Treatment Combining CD200 Immune Checkpoint Inhibitor and Tumor-Lysate Vaccination after Surgery for Pet Dogs with High-Grade Glioma

 , , , and
, , , and
Abstract
:1. Introduction
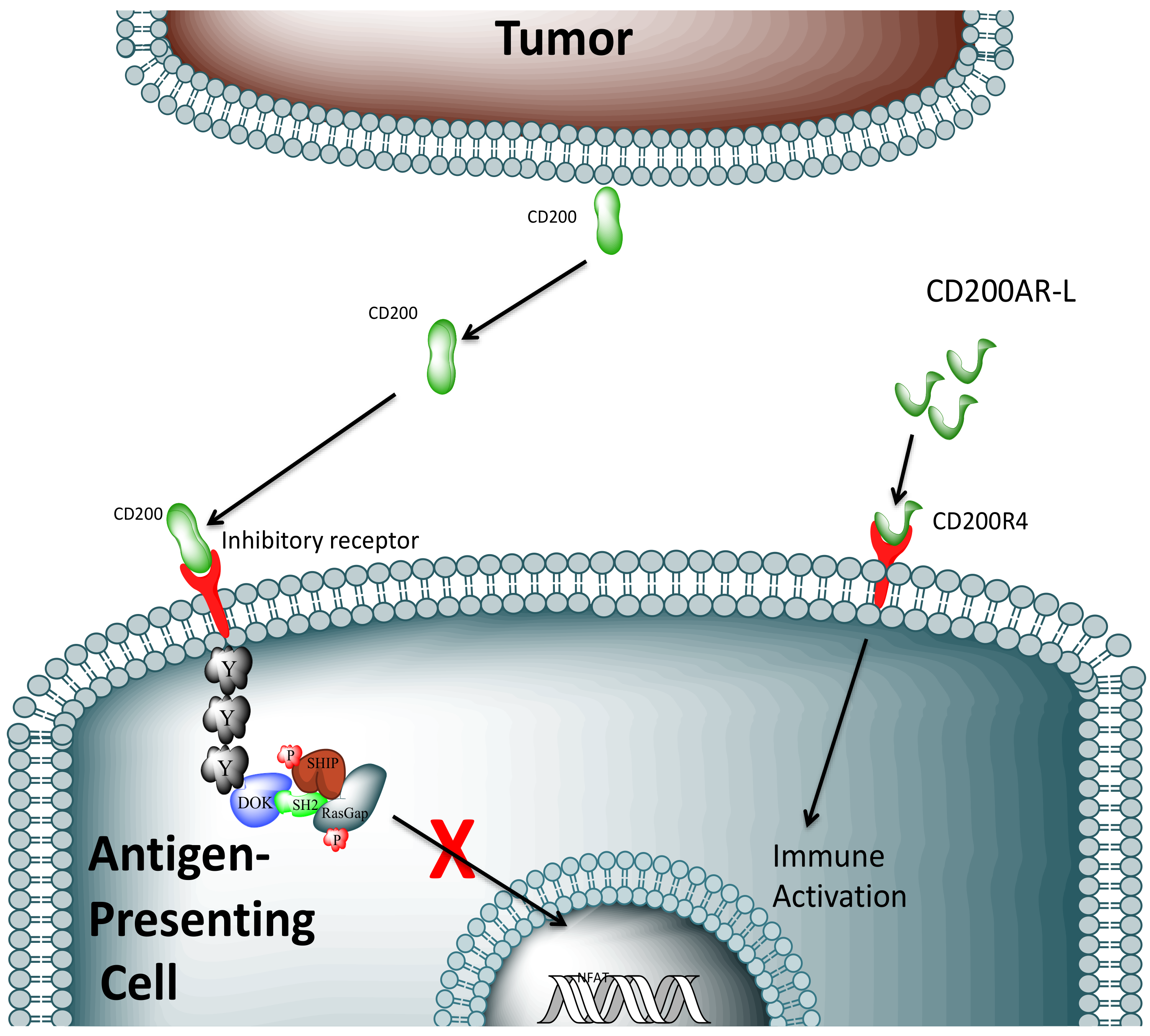
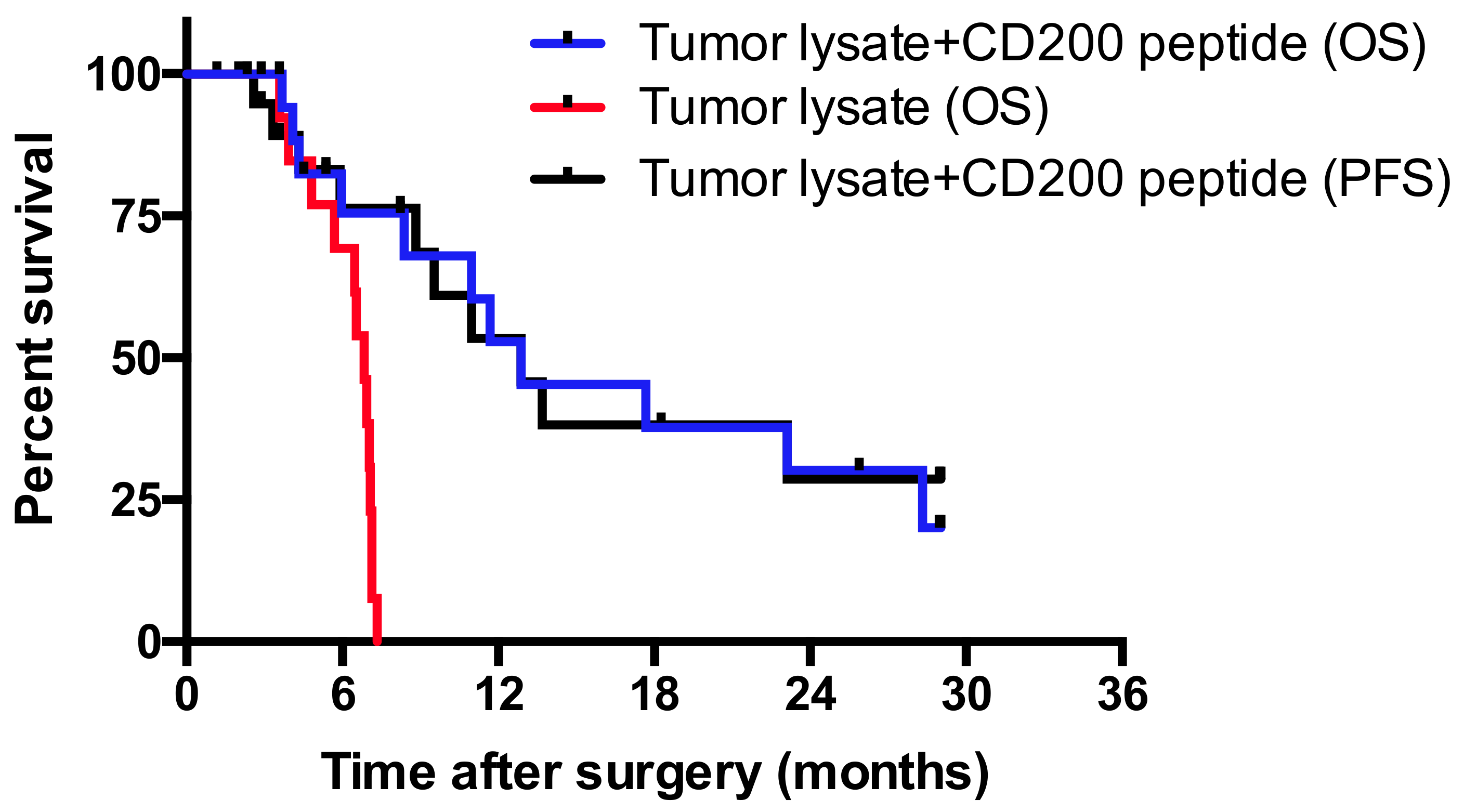
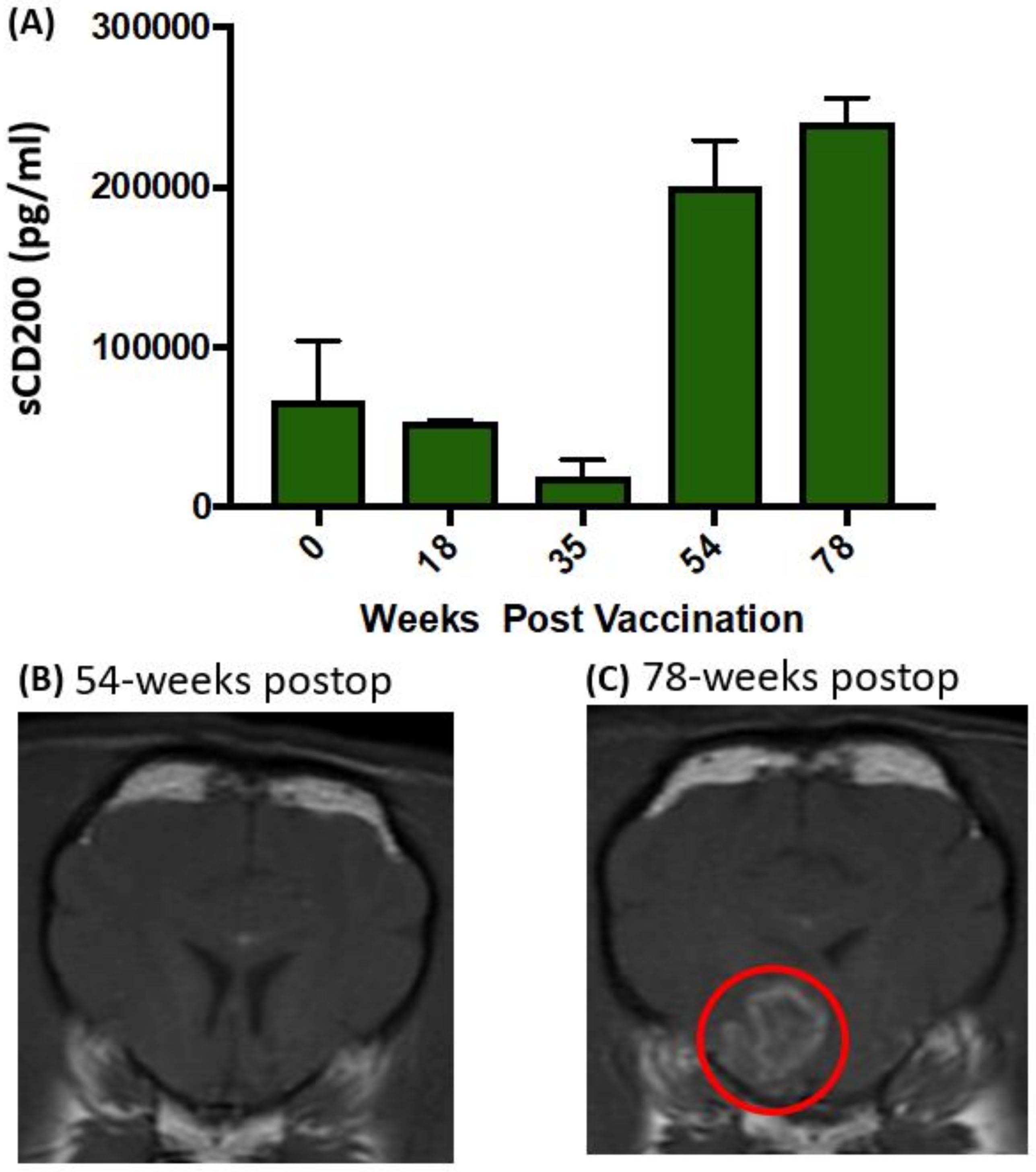
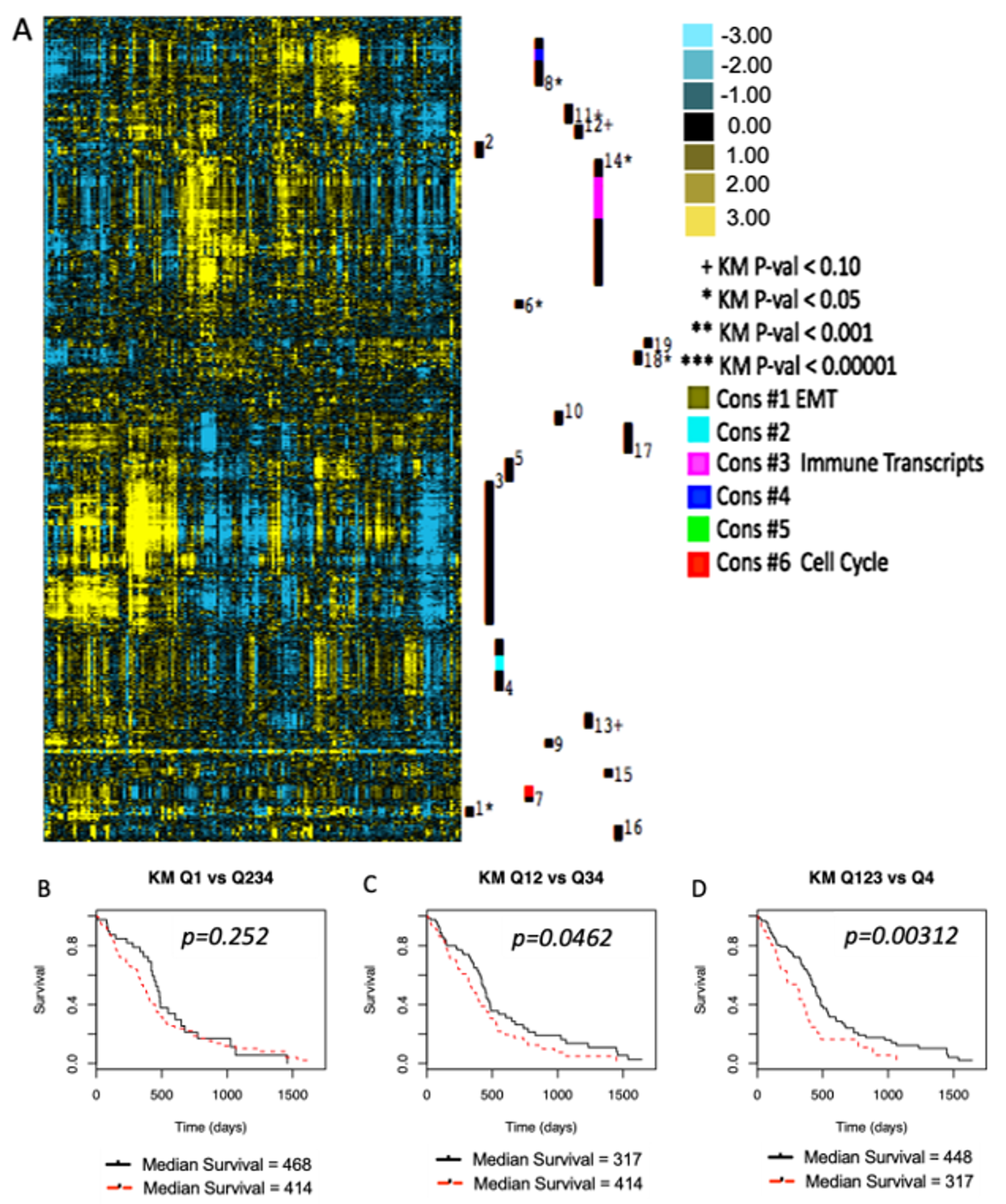
2. Results
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. TCGA Analysis
4.3. Canine Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ascierto, P.A.; Daniele, B.; Hammers, H.; Hirsh, V.; Kim, J.; Licitra, L.; Nanda, R.; Pignata, S. Perspectives in immunotherapy: Meeting report from the “Immunotherapy Bridge”, Napoli, November 30th, 2016. J. Transl. Med. 2017, 15, 205. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesely, M.D.; Schreiber, R.D. Cancer immunoediting: Antigens, mechanisms, and implications to cancer immunotherapy. Ann. N. Y. Acad. Sci. 2013, 1284, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kimmelman, J.; Nalbantoglu, J. Faithful companions: A proposal for neurooncology trials in pet dogs. Cancer Res. 2007, 67, 4541–4544. [Google Scholar] [CrossRef] [PubMed]
- Paoloni, M.; Khanna, C. Translation of new cancer treatments from pet dogs to humans. Nat. Rev. Cancer 2008, 8, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.; Khanna, C. Spontaneous and genetically engineered animal models; use in preclinical cancer drug development. Eur. J. Cancer 2004, 40, 858–880. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, M.E.; Arnold, S.; Bin Zahid, A.; McPheeters, M.; Gerard O’Sullivan, M.; Tabaran, A.F.; Hunt, M.A.; Pluhar, G.E. Naturally occurring canine glioma as a model for novel therapeutics. Cancer Investig. 2018, 36, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Bentley, R.T.; Ahmed, A.U.; Yanke, A.B.; Cohen-Gadol, A.A.; Dey, M. Dogs are man’s best friend: In sickness and in health. Neuro Oncol. 2017, 19, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.; Platt, S.; Kent, M.; Haley, A. Canine brain tumours: A model for the human disease? Vet. Comp. Oncol. 2017, 15, 252–272. [Google Scholar] [CrossRef] [PubMed]
- Moertel, C.L.; Xia, J.; LaRue, R.; Waldron, N.N.; Andersen, B.M.; Prins, R.M.; Okada, H.; Donson, A.M.; Foreman, N.K.; Hunt, M.A.; Pennell, C.A. CD200 in CNS tumor-induced immunosuppression: The role for CD200 pathway blockade in targeted immunotherapy. J. Immunother. Cancer 2014, 2, 46. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Louveau, A.; Harris, T.H.; Kipnis, J. Revisiting the Mechanisms of CNS Immune Privilege. Trends Immunol. 2015, 36, 569–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, C.M.; Lim, M. Immunotherapy for glioblastoma: Playing chess, not checkers. Clin. Cancer Res. 2018, 24, 4059–4061. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.P.; Morford, L.A.; Chougnet, C.; Dix, A.R.; Brooks, A.G.; Torres, N.; Shuman, J.D.; Coligan, J.E.; Brooks, W.H.; Roszman, T.L.; et al. Human glioma-induced immunosuppression involves soluble factor(s) that alters monocyte cytokine profile and surface markers. J. Immunol. 1999, 162, 4882–4892. [Google Scholar] [PubMed]
- Chen, Z.; Hambardzumyan, D. Immune Microenvironment in Glioblastoma Subtypes. Front. Immunol. 2018, 9, 1004. [Google Scholar] [CrossRef] [PubMed]
- Suñol, A.; Mascort, J.; Font, C.; Bastante, A.R.; Pumarola, M.; Feliu-Pascual, A.L. Long-term follow-up of surgical resection alone for primary intracranial rostrotentorial tumors in dogs: 29 cases (2002–2013). Open Vet. J. 2017, 7, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Ampudia-Mesias, E.; Shaver, R.; Horbinski, C.M.; Moertel, C.L.; Olin, M.R. Tumor-derived vaccines containing CD200 inhibit immune activation: Implications for immunotherapy. Immunotherapy 2016, 8, 1059–1071. [Google Scholar] [CrossRef]
- Greenwald, R.J.; Freeman, G.J.; Sharpe, A.H. The B7 family revisited. Annu. Rev. Immunol. 2005, 23, 515–548. [Google Scholar] [CrossRef] [PubMed]
- Erin, N.; Podnos, A.; Tanriover, G.; Duymuş, Ö.; Cote, E.; Khatri, I.; Gorczynski, R.M. Bidirectional effect of CD200 on breast cancer development and metastasis, with ultimate outcome determined by tumor aggressiveness and a cancer-induced inflammatory response. Oncogene 2015, 34, 3860–3870. [Google Scholar] [CrossRef]
- Kretz-Rommel, A.; Qin, F.; Dakappagari, N.; Ravey, E.P.; McWhirter, J.; Oltean, D.; Frederickson, S.; Maruyama, T.; Wild, M.A.; Nolan, M.J.; et al. CD200 expression on tumor cells suppresses antitumor immunity: New approaches to cancer immunotherapy. J. Immunol. 2007, 178, 5595–5605. [Google Scholar] [CrossRef]
- Moreaux, J.; Veyrune, J.L.; Reme, T.; De Vos, J.; Klein, B. CD200: A putative therapeutic target in cancer. Biochem. Biophys. Res. Commun. 2008, 366, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Tonks, A.; Hills, R.; White, P.; Rosie, B.; Mills, K.I.; Burnett, A.K.; Darley, R.L. CD200 as a prognostic factor in acute myeloid leukaemia. Leukemia 2007, 21, 566–568. [Google Scholar] [CrossRef] [Green Version]
- Petermann, K.B.; Rozenberg, G.I.; Zedek, D.; Groben, P.; McKinnon, K.; Buehler, C.; Kim, W.Y.; Shields, J.M.; Penland, S.; Bear, J.E.; et al. CD200 is induced by ERK and is a potential therapeutic target in melanoma. J. Clin. Investig. 2007, 117, 3922–3929. [Google Scholar] [CrossRef] [Green Version]
- Siva, A.; Xin, H.; Qin, F.; Oltean, D.; Bowdish, K.S.; Kretz-Rommel, A. Immune modulation by melanoma and ovarian tumor cells through expression of the immunosuppressive molecule CD200. Cancer Immunol. Immunother. 2008, 57, 987–996. [Google Scholar] [CrossRef]
- Stumpfova, M.; Ratner, D.; Desciak, E.B.; Eliezri, Y.D.; Owens, D.M. The immunosuppressive surface ligand CD200 augments the metastatic capacity of squamous cell carcinoma. Cancer Res. 2010, 70, 2962–2972. [Google Scholar] [CrossRef]
- Twito, T.; Chen, Z.; Khatri, I.; Wong, K.; Spaner, D.; Gorczynski, R. Ectodomain shedding of CD200 from the B-CLL cell surface is regulated by ADAM28 expression. Leuk. Res. 2013, 37, 816–821. [Google Scholar] [CrossRef]
- Wong, K.; Brenneman, F.; Chesney, A.; Spaner, D.E.; Gorczynski, R.M. Soluble CD200 is critical to engraft chronic lymphocytic leukemia cells in immunocompromised mice. Cancer Res. 2012, 72, 4931–4943. [Google Scholar] [CrossRef]
- Gorczynski, R.; Chen, Z.; Kai, Y.; Lee, L.; Wong, S.; Marsden, P.A. CD200 is a ligand for all members of the CD200R family of immunoregulatory molecules. J. Immunol. 2004, 172, 7744–7749. [Google Scholar] [CrossRef]
- Gorczynski, R.; Chen, Z.; Khatri, I.; Yu, K. sCD200 present in mice receiving cardiac and skin allografts causes immunosuppression in vitro and induces Tregs. Transplantation 2013, 95, 442–447. [Google Scholar] [CrossRef]
- Gorczynski, R.M.; Lee, L.; Boudakov, I. Augmented Induction of CD4+ CD25+ Treg using monoclonal antibodies to CD200R. Transplantation 2005, 79, 1180–1183. [Google Scholar] [CrossRef]
- Olin, M.R.; Andersen, B.M.; Zellmer, D.M.; Grogan, P.T.; Popescu, F.E.; Xiong, Z.; Forster, C.L.; Seiler, C.; SantaCruz, K.S.; Chen, W.; et al. Superior efficacy of tumor cell vaccines grown in physiologic oxygen. Clin. Cancer Res. 2010, 16, 4800–4808. [Google Scholar] [CrossRef]
- Olin, M.R.; Andersen, B.M.; Litterman, A.J.; Grogan, P.T.; Sarver, A.L.; Robertson, P.T.; Liang, X.; Chen, W.; Parney, I.F.; Hunt, M.A.; et al. Oxygen is a master regulator of the immunogenicity of primary human glioma cells. Cancer Res. 2011, 71, 6583–6589. [Google Scholar] [CrossRef]
- Scott, M.C.; Temiz, N.A.; Sarver, A.E.; LaRue, R.S.; Rathe, S.K.; Varshney, J.; Wolf, N.K.; Moriarity, B.S.; O’Brien, T.D.; Spector, L.G.; et al. Comparative Transcriptome Analysis Quantifies Immune Cell Transcript Levels, Metastatic Progression, and Survival in Osteosarcoma. Cancer Res. 2018, 78, 326–337. [Google Scholar] [CrossRef]
- Snyder, J.M.; Shofer, F.S.; Van Winkle, T.J.; Massicotte, C. Canine intracranial primary neoplasia: 173 cases (1986–2003). J. Vet. Intern. Med. 2006, 20, 669–675. [Google Scholar]
- Hatherley, D.; Lea, S.M.; Johnson, S.; Barclay, A.N. Structures of CD200/CD200 receptor family and implications for topology, regulation, and evolution. Structure 2013, 21, 820–832. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Name | P-Value | Adjusted P-Value | Z-Score | Combined Score |
|---|---|---|---|---|---|
| 1 | Immune System_Homo sapiens_R-HSA-168256 | 1.116 x10−49 | 8.88x 10−47 | −2.23 | 251.61 |
| 2 | Immunoregulatory interactions between a Lymphoid and non-Lymphoid cell_Homo sapiens_R-HSA-198933 | 4.226 x 10−37 | 1.682 x 10−34 | −2.00 | 167.65 |
| 3 | Extracellular matrix organization_Homo sapiens_R-HSA-1474244 | 1.460 x 10−29 | 3.875 x 10−27 | −2.10 | 139.12 |
| 4 | Adaptive Immune System_Homo sapiens_R-HSA-1280218 | 2.066 x 10−21 | 3.290 x 10−19 | −2.25 | 107.12 |
| 5 | Class A/1 (Rhodopsin-like receptors) Homo sapiens_R-HSA-373076 | 9.042 x 10−21 | 1.200 x 10−18 | −2.10 | 96.97 |
| 6 | Chemokine receptors bind chemokines_Homo sapiens_R-HSA-380108 | 2.178 x 10−22 | 4.334 x 10−20 | −1.93 | 96.39 |
| 7 | Cytokine Signallind in Immune system_Homo sapiens_R-HSA-1280215 | 5.330 x 10−17 | 6.061 x 10−15 | −2.35 | 88.11 |
| 8 | GCPR ligand binding_Homo sapiens_R-HSA-500792 | 1.172 x 10−15 | 1.166 x 10−13 | −2.20 | 75.57 |
| 9 | Innate Immune System_Homo sapiens_R-HSA-168249 | 1.917 x 10−13 | 1.272 x 10−11 | −2.33 | 68.28 |
| 10 | Peptide ligand-binding receptors_Homo sapiens_R-HSA-375276 | 1.771 x 10−15 | 1.566 x 10−13 | −1.91 | 64.80 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olin, M.R.; Ampudia-Mesias, E.; Pennell, C.A.; Sarver, A.; Chen, C.C.; Moertel, C.L.; Hunt, M.A.; Pluhar, G.E. Treatment Combining CD200 Immune Checkpoint Inhibitor and Tumor-Lysate Vaccination after Surgery for Pet Dogs with High-Grade Glioma. Cancers 2019, 11, 137. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11020137
Olin MR, Ampudia-Mesias E, Pennell CA, Sarver A, Chen CC, Moertel CL, Hunt MA, Pluhar GE. Treatment Combining CD200 Immune Checkpoint Inhibitor and Tumor-Lysate Vaccination after Surgery for Pet Dogs with High-Grade Glioma. Cancers. 2019; 11(2):137. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11020137
Chicago/Turabian StyleOlin, Michael R., Elisabet Ampudia-Mesias, Christopher A. Pennell, Aaron Sarver, Clark C. Chen, Christopher L. Moertel, Matthew A. Hunt, and G. Elizabeth Pluhar. 2019. "Treatment Combining CD200 Immune Checkpoint Inhibitor and Tumor-Lysate Vaccination after Surgery for Pet Dogs with High-Grade Glioma" Cancers 11, no. 2: 137. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11020137