainly restricted to cells of the myeloid lineage, is absent in T cells and shows limited expression on hematopoietic stem cells. Recently, a myeloablative CAR-based therapy targeting CD123 (CART123) has shown, in a mouse model, a potent effector activity against cell-line and primary AML [36]. Moreover, CART123 led to long-term survival of mice engrafted, and resulted in the establishment of a T-cell memory pool able to reject diseases [36]. Mardiros et al. [35] developed and evaluated 2 CARs containing a CD123-specific single-chain variable fragment, in combination with a CD28 costimulatory domain and CD3ζ-signaling domain, targeting different epitopes on CD123. These second generation CD123 CAR T cell activated T-cell effector functions against poor-risk primary AML patient samples. Additionally, T cells obtained from patients with active AML and genetically modified to express CAR 123 were able to lyse autologous AML blasts in vitro. Finally, a single injection of CD123 CAR T cells exhibited a significant antileukemic activity in vivo against a xenogeneic model of disseminated AML [35]. Currently, a trial with anti-CD123 CARs for relapsed or refractory AML patients was recently started and is actively recruiting patients (NCT02159495).
- (2)
- CD33. This differentiation antigen is predominantly expressed on myeloid cells, and it is also expressed in a subset of T cells, making it a non-ideal target for a CAR based therapy. A Chinese phase I clinical trial has studied the feasibility of anti-CD33 CAR in the treatment of relapsed or refractory AML. Only one patient was treated. Suggestions of a beneficial effect were present, but severe side effects, such as fever, cytokine release syndrome and pancytopenia, were reported [37].
- (3)
- LeY dicofusylated carbohydrate antigen). Recently, a small study tested the feasibility and the safety of CAR anti-LeY (therapy in patients with relapsed AML, in whom the blasts were shown to express LeY). The transducted and expanded autologous CAR T cells were successfully and safely infused in 4 patients with high-risk AML, showing tissue specific localization, long-term persistence and antileukemic efficacy [38].
5. Checkpoint Inhibitors
6. How Immunosenescence May Influence the Different Immunotherapy Strategies in AML Patients
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Lane, S.W.; Scadden, D.T.; Gilliland, D.G. The leukemic stem cell niche: Current concepts and therapeutic opportunities. Blood 2009, 114, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Döhner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Büchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.M.; Mandrekar, S.J.; Sanford, B.L.; Laumann, K.; Geyer, S.; Bloomfield, C.D.; Thiede, C.; Prior, T.W.; Döhner, K.; Marcucci, G.; et al. Midostaurin plus Chemotherapy for Acute Myeloid Leukemia with a FLT3 Mutation. N. Engl. J. Med. 2017, 377, 454–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute Myeloid Leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef] [PubMed]
- Isidori, A.; Venditti, A.; Maurillo, L.; Buccisano, F.; Loscocco, F.; Manduzio, P.; Sparaventi, G.; Amadori, S.; Visani, G. Alternative novel therapies for the treatment of elderly acute myeloid leukemia patients. Expert Rev. Hematol. 2013, 6, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Ayala, F.; Dewar, R.; Kieran, M.; Kalluri, R. Contribution of bone microenvironment to leukemogenesis and leukemia progression. Leukemia 2009, 23, 2233–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Gajewski, T.F.; Kline, J. PD-1/PD-L1 interactions inhibit antitumor immune responses in a murine acute myeloid leukemia model. Blood 2009, 114, 1545–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konopleva, M.Y.; Jordan, C.T. Leukemia stem cells and microenvironment: Biology and therapeutic targeting. J. Clin. Oncol. 2011, 29, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: Friends or Foes? Front. Immunol. 2018, 8, 1960. [Google Scholar] [CrossRef] [PubMed]
- Nikolich-Žugich, J. The twilight of immunity: Emerging concepts in aging of the immune system. Nat. Immunol. 2018, 19, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Meng, X.; Chen, Y.; Leng, S.X.; Zhang, H. The Biology of Aging and Cancer: Frailty, Inflammation, and Immunity. Cancer J. 2017, 23, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Salvioli, S.; Garagnani, P.; de Eguileor, M.; Monti, D.; Capri, M. Immunobiography and the Heterogeneity of Immune Responses in the Elderly: A Focus on Inflammaging and Trained Immunity. Front. Immunol. 2017, 8, 982. [Google Scholar] [CrossRef] [PubMed]
- Solana, R.; Tarazona, R.; Gayoso, I.; Lesur, O.; Dupuis, G.; Fulop, T. Innate immunosenescence: Effect of aging on cells and receptors of the innate immune system in humans. Semin. Immunol. 2012, 24, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Yanes, R.E.; Gustafson, C.E.; Weyand, C.M.; Goronzy, J.J. Lymphocyte generation and population homeostasis throughout life. Semin. Hematol. 2017, 54, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, R.; Smyth, M.J.; Lane, S.W. Harnessing the immune system in acute myeloid leukemia. Crit. Rev. Oncol. Hematol. 2016, 103, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Parisi, S.; Lecciso, M.; Ocadlikova, D.; Salvestrini, V.; Ciciarello, M.; Forte, D.; Corradi, G.; Cavo, M.; Curti, A. The More, The Better: “Do the Right Thing” For Natural Killer Immunotherapy in Acute Myeloid Leukemia. Front. Immunol. 2017, 19, 1330. [Google Scholar] [CrossRef] [PubMed]
- Geiger, T.L.; Rubnitz, J.E. New approaches for the immunotherapy of acute myeloid leukemia. Discov. Med. 2015, 19, 275–284. [Google Scholar] [PubMed]
- Isidori, A.; Loscocco, F.; Ciciarello, M.; Visani, G.; Corradi, G.; Forte, D.; Lecciso, M.; Ocadlikova, D.; Parisi, S.; Salvestrini, V.; et al. Renewing the immunological approach to AML treatment: From novel pathways to innovative therapies. Cancer Res. Front. 2016, 2, 226–251. [Google Scholar] [CrossRef]
- Greiner, J.; Döhner, H.; Schmitt, M. Cancer vaccines for patients with acute myeloid leukemia—Definition of leukemia-associated antigens and current clinical protocols targeting these antigens. Haematologica 2006, 91, 1653–1661. [Google Scholar] [PubMed]
- Scheibenbogen, C.; Letsch, A.; Thiel, E.; Schmittel, A.; Mailaender, V.; Baerwolf, S.; Nagorsen, D.; Keilholz, U. CD8 T-cell responses to Wilms tumor gene product WT1 and proteinase 3 in patients with acute myeloid leukemia. Blood 2002, 100, 2132–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, Y.; Tsuboi, A.; Taguchi, T.; Osaki, T.; Kyo, T.; Nakajima, H.; Elisseeva, O.A.; Oji, Y.; Kawakami, M.; Ikegame, K.; et al. Induction of WT1 (Wilms’ tumor gene)-specific cytotoxic T lymphocytes by WT1 peptide vaccine and the resultant cancer regression. Proc. Natl. Acad. Sci. USA 2004, 101, 13885–13890. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.; Schmitt, A.; Rojewski, M.T.; Chen, J.; Giannopoulos, K.; Fei, F.; Yu, Y.; Götz, M.; Heyduk, M.; Ritter, G.; et al. RHAMM-R3 peptide vaccination in patients with acute myeloid leukemia, myelodysplastic syndrome, and multiple myeloma elicits immunologic and clinical responses. Blood 2008, 111, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Anguille, S.; Van de Velde, A.L.; Smits, E.L.; Van Tendeloo, V.F.; Juliusson, G.; Cools, N.; Nijs, G.; Stein, B.; Lion, E.; Van Driessche, A.; et al. Dendritic cell vaccination as postremission treatment to prevent or delay relapse in acute myeloid leukemia. Blood 2017, 130, 1713–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, W. Infusions of allogeneic natural killer cells as cancer therapy. Clin. Cancer Res. 2014, 20, 3390–3400. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, L.; Capanni, M.; Urbani, E.; Perruccio, K.; Shlomchik, W.D.; Tosti, A.; Posati, S.; Rogaia, D.; Frassoni, F.; Aversa, F.; et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science 2002, 295, 2097–2100. [Google Scholar] [CrossRef] [PubMed]
- Curti, A.; Ruggeri, L.; D’Addio, A.; Bontadini, A.; Dan, E.; Motta, M.R.; Trabanelli, S.; Giudice, V.; Urbani, E.; Martinelli, G.; et al. Successful transfer of alloreactive haploidentical KIR ligand-mismatched natural killer cells after infusion in elderly high risk acute myeloid leukemia patients. Blood 2011, 118, 3273–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curti, A.; Ruggeri, L.; Parisi, S.; Bontadini, A.; Dan, E.; Motta, M.R.; Rizzi, S.; Trabanelli, S.; Ocadlikova, D.; Lecciso, M.; et al. Larger Size of Donor Alloreactive NK Cell Repertoire Correlates with Better Response to NK Cell Immunotherapy in Elderly Acute Myeloid Leukemia Patients. Clin. Cancer Res. 2016, 22, 1914–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisaki, H.; Kakuda, H.; Shimasaki, N.; Imai, C.; Ma, J.; Lockey, T.; Eldridge, P.; Leung, W.H.; Campana, D. Expansion of highly cytotoxic human natural killer cells for cancer cell therapy. Cancer Res. 2009, 69, 4010–4017. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.H.; Vong, Q.P.; Lin, W.; Janke, L.; Chen, T.; Leung, W. Modulation of NKG2D ligand expression and metastasis in tumors by spironolactone via RXRγ activation. J. Exp. Med. 2013, 210, 2675–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, W.K.; Kung Sutherland, M.; Li, Y.; Zalevsky, J.; Schell, S.; Leung, W. Antibody-dependent cell-mediated cytotoxicity overcomes NK cell resistance in MLL-rearranged leukemia expressing inhibitory KIR ligands but not activating ligands. Clin. Cancer Res. 2012, 18, 6296–6305. [Google Scholar] [CrossRef] [PubMed]
- Vey, N.; Bourhis, J.H.; Boissel, N.; Bordessoule, D.; Prebet, T.; Charbonnier, A.; Etienne, A.; Andre, P.; Romagne, F.; Benson, D.; et al. A phase 1 trial of the anti-inhibitory KIR mAb IPH2101 for AML in complete remission. Blood 2012, 120, 4317–4323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachanova, V.; Cooley, S.; Defor, T.E.; Verneris, M.R.; Zhang, B.; McKenna, D.H.; Curtsinger, J.; Panoskaltsis-Mortari, A.; Lewis, D.; Hippen, K.; et al. Clearance of acute myeloid leukemia by haploidentical natural killer cells is improved using IL-2 diphtheria toxin fusion protein. Blood 2014, 123, 3855–3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenegger, F.S.; Krupka, C.; Kohnke, T.; Subklewe, M. Immunotherapy for Acute Myeloid Leukemia. Semin. Hematol. 2015, 52, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.T.; Upchurch, D.; Szilvassy, S.J.; Guzman, M.L.; Howard, D.S.; Pettigrew, A.L.; Meyerrose, T.; Rossi, R.; Grimes, B.; Rizzieri, D.A.; et al. The interleukin-3 receptor alpha chain is a unique marker for human acute myelogenous leukemia stem cells. Leukemia 2000, 14, 1777–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardiros, A.; Dos Santos, C.; McDonald, T.; Brown, C.E.; Wang, X.; Budde, L.E.; Hoffman, L.; Aguilar, B.; Chang, W.C.; Bretzlaff, W.; et al. T cells expressing CD123-specific chimeric antigen receptors exhibit specific cytolytic effector functions and antitumor effects against human acute myeloid leukemia. Blood 2013, 122, 3138–3148. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Tasian, S.K.; Ruella, M.; Shestova, O.; Li, Y.; Porter, D.L.; Carroll, M.; Danet-Desnoyers, G.; Scholler, J.; Grupp, S.A.; et al. Preclinical targeting of human acute myeloid leukemia and myeloablation using chimeric antigen receptor-modified T cells. Blood 2014, 123, 2343–2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.S.; Wang, Y.; Lv, H.Y.; Han, Q.W.; Fan, H.; Guo, B.; Wang, L.L.; Han, W.D. Treatment of CD33-directed chimeric antigen receptor-modified T cells in one patient with relapsed and refractory acute myeloid leukemia. Mol. Ther. 2015, 23, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, D.S.; Neeson, P.J.; Khot, A.; Peinert, S.; Tai, T.; Tainton, K.; Chen, K.; Shin, M.; Wall, D.M.; Hönemann, D.; et al. Persistence and efficacy of second generation CAR T cell against the LeY antigen in acute myeloid leukemia. Mol. Ther. 2013, 21, 2122–2129. [Google Scholar] [CrossRef] [PubMed]
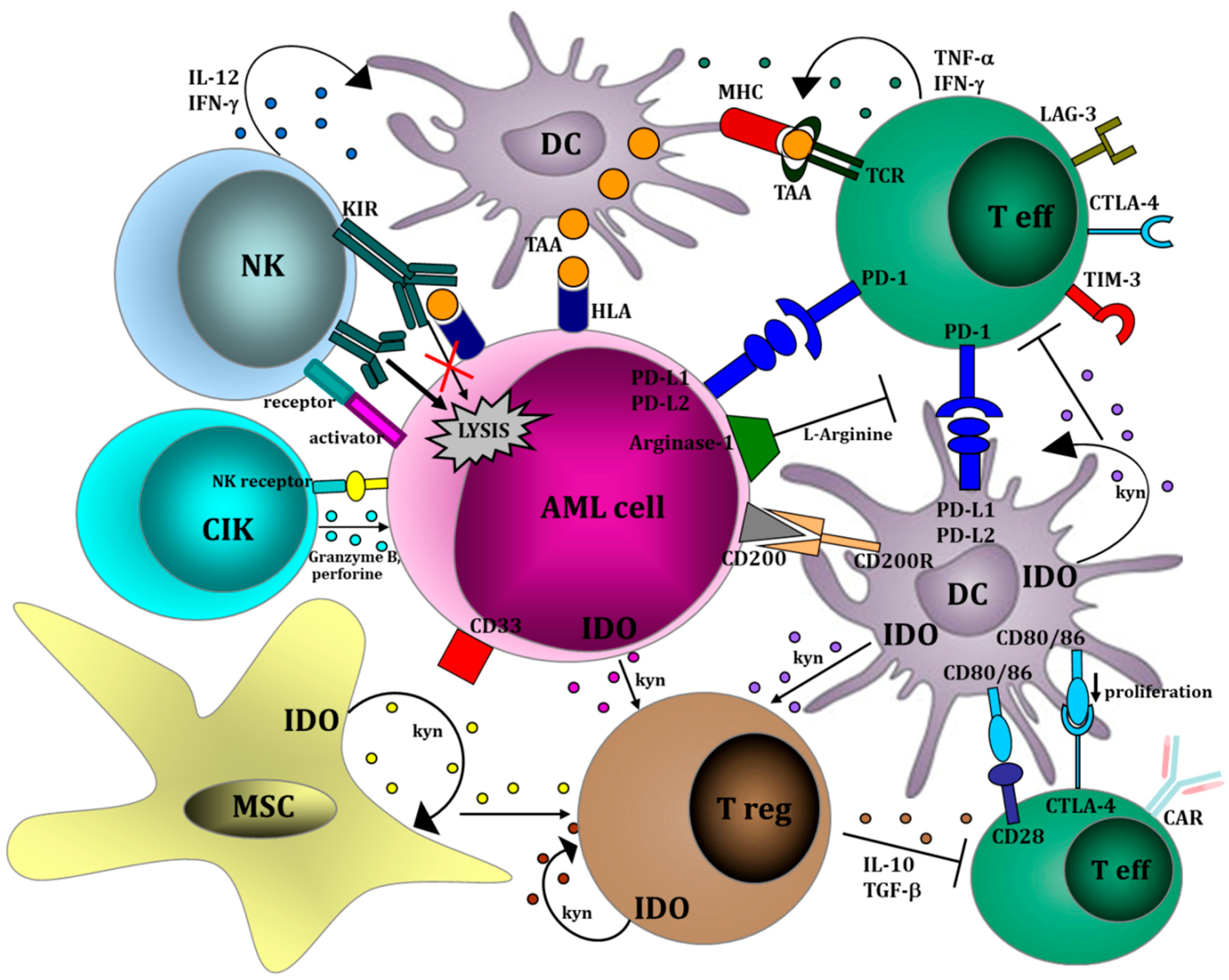
- Isidori, A.; Salvestrini, V.; Ciciarello, M.; Loscocco, F.; Visani, G.; Parisi, S.; Lecciso, M.; Ocadlikova, D.; Rossi, L.; Gabucci, E.; et al. The role of the immunosuppressive microenvironment in acute myeloid leukemia development and treatment. Expert Rev. Hematol. 2014, 7, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Thommen, D.S.; Schumacher, T.N. T Cell Dysfunction in Cancer. Cancer Cell 2018, 33, 547–562. [Google Scholar] [CrossRef] [PubMed]
- McKinney, E.F.; Smith, K.G.C. Metabolic exhaustion in infection, cancer and autoimmunity. Nat. Immunol. 2018, 19, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed]
- Francisco, L.M.; Sage, P.T.; Sharpe, A.H. The PD-1 pathway in tolerance and autoimmunity. Immunol. Rev. 2010, 236, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Bour-Jordan, H.; Esensten, J.H.; Martinez-Llordella, M.; Penaranda, C.; Stumpf, M.; Bluestone, J.A. Intrinsic and extrinsic control of peripheral T-cell tolerance by costimulatory molecules of the CD28/B7 family. Immunol. Rev. 2011, 241, 180–205. [Google Scholar] [CrossRef] [PubMed]
- Knaus, H.A.; Kanakry, C.G.; Luznik, L.; Gojo, I. Immunomodulatory Drugs: Immune Checkpoint Agents in Acute Leukemia. Curr. Drug Targets 2017, 18, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Bueso-Ramos, C.; DiNardo, C.; Estecio, M.R.; Davanlou, M.; Geng, Q.R.; Fang, Z.; Nguyen, M.; Pierce, S.; Wei, Y.; et al. Expression of PD-L1, PD-L2, PD-1 and CTLA4 in myelodysplastic syndromes is enhanced by treatment with hypomethylating agents. Leukemia 2014, 28, 1280–1288. [Google Scholar] [CrossRef] [PubMed]
- Ørskov, A.D.; Treppendahl, M.B.; Skovbo, A.; Holm, M.S.; Friis, L.S.; Hokland, M.; Grønbæk, K. Hypomethylation and up-regulation of PD-1 in T cells by azacytidine in MDS/AML patients: A rationale for combined targeting of PD-1 and DNA methylation. Oncotarget 2015, 6, 9612–9626. [Google Scholar] [CrossRef] [PubMed]
- Youngblood, B.; Oestreich, K.J.; Ha, S.J.; Duraiswamy, J.; Akondy, R.S.; West, E.E. Chronic virus infection enforces demethylation of the locus that encodes PD-1 in antigenspecific CD8(+) T cells. Immunity 2011, 35, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Berger, R.; Rotem-Yehudar, R.; Slama, G.; Landes, S.; Kneller, A.; Leiba, M.; Koren-Michowitz, M.; Shimoni, A.; Nagler, A. Phase I safety and pharmacokinetic study of CT-011, a humanized antibody interacting with PD-1, in patients with advanced hematologic malignancies. Clin. Cancer Res. 2008, 14, 3044–3051. [Google Scholar] [CrossRef] [PubMed]
- Daver, N.; Boddu, P.; Garcia-Manero, G.; Yadav, S.S.; Sharma, P.; Allison, J.; Kantarjian, H. Hypomethylating agents in combination with immune checkpoint inhibitors in acute myeloid leukemia and myelodysplastic syndromes. Leukemia 2018, 32, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Daver, N.G.; Basu, S.; Garcia-Manero, G.; Cortes, J.E.; Ravandi, F.; Jabbour, E.; Hendrickson, S.; Brandt, M.; Pierce, S.; Gordon, T.; et al. Phase IB/II study of nivolumab with azacytidine (AZA) in patients (PTS) with relapsed AML. J. Clin. Oncol. 2017, 35, 7026. [Google Scholar]
- Daste, A.; Domblides, C.; Gross-Goupil, M.; Chakiba, C.; Quivy, A.; Cochin, V.; de Mones, E.; Larmonier, N.; Soubeyran, P.; Ravaud, A. Immune checkpoint inhibitors and elderly people: A review. Eur. J. Cancer 2017, 82, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, R.; Mezquita, L.; Auclin, E.; Chaput, N.; Besse, B. Immunosenescence and immunecheckpoint inhibitors in non-small cell lung cancer patients: Does age really matter? Cancer Treat. Rev. 2017, 60, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Berthon, C.; Driss, V.; Liu, J.; Kuranda, K.; Leleu, X.; Jouy, N.; Hetuin, D.; Quesnel, B. In acute myeloid leukemia, B7-H1 (PD-L1) protection of blasts from cytotoxic T cells is induced by TLR ligands and interferon-gamma and can be reversed using MEK inhibitors. Cancer Immunol. Immunother. 2010, 59, 1839–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Munger, M.E.; Highfill, S.L.; Tolar, J.; Weigel, B.J.; Riddle, M.; Sharpe, A.H.; Vallera, D.A.; Azuma, M.; Levine, B.L.; et al. Program death-1 signaling and regulatory T cells collaborate to resist the function of adoptively transferred cytotoxic T lymphocytes in advanced acute myeloid leukemia. Blood 2010, 116, 2484–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Correa, B.; Campos, C.; Pera, A.; Bergua, J.M.; Arcos, M.J.; Bañas, H.; Casado, J.G.; Morgado, S.; Duran, E.; Solana, R.; Tarazona, R. Natural killer cell immunosenescence in acute myeloid leukaemia patients: New targets for immunotherapeutic strategies? Cancer Immunol. Immunother. 2016, 65, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Martner, A.; Rydström, A.; Riise, R.E.; Aurelius, J.; Brune, M.; Foà, R.; Hellstrand, K.; Thorén, F.B. NK cell expression of natural cytotoxicity receptors may determine relapse risk in older AML patients undergoing immunotherapy for remission maintenance. Oncotarget 2015, 6, 42569–42574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
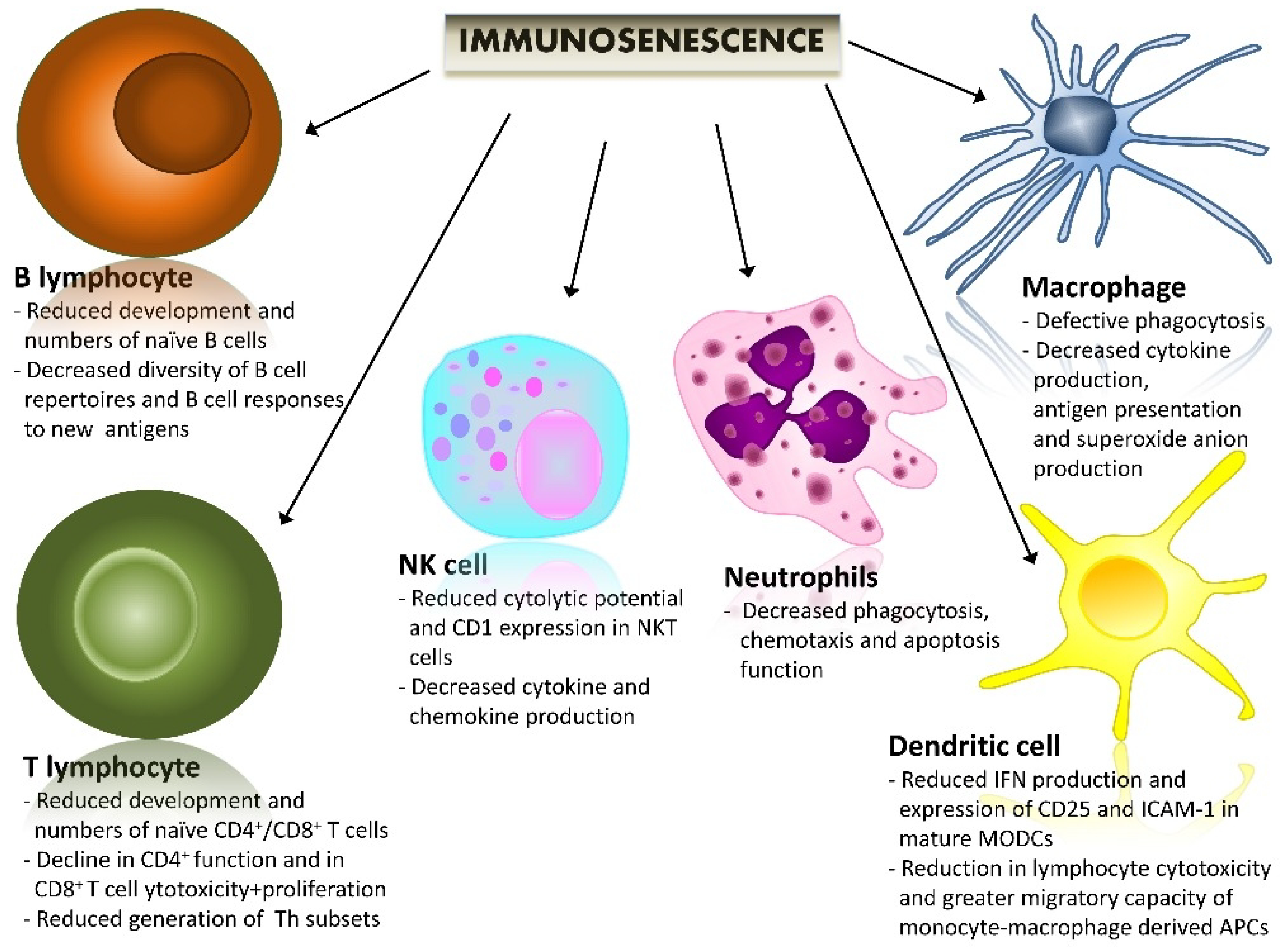

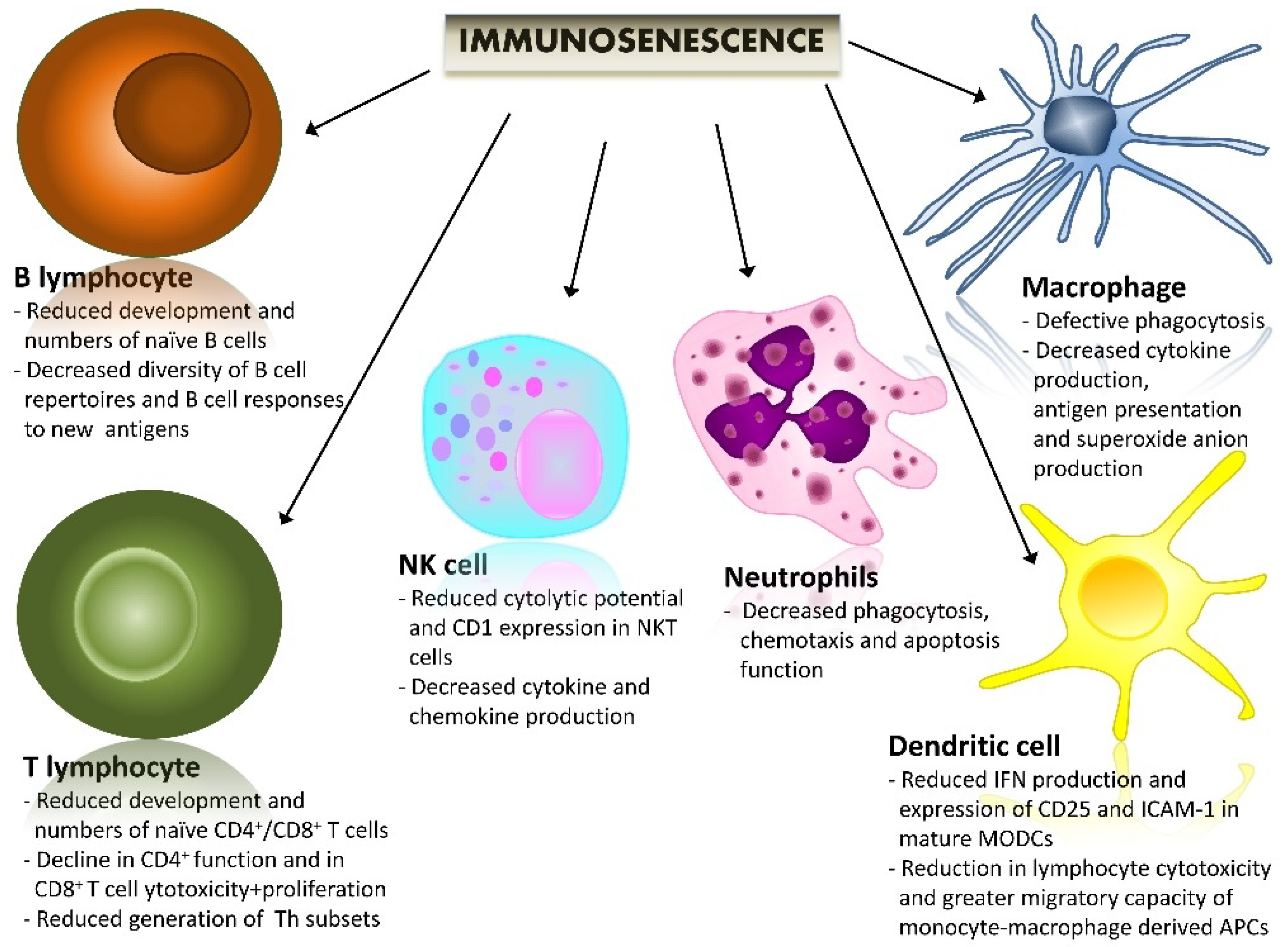{kind=link}
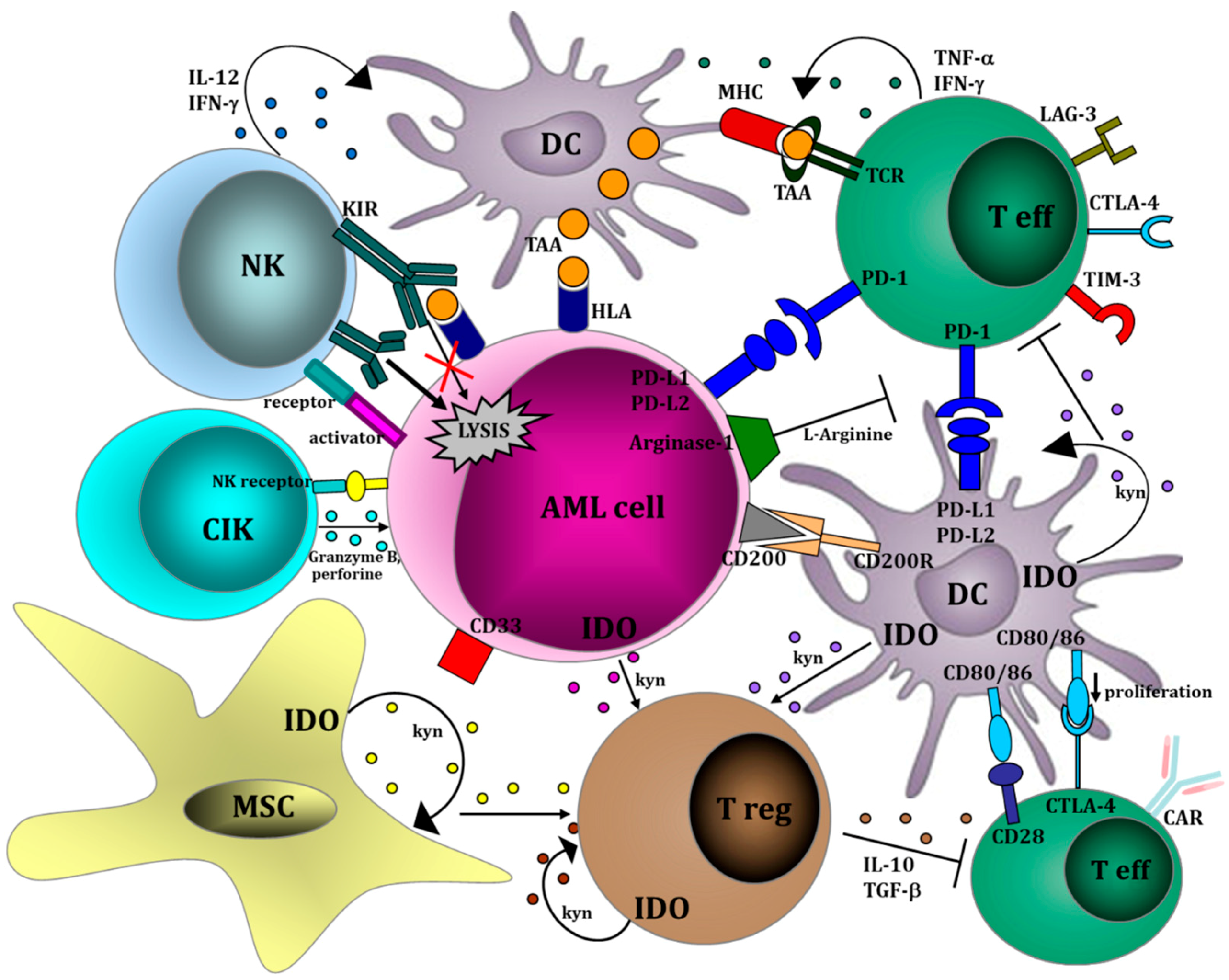{kind=link}
| Pathway | Therapeutical Action | Effect | |
|---|---|---|---|
| Type | Mechanism | ||
| Checkpoint inhibitors | PD-1/PD-L1 |
|
|
| KIR |
|
| |
| Tolerogenic molecules | Arginine |
|
|
| IDO |
|
| |
| Adoptive cell-therapy | NK cells |
|
|
| CAR-T cells |
|
| |
| TCR-edited T cells |
|
| |
| Antigens/Dendritic cells loaded with antigens | WT1, RHAMM, PR-3, DC/WT1 |
|
|
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isidori, A.; Loscocco, F.; Ciciarello, M.; Corradi, G.; Lecciso, M.; Ocadlikova, D.; Parisi, S.; Salvestrini, V.; Amadori, S.; Visani, G.; et al. Immunosenescence and Immunotherapy in Elderly Acute Myeloid Leukemia Patients: Time for a Biology-Driven Approach. Cancers 2018, 10, 211. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10070211
Isidori A, Loscocco F, Ciciarello M, Corradi G, Lecciso M, Ocadlikova D, Parisi S, Salvestrini V, Amadori S, Visani G, et al. Immunosenescence and Immunotherapy in Elderly Acute Myeloid Leukemia Patients: Time for a Biology-Driven Approach. Cancers. 2018; 10(7):211. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10070211
Chicago/Turabian StyleIsidori, Alessandro, Federica Loscocco, Marilena Ciciarello, Giulia Corradi, Mariangela Lecciso, Darina Ocadlikova, Sarah Parisi, Valentina Salvestrini, Sergio Amadori, Giuseppe Visani, and et al. 2018. "Immunosenescence and Immunotherapy in Elderly Acute Myeloid Leukemia Patients: Time for a Biology-Driven Approach" Cancers 10, no. 7: 211. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10070211