A DNA Vaccine Encoding SA-4-1BBL Fused to HPV-16 E7 Antigen Has Prophylactic and Therapeutic Efficacy in a Cervical Cancer Mouse Model

 ,
,  , , , and
, , , and 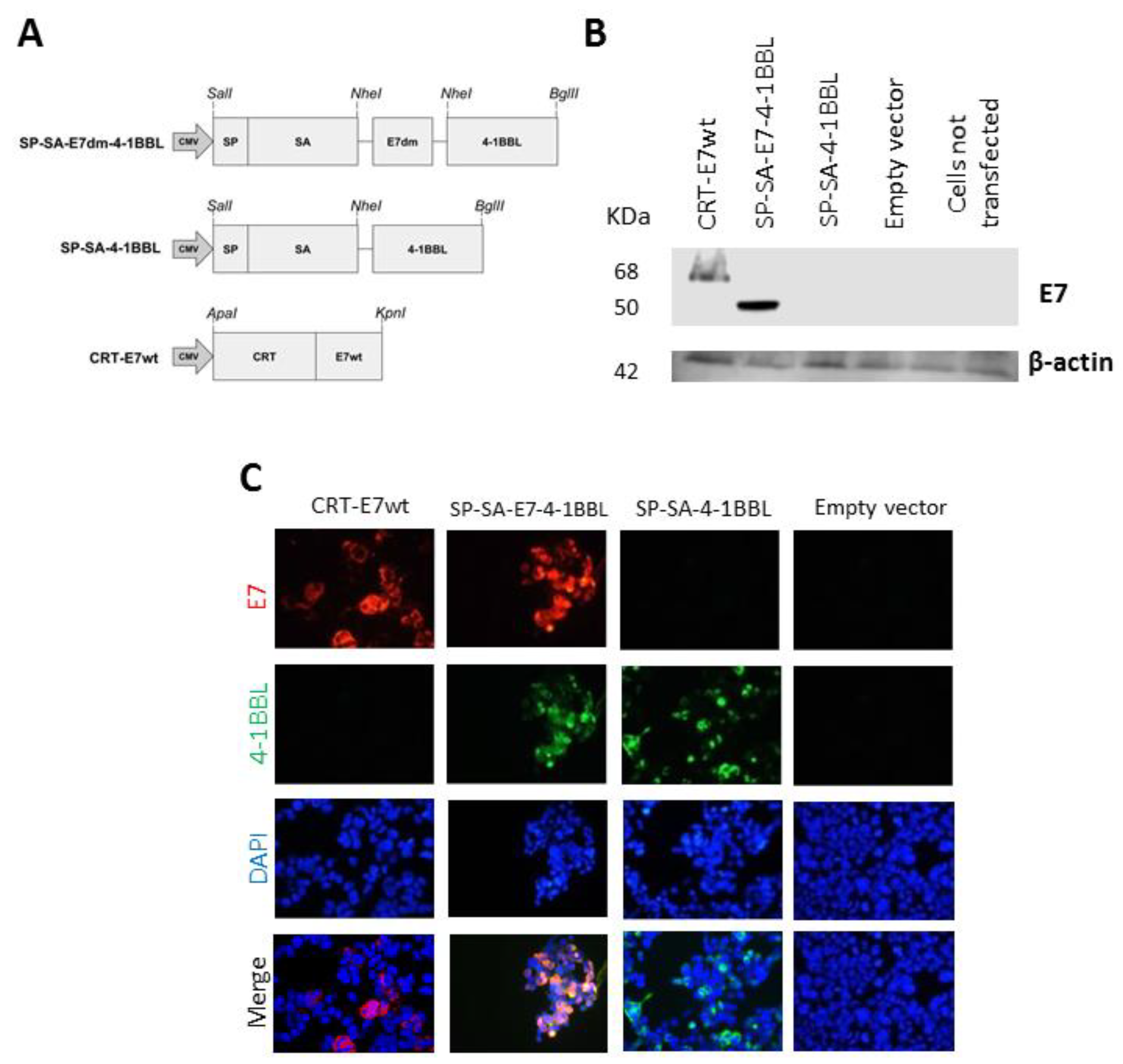{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Detection of Fusion Proteins from DNA Vaccine Plasmids
2.2. Vaccination with SP-SA-E7-4-1BBL Protected Mice against TC-1 Tumor Challenge
2.3. Vaccination with SP-SA-E7-4-1BBL Induces a Therapeutic Effect against Established TC-1 Tumors
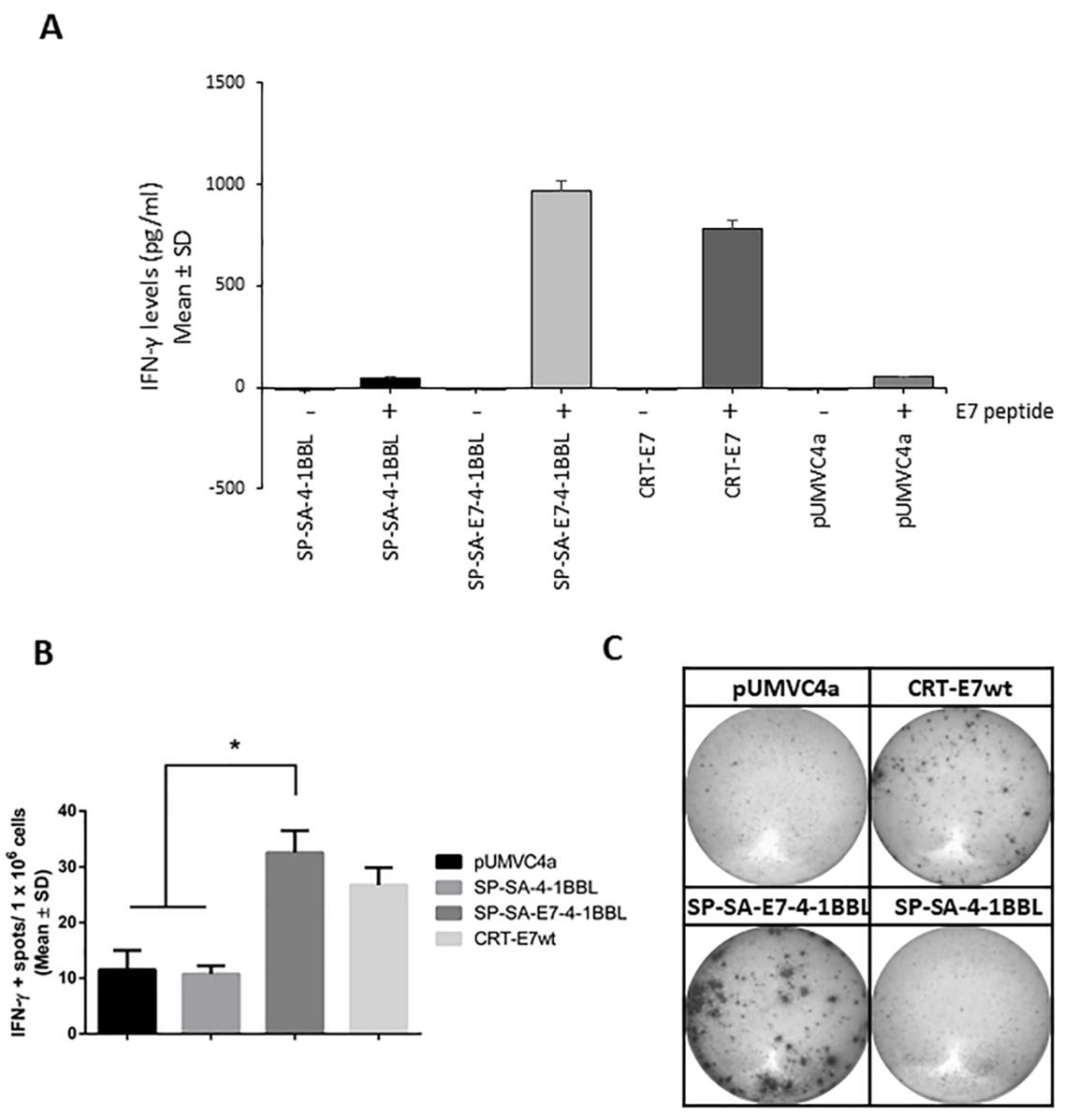
2.4. Immunization with SP-SA-E7-4-1BBL DNA Construct Induces E7-Specific IFN-γ Production
3. Discussion
4. Materials and Methods
4.1. DNA Constructs
4.2. Cell Lines and Culture Conditions
4.3. Immunofluorescence and Western Blot Analysis
4.4. DNA Vaccine Preparation
4.5. Mice
4.6. In Vivo Prophylactic Vaccinations
4.7. In Vivo Therapeutic Vaccinations
4.8. Antigen-Specific Interferon (IFN)-γ Quantification by ELISpot and ELISA Assays
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharpe, A.H. Mechanisms of costimulation. Immunol. Rev. 2009, 229, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liechtenstein, T.; Dufait, I.; Lanna, A.; Breckpot, K.; Escors, D. Modulating co-stimulation during antigen presentation to enhance cancer immunotherapy. Immunol. Endocr. Metab. Agents Med. Chem. 2012, 12, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Madireddi, S.; Schabowsky, R.H.; Srivastava, A.K.; Sharma, R.K.; Yolcu, E.S.; Shirwan, H. SA-4-1BBL costimulation inhibits conversion of conventional CD4+ T cells into CD4+ FoxP3+ T regulatory cells by production of IFN-gamma. PLoS ONE 2012, 7, e42459. [Google Scholar] [CrossRef]
- Shirwan, H.; Sharma, R.K.; Srivastava, A.K.; Yolcu, E.S. Co-stimulatory tumor necrosis factor ligands as adjuvants for the development of subunit-based anticancer vaccines. Oncoimmunology 2013, 2, e23440. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.S.; Weissman, S.M. CDNA sequences of two inducible T-cell genes. Proc. Natl. Acad. Sci. USA 1989, 86, 1963–1967. [Google Scholar] [CrossRef] [PubMed]
- Pollok, K.E.; Kim, Y.J.; Zhou, Z.; Hurtado, J.; Kim, K.K.; Pickard, R.T.; Kwon, B.S. Inducible T cell antigen 4-1BB. Analysis of expression and function. J. Immunol. 1993, 150, 771–781. [Google Scholar] [PubMed]
- Melero, I.; Johnston, J.V.; Shufford, W.W.; Mittler, R.S.; Chen, L. NK1.1 cells express 4-1BB (CDw137) costimulatory molecule and are required for tumor immunity elicited by anti-4-1BB monoclonal antibodies. Cell. Immunol. 1998, 190, 167–172. [Google Scholar] [CrossRef] [PubMed]
- DeBenedette, M.A.; Shahinian, A.; Mak, T.W.; Watts, T.H. Costimulation of CD28-T lymphocytes by 4-1BB ligand. J. Immunol. 1997, 158, 551–559. [Google Scholar]
- Pollok, K.E.; Kim, Y.J.; Hurtado, J.; Zhou, Z.; Kim, K.K.; Kwon, B.S. 4-1BB T-cell antigen binds to mature B cells and macrophages, and costimulates anti-mu-primed splenic B cells. Eur. J. Immunol. 1994, 24, 367–374. [Google Scholar] [CrossRef]
- Vinay, D.S.; Kwon, B.S. Role of 4-1BB in immune responses. Semin. Immunol. 1998, 10, 481–489. [Google Scholar] [CrossRef]
- Croft, M. Co-stimulatory members of the TNFR family: Keys to effective T-cell immunity? Nat. Rev. Immunol. 2003, 3, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Watts, T.H. TNF/TNFR family members in costimulation of T cell responses. Annu. Rev. Immunol. 2005, 23, 23–68. [Google Scholar] [CrossRef] [PubMed]
- Gramaglia, I.; Cooper, D.; Miner, K.T.; Kwon, B.S.; Croft, M. Co-stimulation of antigen-specific CD4 T cells by 4-1BB ligand. Eur. J. Immunol. 2000, 30, 392–402. [Google Scholar] [CrossRef]
- Shuford, W.W.; Klussman, K.; Tritchler, D.D.; Loo, D.T.; Chalupny, J.; Siadak, A.W.; Brown, T.J.; Emswiler, J.; Raecho, H.; Larsen, C.P.; et al. 4-1BB costimulatory signals preferentially induce CD8+ T cell proliferation and lead to the amplification in vivo of cytotoxic T cell responses. J. Exp. Med. 1997, 186, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, C.; Mittler, R.S.; Vella, A.T. Cutting edge: 4-1BB is a bona fide CD8 T cell survival signal. J. Immunol. 1999, 162, 5037–5040. [Google Scholar] [PubMed]
- Won, E.Y.; Cha, K.; Byun, J.S.; Kim, D.U.; Shin, S.; Ahn, B.; Kim, Y.H.; Rice, A.J.; Walz, T.; Kwon, B.S.; et al. The structure of the trimer of human 4-1BB ligand is unique among members of the tumor necrosis factor superfamily. J. Biol. Chem. 2010, 285, 9202–9210. [Google Scholar] [CrossRef] [PubMed]
- Rabu, C.; Quemener, A.; Jacques, Y.; Echasserieau, K.; Vusio, P.; Lang, F. Production of recombinant human trimeric CD137L (4-1BBL). Cross-linking is essential to its T cell co-stimulation activity. J. Biol. Chem. 2005, 280, 41472–41481. [Google Scholar] [CrossRef] [PubMed]
- Elpek, K.G.; Yolcu, E.S.; Franke, D.D.; Lacelle, C.; Schabowsky, R.H.; Shirwan, H. Ex vivo expansion of CD4+ CD25+ FoxP3+ T regulatory cells based on synergy between IL-2 and 4-1BB signaling. J. Immunol. 2007, 179, 7295–7304. [Google Scholar] [CrossRef]
- Schabowsky, R.H.; Elpek, K.G.; Madireddi, S.; Sharma, R.K.; Yolcu, E.S.; Bandura-Morgan, L.; Miller, R.; MacLeod, K.J.; Mittler, R.S.; Shirwan, H. A novel form of 4-1BBL has better immunomodulatory activity than an agonistic anti-4-1BB Ab without Ab-associated severe toxicity. Vaccine 2009, 28, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.K.; Srivastava, A.K.; Yolcu, E.S.; MacLeod, K.J.; Schabowsky, R.H.; Madireddi, S.; Shirwan, H. SA-4-1BBL as the immunomodulatory component of a HPV-16 E7 protein based vaccine shows robust therapeutic efficacy in a mouse cervical cancer model. Vaccine 2010, 28, 5794–5802. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.K.; Sharma, R.K.; Yolcu, E.S.; Ulker, V.; MacLeod, K.; Dinc, G.; Shirwan, H. Prime-boost vaccination with SA-4-1BBL costimulatory molecule and survivin eradicates lung carcinoma in CD8+ T and NK cell dependent manner. PLoS ONE 2012, 7, e48463. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Yolcu, E.S.; Srivastava, A.K.; Shirwan, H. CD4+ T cells play a critical role in the generation of primary and memory antitumor immune responses elicited by SA-4-1BBL and TAA-based vaccines in mouse tumor models. PLoS ONE 2013, 8, e73145. [Google Scholar] [CrossRef]
- Sharma, R.K.; Schabowsky, R.H.; Srivastava, A.K.; Elpek, K.G.; Madireddi, S.; Zhao, H.; Zhong, Z.; Miller, R.W.; Macleod, K.J.; Yolcu, E.S.; et al. 4-1BB ligand as an effective multifunctional immunomodulator and antigen delivery vehicle for the development of therapeutic cancer vaccines. Cancer Res. 2010, 70, 3945–3954. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Danishmalik, S.N.; Sin, J.I. DNA vaccines, electroporation and their applications in cancer treatment. Hum. Vaccines Immunother. 2015, 11, 1889–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Trujillo, J.J.; Robles-Rodriguez, O.A.; Garza-Morales, R.; Garcia-Garcia, A.; Rodriguez-Rocha, H.; Villanueva-Olivo, A.; Segoviano-Ramirez, J.C.; Esparza-Gonzalez, S.C.; Saucedo-Cardenas, O.; Montes-de-Oca-Luna, R.; et al. Antitumor Response by Endoplasmic Reticulum-Targeting DNA Vaccine is Improved by Adding a KDEL Retention Signal. Nucleic Acid. Ther. 2018, 28, 252–261. [Google Scholar] [CrossRef]
- Lin, K.; Doolan, K.; Hung, C.F.; Wu, T.C. Perspectives for preventive and therapeutic HPV vaccines. J. Formos. Med. Assoc. 2010, 109, 4–24. [Google Scholar] [CrossRef]
- Lowe, D.B.; Shearer, M.H.; Kennedy, R.C. DNA vaccines: Successes and limitations in cancer and infectious disease. J. Cell. Biochem. 2006, 98, 235–242. [Google Scholar] [CrossRef]
- Cheng, M.A.; Farmer, E.; Huang, C.; Lin, J.; Hung, C.F.; Wu, T.C. Therapeutic DNA vaccines for human papillomavirus and associated diseases. Hum. Gene Ther. 2018, 29, 971–996. [Google Scholar] [CrossRef]
- Ragonnaud, E.; Andersson, A.M.; Pedersen, A.E.; Laursen, H.; Holst, P.J. An adenoviral cancer vaccine co-encoding a tumor associated antigen together with secreted 4-1BBL leads to delayed tumor progression. Vaccine 2016, 34, 2147–2156. [Google Scholar] [CrossRef]
- Perez-Trujillo, J.J.; Garza-Morales, R.; Barron-Cantu, J.A.; Figueroa-Parra, G.; Garcia-Garcia, A.; Rodriguez-Rocha, H.; Garcia-Juarez, J.; Munoz-Maldonado, G.E.; Saucedo-Cardenas, O.; Montes-De-Oca-Luna, R.; et al. DNA vaccine encoding human papillomavirus antigens flanked by a signal peptide and a KDEL sequence induces a potent therapeutic antitumor effect. Oncol. Lett. 2017, 13, 1569–1574. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Gutierrez, J.G.; Elpek, K.G.; Montes de Oca-Luna, R.; Shirwan, H.; Sam Zhou, H.; McMasters, K.M. Vaccination with an adenoviral vector expressing calreticulin-human papillomavirus 16 E7 fusion protein eradicates E7 expressing established tumors in mice. Cancer Immunol. Immunother. 2007, 56, 997–1007. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garza-Morales, R.; Perez-Trujillo, J.J.; Martinez-Jaramillo, E.; Saucedo-Cardenas, O.; Loera-Arias, M.J.; Garcia-Garcia, A.; Rodriguez-Rocha, H.; Yolcu, E.; Shirwan, H.; Gomez-Gutierrez, J.G.; et al. A DNA Vaccine Encoding SA-4-1BBL Fused to HPV-16 E7 Antigen Has Prophylactic and Therapeutic Efficacy in a Cervical Cancer Mouse Model. Cancers 2019, 11, 96. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010096
Garza-Morales R, Perez-Trujillo JJ, Martinez-Jaramillo E, Saucedo-Cardenas O, Loera-Arias MJ, Garcia-Garcia A, Rodriguez-Rocha H, Yolcu E, Shirwan H, Gomez-Gutierrez JG, et al. A DNA Vaccine Encoding SA-4-1BBL Fused to HPV-16 E7 Antigen Has Prophylactic and Therapeutic Efficacy in a Cervical Cancer Mouse Model. Cancers. 2019; 11(1):96. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010096
Chicago/Turabian StyleGarza-Morales, Rodolfo, Jose J. Perez-Trujillo, Elvis Martinez-Jaramillo, Odila Saucedo-Cardenas, Maria J. Loera-Arias, Aracely Garcia-Garcia, Humberto Rodriguez-Rocha, Esma Yolcu, Haval Shirwan, Jorge G. Gomez-Gutierrez, and et al. 2019. "A DNA Vaccine Encoding SA-4-1BBL Fused to HPV-16 E7 Antigen Has Prophylactic and Therapeutic Efficacy in a Cervical Cancer Mouse Model" Cancers 11, no. 1: 96. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010096