Tiny miRNAs Play a Big Role in the Treatment of Breast Cancer Metastasis
by
 , , , ,
, , , ,
Andrea York Tiang Teo
1,2 ,
,
Xiaoqiang Xiang
3,
Minh TN Le
4,
Andrea Li-Ann Wong
1,2,5,
Qi Zeng
6,
Lingzhi Wang
2,4,* and
Boon-Cher Goh
1,2,4,5,* 1
Department of Medicine, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 117600, Singapore
2
Cancer Science Institute of Singapore, National University of Singapore, Singapore 117599, Singapore
3
Department of Clinical Pharmacy, School of Pharmacy, Fudan University, Shanghai 20203, China
4
Institute for Digital Medicine and Department of Pharmacology, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 117600, Singapore
5
Department of Haematology–Oncology, National University Cancer Institute, Singapore 119228, Singapore
6
Institute of Molecular and Cell Biology, Agency for Science, Technology and Research (A*STAR), Singapore 138673, Singapore
*
Authors to whom correspondence should be addressed.
Cancers 2021, 13(2), 337; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13020337
Submission received: 18 December 2020
/
Revised: 14 January 2021
/
Accepted: 14 January 2021
/
Published: 18 January 2021
(This article belongs to the Special Issue Targeting Cancer Metastasis)
Abstract
:Simple Summary
MicroRNAs (miRNAs) have emerged as important regulators of tumour progression and metastasis in breast cancer. Through a review of multiple studies, this paper has identified the key regulatory roles of oncogenic miRNAs in breast cancer metastasis including the potentiation of angiogenesis, epithelial-mesenchymal transition, the Warburg effect, and the tumour microenvironment. Several approaches have been studied for selective targeting of breast tumours by miRNAs, ranging from delivery systems such as extracellular vesicles and liposomes to the use of prodrugs and functionally modified vehicle-free miRNAs. While promising, these miRNA-based therapies face challenges including toxicity and immunogenicity, and greater research on their safety profiles must be performed before progressing to clinical trials.
Abstract
Distant organ metastases accounts for the majority of breast cancer deaths. Given the prevalence of breast cancer in women, it is imperative to understand the underlying mechanisms of its metastatic progression and identify potential targets for therapy. Since their discovery in 1993, microRNAs (miRNAs) have emerged as important regulators of tumour progression and metastasis in various cancers, playing either oncogenic or tumour suppressor roles. In the following review, we discuss the roles of miRNAs that potentiate four key areas of breast cancer metastasis—angiogenesis, epithelial-mesenchymal transition, the Warburg effect and the tumour microenvironment. We then evaluate the recent developments in miRNA-based therapies in breast cancer, which have shown substantial promise in controlling tumour progression and metastasis. Yet, certain challenges must be overcome before these strategies can be implemented in clinical trials.
1. Introduction
Breast cancer is the most commonly occurring cancer in women and the second-most prevalent cancer overall [1], with the development of distant organ metastases accounting for 90% of breast cancer deaths [2]. Metastasis is a multi-step process characterized by (i) tumour cell invasion into adjacent tissue, (ii) transendothelial migration of cancer cells into vessels, (iii) survival in the circulatory system, (iv) extravasation to secondary tissue, and (v) subsequent proliferation and colonization in competent organs [3]. Furthermore, the Warburg effect has been shown to facilitate metastatic dissemination by minimizing oxidative stress [4], while the tumour microenvironment induces tumour growth and metastasis via various mechanisms [5].
In the past decade, microRNAs (miRNAs) have emerged as important regulators of various steps in tumour progression and metastasis. miRNAs are small non-coding RNA molecules of 19 to 24 nucleotides, which regulate gene expression in a sequence-specific fashion [6]. Following incorporation into the ribonucleoprotein RNA-induced silencing complex (RISC) via association with Argonaute 2, miRNAs often base-pair with the 3′UTRs of target mRNAs, thereby reducing mRNA translation or causing degradation of the mRNA transcript. Alternatively, miRNAs may also bind to the 5′UTR and coding sequence of the mRNA transcript [7,8]. The specific fate of the mRNA depends on the degree of base-pairing complementarity between the miRNA and mRNA [9]. This imperfect match means each miRNA possesses the potential to target multiple different mRNAs [10,11]. Furthermore, crosstalk between miRNAs and long non-coding RNAs (lncRNAs) has also been documented, with these interactions forming complex networks in targeted gene regulation [12]. Yoon et al. described four key interactions between lncRNAs and miRNAs, in which (i) miRNAs may trigger lncRNA decay, (ii) lncRNAs act as miRNA decoys, (iii) lncRNAs and miRNAs compete for target mRNAs, and (iv) lncRNAs generate miRNAs [13]. Consequently, various studies have established that miRNAs elicit either oncogenic or tumour suppressive functions by silencing target protein-coding genes [14,15], indicating the potential for miRNAs to regulate multiple signalling processes necessary for breast cancer progression and metastasis. Thus, the use of miRNAs in breast cancer therapy holds huge potential.
This review will first study the various pathways through which oncogenic miRNAs potentiate breast cancer metastasis (Figure 1), and thereafter discuss the recent developments and challenges in novel miRNA-based therapies for breast cancer.
2. miRNAs in Breast Cancer Metastasis
For malignant cells to metastasise, several key processes must occur (Figure 2). The induction of angiogenesis enables tumour cells to gain access to the vasculature for subsequent metastatic spread to other tissues [16]. Greater vascular density within the tumour increases the chances of tumour cell escape and intravasation [17], while the leaky and fragmented basement membranes of newly formed capillaries increase the ease of tumour cell penetrability and migration [18]. Meanwhile, the neoplastic cells may invade the surrounding stroma via collective migration or epithelial-mesenchymal transition (EMT) [19]. During invasion, the tumour cells disrupt the basement membrane and penetrate the underlying stroma, a process in which regulation of adhesion, ECM reorganization and motility must occur [20,21]. In EMT, tumour cells lose their epithelial properties and gain migratory and invasive traits. Importantly, the loss of epithelial marker E-cadherin from the adherens junctions, along with a change in the upregulation of keratin expression to the mesenchymal intermediate filament vimentin expression, is characteristic of cancer EMT [22]. It is postulated that this switch to a mesenchymal phenotype endows the cells with migratory and stem-like properties as well as reduced cell-cell adhesion [19,23].
Interestingly, the metastatic process does not occur due to the properties of the tumour cells alone. The tumour microenvironment, consisting of a variety of resident and infiltrating host cells, secreted factors and ECM proteins, plays a large role in determining the fate of the cancer cells [24]. It has been implicated in tumorigenesis, tumour progression and metastasis formation. For instance, certain cells such as cancer-associated fibroblasts and M2-type tumour-associated macrophages have been shown to assist angiogenesis [25] and tumour cell invasion, migration and eventual intravasation [26,27], while M1 macrophages have been described as anti-tumour effectors [28]. Furthermore, the unique cancer cell metabolic phenotype, known as the Warburg effect, has been shown to be a significant contributory factor in these processes as well. By constraining the pyruvate flux into mitochondrial oxidative metabolism, the Warburg effect minimizes oxidative stress from mitochondrial respiration within cancer cells. This then facilitates metastatic dissemination by providing cancer cells with a survival advantage [4,29].
Following intravasation and survival in the circulatory system, the tumour cells then extravasate to pre-metastatic niches in secondary sites that are suitable for proliferation and colonization [30]. If colonization is successful, this will result in the formation of a distant metastasis.
miRNAs have been implicated in each stage of cancer metastasis, acting either as tumour suppressors or oncogenic miRNAs to suppress or promote metastasis respectively. In some cases, miRNAs may act as both tumour suppressors and oncogenic miRNAs depending on the type of cancer and cellular context [31]. While most miRNAs are found within the cellular microenvironment, miRNAs have also been detected in the extracellular environment, protected and carried by vesicles such as exosomes [32], or associated with proteins [33]. These extracellular miRNAs function as chemical messengers to mediate cell-cell communication. For instance, exosomal release of various miRNAs by breast cancer cells has been suggested to promote their own survival and invasion, thereby promoting metastasis [34,35]. In addition, the release of miRNAs from multiple cells types could further enhance the metastatic progression of cancer through modulating the tumour microenvironment, as discussed below in Section 2.4. Indeed, multiple stages in the metastatic cascade are tightly regulated by miRNAs. The following subsections will provide a closer look at miRNAs involved in potentiating four key processes of breast cancer metastasis—angiogenesis, EMT and invasion, the Warburg effect and the tumour microenvironment.
2.1. miRNAs in Angiogenesis
Besides its pivotal role in tumour survival and growth, angiogenesis enables tumour cells to break off from the primary tumour and travel to distant sites via the vasculature [36]. Various growth factors and proteins secreted by tumours promote angiogenesis, including VEGF, angiopoietin 1 and 2 and TGF-ß. Notably, hypoxia induces the expression of VEGF and its receptor via HIF1α, and VEGF in turn causes ECM remodelling [37] and proliferation of blood vessels [38]. Angiopoietin 1 mediates vessel maturation, migration and adhesion of endothelial cells, while angiopoietin 2 promotes neovascularisation in conjunction with VEGF [39]. Oncogenic miRNAs have been found to stimulate this angiogenic process by regulating the expression of growth factors and proteins in angiogenesis, thereby promoting breast cancer metastasis (Table 1).
For instance, miR-155 was demonstrated to induce angiogenesis in breast cancer via its target, von Hippel-Lindau, a ubiquitin ligase that targets HIF1α [45]. Notably, extensive angiogenesis, proliferation, tumour necrosis and recruitment of pro-inflammatory cells such as tumour-associated macrophages were observed following mammary fat pad xenotransplantation of miR-155 [41]. In addition, miR-93 was found to promote tumour angiogenesis and metastasis in breast cancer by suppressing tumour suppressor LATS2 [46] expression, with increased lung metastasis demonstrated in a mouse model [43].
VEGF is also a crucial target protein of miRNA regulation in angiogenesis. miR-9-mediated E-cadherin downregulation has been found to play a role in upregulating the expression of the gene encoding VEGF via activation of ß-catenin signalling [40]. miR-20a was also found to induce angiogenic effects in breast cancer cell lines, with its expression associated with increases in mean vessel size, VEGFA expression and the presence of glomeruloid microvascular proliferations [42]. Lastly, miR-21, which regulates multiple pathways in cancer metastasis, also plays a role in tumour angiogenesis in breast cancer. Knockdown of miR-21 demonstrated suppressed tumour growth and angiogenesis by targeting the VEGF/VEGFR2/HIF1α axis in a VEGFR2-luc mouse model of breast tumorigenesis [44].
2.2. miRNAs in Epithelial-Mesenchymal Transition, Invasion and Migration
EMT is a crucial contributary factor in cancer metastasis, in which cells lose their epithelial properties and acquire a morphology appropriate for invasion and migration [47,48]. Mesenchymal-to-epithelial transition (MET), the reverse process of EMT, is associated with metastatic colonisation in a distant site [49,50]. Epithelial markers include E-cadherin, cytokeratin and claudin-1 while mesenchymal markers include fibronectin, N-cadherin, SNAIL, SLUG, ZEB1, TWIST and vimentin [51,52]. The loss of E-cadherin expression, which induces the formation of cell-cell contact and adherens junctions, is heavily involved in EMT [53]. Repression of E-cadherin expression is mediated by multiple transcription factors such as SNAIL, ZEB 1/2 and SLUG via changes in several pathways, including TGF-β, Wnt and NOTCH [48,54,55]. Furthermore, recent studies have highlighted a link between EMT and cancer stem cells [56,57], a small subpopulation of tumour cells with self-renewal, differentiation and tumorigenicity properties when transplanted into another animal host [58]. EMT endows stem-like properties to cancer cells, while cancer stem cells commonly exhibit EMT properties [59]. Understanding the miRNAs which tightly regulate the EMT/MET pathway in cancer metastasis is thus ideal in developing targets for breast cancer therapy.
Multiple miRNAs have been identified to participate in EMT induction (Table 2).
LZTFL1 acts as a tumour suppressor which regulates β-catenin signalling in a number of cancers [79,80], consequently activating EMT. Down-regulation of miR-21 was found to inhibit EMT-mediated metastasis of breast cancer in vitro and in vivo by promoting LZTFL1 expression via the miR-21/LZTFL1/β-catenin axis [60]. Interestingly, miR-21 was also found to regulate β-catenin signalling via activation of the Akt/β-catenin pathway through PTEN, inducing subsequent EMT [60]. Besides its role in angiogenesis, miR-9 has also been linked to the promotion of EMT. miR-9 directly supresses CDH1 and thus E-cadherin, thereby leading to increased cell motility and invasiveness [40]. This was supported by another study showing that miR-9 expression in breast tumours is associated with E-cadherin loss and vimentin expression, thus playing a probable role in EMT in breast cancer [81]. Another key miRNA shown to augment migration, invasion and metastasis in breast cancer is miR-10b. An early study indicated that TWIST-induced miR-10b expression represses homeobox D10 mRNA translation, thereby increasing pro-metastatic RHoC gene expression with tumour invasion and metastasis [61]. This was supported by subsequent studies demonstrating development of breast cancer brain metastasis with upregulation of miR-10b [82] and EMT induction in breast cancer by miR-10b, which was found to act as a target gene of TGF-ß1 [83]. It was further shown that transfection of non-malignant mammary gland epithelial cells with exosomal-derived miR-10b induced cell invasion [35].
In addition, miR-221/222 has been linked to the aggressive basal-like subtype of breast cancer through its activation of EMT [84]. miR-221/222 acts downstream of the oncogenic Ras-Raf-MEK-ERK pathway, increasing ZEB2 levels and consequently repressing E-cadherin by targeting the 3’UTR of the GATA family transcriptional repressor TRPS1, resulting in heightened EMT in basal-like breast cancer [62]. miR-374a was also found to aid in the development of a pro-metastatic phenotype of breast cancer cells in vitro via induction of EMT, with overexpression of miR-374a resulting in cell morphologies characteristic of EMT. In miR-374a-transduced breast cancer cell lines, epithelial markers including E-cadherin, γ-catenin, and CK18 were drastically downregulated, while mesenchymal markers such as vimentin and N-cadherin were upregulated. Furthermore, miR-374a was found to interfere with EMT via Wnt/β-catenin signalling by directly suppressing WIF1, PTEN, and WNT5A expression in breast cancer cell lines [63]. Hypoxia-induced upregulation of miR-191 was also demonstrated to enhance breast cancer cell proliferation, migration and survival by increasing levels of TGFß2 and downstream proteins including VEGFA both directly and indirectly [64].
However, the roles of miR-125b, miR-155 and the miR-200 family in breast cancer invasion and EMT are less clearly delineated. The tumour suppressor gene STARD13 was identified as a target protein of miR-125b. Repression of STARD13 by miR-125b in MCF-7 and MDA-MB-231 cells was responsible for EMT and metastasis in breast cancer cell lines via upregulation of vimentin and α-smooth muscle actin [67]. On the contrary, miR-125b has been found to target SNAIL-1, with knockdown of miR-125b and consequent overexpression of SNAIL-1 increasing migration, invasion, and EMT in SKBR3-TR and BT474-TR cells [65]. miR-125b was further found to reverse motility, invasion and EMT in MCF-7 and SKBR3 paclitaxel-resistant breast cancer cells by targeting SEMA4C [66], an oncogenic protein demonstrated to upregulate SNAIL and SLUG [85]. This difference in observations could possibly be reconciled by the use of different breast cancer cell lines—while miR-125b expression is reported to be repressed in most breast cancer cell lines, it appears to be elevated in MDA-MB-231 cells [67,86].
Separately, miR-155 was found to target and repress C/EBPβ, thereby potentiating TGF-ß-mediated EMT [70]. It was further discovered that mutant p53-mediated upregulation of miR-155 and silencing of its target gene ZNF652, which directly represses key drivers of invasion and metastasis, drives local invasion of breast cancers [71]. However, seemingly contradictory reports have been made. miR-155 was reported to inhibit lung metastasis from mammary fat pads by preventing EMT through suppression of TCF4 expression [68]. More recently, miR-155 was also shown to downregulate ZEB2, with consequent reduced expression of vimentin and reduced invasion. Yet, ZEB2 repression did not change E-cadherin levels, and migration was instead enhanced [69]. Overall, it is suggested that miR-155 plays a larger pro-metastatic role in breast cancer—besides its roles in EMT, it has also been demonstrated to act as an oncogenic miRNA in angiogenesis and the Warburg effect as discussed in this review.
The miR-200 family has also been found to act as either tumour suppressors or oncogenic miRNAs at different junctures in the metastatic cascade. Several studies suggest that miR-200 miRNAs are downregulated and released from cancer cells during invasion as they undergo EMT, but upregulated during colonization. These miRNAs exist in two clusters: one on chromosome 1 (miR-200b, miR-200a and miR-429) and the other on chromosome 12 (miR-200c and miR-141) [87]. The miR-200 miRNAs repress invasion and EMT by targeting ZEB1 and ZEB2, repressors of the cell-cell contact protein E-cadherin [74,75,76], and regulating genes involved with cell motility and invasion [77,88]. In particular, miR-200c directly targets actin-regulatory proteins FHOD1 and PPM1F, inhibiting cancer migration and invasion through regulation of stress fibre formation and contractility [73]. Moreover, miR-200b was found to mediate many pathways including those in axonal guidance, chemokine, epithelial adherens junction and actin cytoskeleton signalling [72]. On the other hand, it has been discovered that the expression of miR-200 miRNAs is upregulated in metastases [89,90] and its levels are also elevated in the circulation of breast cancer patients with brain metastases [91]. Uptake of extracellular vesicles containing miR-200 was found to promote MET, thereby enabling the formation of adherent cellular contacts during colonization [78]. Hence the miR-200 family is a “double-edged sword” that can act as tumour suppressor at the beginning, but as an oncogenic factor at the end of the metastatic cascade.
2.3. miRNAs in the Warburg Effect
First discovered in the 1920s by Otto Heinrich Warburg, the Warburg effect and its implications on tumorigenesis and cancer progression have been studied extensively [92]. The Warburg effect is characterized by the preferential metabolism of glucose via aerobic glycolysis over oxidative phosphorylation in cancer cells [93]. This process supports macromolecular synthesis by providing an abundant supply of glycolysis intermediates and promotes cancer cell proliferation [94]. Emerging literature has demonstrated the involvement of miRNAs in the regulation of cellular metabolism and the Warburg effect in breast cancer (Table 3).
Various miRNAs have been found to display oncogenic effects in pathways involved in the Warburg effect. Notably, miR-155 has been found to play an integral role in regulating various pathways of aerobic glycolysis in breast cancer [101]. miR-155 represses cMyc, a master regulator of glycolysis [102], through the PIK3R1-PDK1/Akt-FOXO3a pathway. This results in the upregulation of glucose transporters and metabolic enzymes including GLUT1, HK2, PKM2 and LDHA [95]. The study corroborates earlier research showing that FOXO3a is a direct target of miR-155 [96] and deregulates cMyc [103]. In particular, HK2 has been credited as a crucial player in the Warburg effect as it is one of the chief isozymes overexpressed in tumours that promotes aerobic glycolysis [104]. In addition to the PIK3R1-PDK1/Akt-FOXO3a pathway, further studies show that miR-155 also regulates HK2 expression in breast cancer cells in two other ways. miR-155 does so by (i) promoting the activation of STAT3, which in turn facilitates the transcription of HK2, and (ii) repressing miR-143 by targeting C/EBPβ, a transcriptional activator for miR-143, resulting in post-transcriptional upregulation of HK2 [97]. These studies have lent credence to the central role played by miR-155 in the Warburg effect in breast cancer.
Another miRNA responsible for potentiating the Warburg effect in breast cancer is miR-27b. PDH is found at the crossroads of glycolysis and the citric acid cycle, thus a loss of function or reduced expression of PDH complex components has been associated with deregulation of glucose metabolism in cancer [105]. miR-27b has been found to directly reduce the expression of PDHX, a structural component of the PDH complex. This in turn suppresses oxidative glucose metabolism and facilitates tumour growth while increasing lactate production, a distinctive characteristic of Warburg metabolism [98]. miR-378* has also been found to mediate metabolic shift in breast cancer cells by inhibiting the expression of ERRγ and GABPA, partners of PGC-1ß, a transcriptional regulator of oxidative energy metabolism [106]. Downstream effects include a reduction in tricarboxylic acid cycle gene expression and an increase in lactate production and cell proliferation, with miR-378* expression demonstrating a correlation with breast cancer progression [99].
2.4. miRNAs in the Tumour Microenvironment
Besides modulating the nature of breast cancer cells directly, miRNAs have been found to play an important role in the tumour microenvironment (TME). It has been suggested that the outcomes of tumour cells are largely dependent on their interaction with stromal cells; they may then either remain dormant, or progress into invasive and eventually metastatic cancer [107]. Multiple cell populations regulate the TME through intriguing mechanisms including the secretion of pro-inflammatory molecules and immune regulation (Figure 3), which will be discussed below.
Cancer-associated fibroblasts (CAFs) are one of the most common cell types found in the TME, and are responsible for the synthesis of proteins which remodel the extracellular matrix and growth factors that regulate tumour cell proliferation, survival and metastasis [108,109]. CAFs release exosomal miRNAs (miR-21, miR-378e and miR-143), increasing the stemness and EMT phenotypes of breast cancer cells [110]. miR-181d-5p was also identified in exosomes derived from CAFs, and promoted EMT via targeting CDX2, a transcription factor that binds to the homeobox A5 promoter [111]. Furthermore, miR-125b released by TNBC cells in extracellular vesicles also promotes the conversion of NFs into CAFs [112]. Interestingly, this communication between CAFs and tumour cells goes both ways: tumour-derived miRNAs have also been shown to play a role in inducing the transformation of normal fibroblasts (NFs) into CAFs [113]. For instance, exosome-mediated delivery of miR-9 from breast cancer cells induces CAF-like properties in human breast fibroblasts through modulating the expression of various extracellular matrix proteins [114]. In addition, breast cancer cell-secreted miR-205 contributes to the conversion of breast NFs into CAFs by promoting YAP1 expression and subsequent tumour angiogenesis [115], and activates Myc signalling in CAFs to induce an optimal metabolic environment for sustained tumour growth [116].
Furthermore, changes in immune cells within the tumour microenvironment may lead to inhibition of the antitumour immune response and thus breast tumour progression, with miRNAs being crucial mediators of this process. For instance, miR-375, released by breast cancer cells during apoptosis, was found to accumulate in tumour-associated macrophages and enhance phagocyte migration and infiltration in vitro and in vivo, forming a tumour-promoting microenvironment [117]. MDSCs, shown to inhibit anti-tumour T cells, were activated by doxorubicin treatment and subsequently increased breast tumour angiogenesis and induced Th2 cell activation via exosomal release of miR-126a [118]. Prostaglandin E2-induced miR-10a production has also been shown to activate AMPK signalling, which in turn promotes the expansion and activation of MDSCs [119].
In addition, miRNAs were found to modulate the function of T cells, which are associated with cancer progression [120]. Silencing of miR-126 in a murine breast cancer model caused reduced induction and suppressive function of CD4+ FOXP3+ regulatory T cells (Tregs) through the PI3K/Akt pathway, and also endowed antitumour effects of CD8+ T cells [121]. Similarly, silencing of miR-21 regulated the PTEN/Akt pathway transduction in the expansion of CCR6+ Tregs and endowed the antitumour effects of CD8+ T cells in breast tumours [122].
3. miRNA-Based Therapies for Breast Cancer
With strong evidence supporting the role of miRNAs in breast cancer metastasis, much research into miRNA-based therapies has been conducted in recent years. miRNA delivery is centred around two main approaches: the delivery of (i) anti-miRNA oligonucleotides (anti-miRs) against oncogenic miRNAs, or (ii) tumour-suppressor miRNA mimetics, while other approaches to oncogenic miRNA inhibition have been explored as well (Figure 4).
However, several concerns regarding these approaches have been raised. One overarching consideration would be the off-target effects of miRNA-based therapies. For one, the downstream effects of miRNAs are multiple and varied, as a single miRNA is capable of regulating transcriptional networks involving a multitude of gene transcripts [8]. In fact, most mammalian mRNAs have been shown to be conserved targets of miRNAs [123]. Thus, changing the miRNA expression levels could see diverse downstream effects aside from the intended outcome, making it challenging to avoid off-target effects [124]. Furthermore, present research on miRNA therapeutics utilising tumour suppressor miRNAs relies principally on synthetic miRNAs. This surfaces the concern of potential toxicity and immunogenicity due to the introduction of foreign genetic materials, and could contain artificial modifications that may affect the biochemical properties of the synthetic miRNAs [125]. Similarly, the delivery vehicles used in miRNA-based therapies could also result in toxicity and immunogenicity [126]. To circumvent these limitations in miRNA delivery, multiple delivery platforms have been studied with the aim of achieving a desirable balance between efficacious miRNA delivery and reduced vehicular toxicity. In the following sections, we explore the various approaches taken in miRNA-based therapies and the associated challenges for each approach.
3.1. miRNA Delivery
3.1.1. Liposomes
Liposomes synthesized in the nanometer-size range (<250 nm in diameter) have been increasingly utilised in cancer drug delivery, and are associated with improved pharmacokinetic properties [127,128]. Achieving a good balance between size, payload concentration, drug solubility, protection from enzymatic degradation and systemic clearance has previously been emphasized in various reports [129,130]. Surface modification of liposomes can further improve the delivery of therapeutic miRNAs and antisense miRNAs against various cancers [131]. In their study, Sharma et al. developed a stearylamine based cationic liposome for the delivery of anti-miR-191 to MCF-7 and ZR-75-1 breast cancer cells. These liposomes showed efficient delivery to cancer cells with low cytotoxicity in human erythrocytes, as well as increased cancer cell apoptosis and suppressed cell migration in vitro. Furthermore, the liposomes also increased the chemosensitivity of the breast cancer cells to doxorubicin and cisplatin [132]. Recently, Lujan et al. has also described the optimization and synthesis of nanometer-sized liposomes for miRNA delivery, with miR-203 delivery to MDA-MD-231 breast cancer cells enhanced by up to 40-fold [133]. However, liposomes have limited capacity for in vivo delivery due to their rapid clearance as well as concerns over their toxicity, nonspecific uptake and immunogenicity [134].
3.1.2. Inorganic Nanoparticles
Extensive research has been conducted on superparamagnetic iron oxide nanoparticles (SPIONs) for their use in biotherapeutic delivery systems [135], as they have been demonstrated to possess biocompatible and non-toxic profiles at lower therapeutic levels [136]. Separately, argonaute proteins have been found to stabilize and guide mature miRNAs to their target messenger RNAs [137,138]. By capitalizing on the special properties of SPIONs and argonaute proteins, Unal et al. designed Argonaute 2 conjugated SPIONs as tumour targeted miRNA vehicles to deliver autophagy-inhibiting miR-376b into HER2-positive breast cancer cell lines. Effective inhibition of autophagic activity by the nanoparticles was demonstrated both in vitro and in vivo in a mice xenograft model of breast cancer [139]. However, concerns over iron oxide nanoparticle-induced toxicity still remain. Toxicity of SPIONs has been shown to be dependent on exposure time and concentration: while minimal toxicity is observed at lower levels of SPIONs with good body clearance, high dose exposure to SPIONs could trigger oxidative stress and altered cellular response [140]. Thus, achieving good therapeutic efficacy with minimal toxicity is a challenge that has to be overcome before this strategy can reach clinical trials.
Using a different approach, one study reported the development of multifunctional tumour-penetrating mesoporous silica nanoparticles (MSNs) for the co-delivery of siRNA (siPlk1) and a tumour suppressor miRNA (miR-200c) to breast tumours. Previously, it was shown that iRGD, a tumour-homing and penetrating peptide, increases accumulation and penetration of anticancer drugs and nanoparticles into tumours via a three-step endocytotic transport pathway [141,142]. In addition, it has also been demonstrated that light-activated generation of ROS disrupts endosomal and lysosomal membranes, thus facilitating cytoplasmic delivery of small RNAs [143,144]. In this study, MSNs were stabilized by a surface lipid layer conjugated to iRGD. MSNs were then loaded with photosensitizer indocyanine green which generated ROS to aid endosomal escape and surface conjugation of iRGD for enhanced cytosolic RNA delivery. Upon short light irradiation, the iRGD-modified MSNs loaded with siPlk1 and miR-200c showed improved delivery, cellular uptake and tumour penetration in vitro, and significant suppression of primary tumour growth with reduction of metastasis in vivo [145]. Yet, as with most cationic nanoparticle carriers, poor elimination, immunogenicity and toxicity remain as chief concerns for MSNs [146].
Gold nanoparticles (AuNPs) have also been widely used in miRNA delivery systems. Their unique properties allow them to have low cytotoxicity, good biodistribution, tunable size and functional diversity [147]. Capitalising on these properties, Ekin et al. designed a AuNP-based nanocarrier for miR-145 transfection into prostate and breast cancer cells. Since AuNPs have a high affinity for biomolecules and can be chemically functionalised with alkyl-thiol-terminated oligonucleotides [148], the study chose to modify AuNPs with thiolated RNAs to which pre-miR-145 could then be hybridised. Effective in vitro delivery of miR-145 into MCF7 breast cancer cells was demonstrated [149]. More recently, Ramchandani et al. devised a layer-by-layer fabrication method to layer negatively charged miR-708 mimetics between positively charged PLL layers onto an inert AuNP. Subsequent degradation of the PLL layers by proteases upregulated in tumours released the miR-708 mimetics, restoring tumour suppressive miR-708 and inhibiting TNBC metastasis in vivo [150]. While promising, toxicity-related concerns over AuNPs have surfaced depending on their size [151] and ability to cross the blood-brain barrier to accumulate in neural tissue [152].
3.1.3. Polymer-Based Delivery Systems
Given the associated toxicity with inorganic nanoparticles, cationic polymers have been explored as an alternative delivery platform. While concerns over the significant toxicity of first-generation carriers such as polyethylenimine (PEI) have surfaced, especially for the high-molecular weight forms [153], there is promise that this may be overcome by the introduction of biodegradable polymers in gene therapy [154]. PEIs are cationic linear or branched polymers that are able to form nanoscale complexes with small RNAs, reducing RNA degradation and increasing cellular delivery and intracellular release. They have thus been used extensively in RNA interference and gene delivery systems as an alternative to viral vectors [155]. Recent developments in PEI-based systems include disulfide cross-linked PEIs (PEI-SS) for miRNA delivery. PEI-SS was found to complex efficiently with anti-miR-155, forming nano-sized spherical structures. Subsequent biodegradation by the reducing agent glutathione in cancer cells released anti-miR-155 for the inhibition of tumour growth in vivo [156]. Another study developed a PLL-modified PEI (PEI-PLL) copolymer to transfect either miR-21 sponge plasmid DNA or anti-miR-21 oligonucleotides into MCF-7 breast cancer cells. The treated cells displayed greater miR-21 inhibition with cell cycle arrest in the G1 phase, as well as upregulation of PDCD4 involved in the caspase-3 apoptosis pathway. Furthermore, both groups of cells also showed increased sensitisation to anti-cancer drugs doxorubicin and cisplatin [157].
Poly(D,L-lactide-co-glycolide) (PLGA) is a synthetic polymer which has also been studied widely in anticancer drug delivery platforms due to its extensive functionalisation options, biodegradability, sustained-release efficacy, and stabilisation of loaded molecules [158]. Combining PLGA with PEI, Wang et al. devised a hyaluronic acid-decorated PEI-PLGA (HA/PEI-PLGA) nanoparticle system for the co-delivery of doxorubicin and miR-542-3p in TNBC therapy. Increased intracellular levels of miR-542-3p activated p53, thus promoting TNBC cell apoptosis and tumour suppression [159]. Polyethylene glycol (PEG), another biocompatible polymer, was used in combination with PGLA to form biodegradable PLGA-bPEG copolymers for the delivery of anti-miR-21 and anti-miR-10b to TNBC tumours. In vitro models displayed significant reduction in cell migration in treated cells, while in vivo models showed substantial reduction in tumour growth at low doses [160].
Another cationic polymer used in gene delivery systems is chitosan, which is biodegradable and biocompatible, and has strong nucleic acid binding affinity [161]. Several studies have established that chitosan-oligonucleotide complexes show low cytotoxicity [162,163], however, it was also reported that chitosans with higher degrees of acetylation and molecular weight may be cytotoxic [164]. In one study, chitosan was acetylated to varying degrees and used in the formation of chitosan-hsa-miR-145 (CS-miRNA) nanocomplexes, with chitosans of low degrees of acetylation forming highly stable complexes regardless of molecular weight. CS-miRNA nanocomplexes at 12% and 29% degrees of acetylation were biologically active, displaying downregulation of miR-145 target mRNA (junction adhesion molecule A mRNA) in MCF-7 breast cancer cells [165]. In addition, co-encapsulation of doxorubicin and miR-34a into hyaluronic acid-chitosan nanoparticles for simultaneous delivery into breast cancer cells was performed in another study. Enhanced anti-tumour effects of doxorubicin by suppressing the expression of non-pump resistance and anti-apoptosis proto-oncogene Bcl-2 was observed, along with the inhibition of breast cancer cell migration and metastasis by miR-34a via NOTCH-1 signalling [166].
In an unprecedented study, Conde et al. developed a self-assembling RNA-triple-helix assembly for miRNA delivery. This helix was conjugated to dendrimers and reacted with dextran aldehyde, forming an adhesive dextran-dendrimer-RNA triplex hydrogel scaffold which was able to adhere to tumour tissue and administer its miRNAs. The helix conjugate, comprising miRNA nucleotides miR-205 sense, antisense and antagomir-221, showed high structural stability and synergistic abrogation of TNBC tumours via a dual-pronged miRNA inhibition and miRNA replacement approach. Cell migration and proliferation were dramatically reduced, and nearly 90% levels of tumour shrinkage was achieved in a TNBC mouse model [167]. These results show great promise for the use of the RNA-triple-helix hydrogel in breast cancer therapy.
3.1.4. Extracellular Vesicles
Another strategy to reduce vehicle-associated toxicity is the use of extracellular vesicles in gene therapy. Extracellular vesicles such as exosomes are secreted by various cell types including tumour cells, and function as natural carriers of miRNAs. These miRNAs are then taken up by recipient cells where they elicit downstream responses [168,169]. This interesting phenomenon positions extracellular vesicles as suitable miRNA delivery vehicles in breast cancer therapy due to better biocompatibility and high delivery efficiency. For instance, a tumour cell-derived extracellular vesicle (TEV)-based nanoplatform was developed for the delivery of anti-miR-21 to 4T1 breast cancer cells. Subsequent functionalisation of gold-iron oxide nanoparticles (GIONs) in TEVs to yield TEV-GIONs demonstrated the potential of TEV-GIONs for simultaneous therapy using miRNA and cancer imaging, while improving tumour-specific targeting [170]. Separately, delivery of tumour suppressor let-7a miRNA using exosomes from HEK-293 cells to EGFR-expressing xenograft breast cancer tissue in mice was performed by Ohno et al., resulting in inhibition of tumour growth and development. This was achieved by fusing a GE11 peptide, which binds specifically to EGFR, to the transmembrane domain of PDGF on donor exosomes containing let-7a. However, the same study highlighted that accumulation of exosomes was observed in the liver after injection, surfacing potential bioelimination difficulties [171].
Exosomes were also used for the co-delivery of doxorubicin, a chemotherapy drug, and hydrophobically modified miR-159 in TNBC therapy. Exosomes highly expressing ADAM15 have been found to show an enhanced binding affinity for integrin αvβ3 [172], which is overexpressed on many tumours. Hydrophobically modifying a short RNA strand via addition of a cholesterol group enables quicker membrane association and facilitates internalisation [173]. In vitro targeting of MDA-MB-231 breast cancer cells showed greater internalisation of ADAM15-rich exosomes compared to the control exosomes, and consequently significantly higher cellular uptake of doxorubicin and modified miR-159, prompting increased apoptosis. In vivo studies also showed significant tumour suppression in a xenografted-nude mouse model by ADAM15-rich exosomes [174].
One drawback of using extracellular vesicles from cell lines for miRNA or anti-miR delivery is the risk of secondary transformation. Cell lines release multiple oncogenic factors into the extracellular vesicles, thus inducing transformation of bystander benign cells in target organs. Delivery of miRNA or anti-miR oligos using extracellular vesicles from primary cells is a safer alternative, however, it is challenging to obtain sufficient extracellular vesicles from primary cells for therapeutic treatments. To overcome these limitations, a recent study by Usman et al. described the use of red blood cell-derived extracellular vesicles to deliver anti-miR-125b for an effective treatment of breast cancer and leukemia in vitro and in vivo. This approach is compelling because red blood cells are the most abundant primary cells in the body and treatment of red blood cells with calcium ionophore induces a massive release of EVs, making this an ideal approach for clinical applications [175].
3.2. Other Approaches to Oncogenic miRNA Inhibition
3.2.1. Synthetic miRNA Sponges
First introduced in 2007, miRNA sponges contain multiple target sites complementary to a mature miRNA of interest, and are able to inhibit the activity of a family of miRNAs sharing a common seed (nucleotides 2-7 on the miRNA) [176]. The concept of miRNA sponges is a simple yet ingenious one, with researchers behind the first study reasoning that multiple binding sites could be inserted into the 3′ UTR of a decoy target to improve its affinity for its cognate miRNA. Furthermore, by designing the miRNA binding sites with a bulge at the position normally cleaved by Argonaute 2, the decoy targets would be able to stably hold on to ribonucleoprotein complexes containing numerous miRNAs [177]. Since then, multiple endogenous miRNA sponges regulating miRNA levels in breast cancer have been identified as well [178,179]. Using this concept, an miR-9 sponge construct introduced into highly malignant cells using a retroviral vector showed inhibition of metastasis formation [40]. Similar results were obtained for an miR-21 sponge, which demonstrated marked reduction in the expression of downstream CSF1, a potent activator of malignancy and metastasis [180]. More recently, Zhang et al. developed a self-assembled DNA nanosponge for the clearance of intracellular miR-21. MCF-7 breast cancer cells transfected with the nanosponge displayed increased apoptotic-related protein expression, while normal cells were minimally affected [181].
3.2.2. Non-Conventional Approaches to miRNA Inhibition
Recently, novel approaches to the indirect inhibition or suppression of oncogenic miRNAs in breast cancer have been explored. In one instance, photodynamic therapy, in which ROS are produced to damage cancer cells, was used to regulate miR-155-5p expression and the Warburg effect in breast cancer. The use of 3B, a novel photosensitizer, in photodynamic therapy demonstrated impaired glucose consumption and ATP generation, inhibition of miR-155-5p expression in MCF-7 cells and decreased tumour growth in vivo [182]. In another innovative approach taken by Costales et al., a small molecule that targets the three-dimensional folds in pre-miR-21 was designed and optimised for avidity, and its target engagement of pre-miR-21 was demonstrated in MDA-MB-231 cells. Subsequent conjugation of the small molecule to a heterocyclic molecule that recruits latent ribonuclease to cleave pre-miR-21 was performed, and a drastic reduction in miR-21 levels was observed. Furthermore, this conjugated small molecule effectively inhibited invasion in multiple miR-21-expressing cancer cell lines, and inhibited breast cancer metastasis to the lung in vivo [183].
4. Conclusions and Future Perspectives
To conclude, miRNAs are shown to be key regulators of metastasis in breast cancer. A growing pool of studies has demonstrated the huge potential for miRNA-based therapies in breast cancer, with multiple novel approaches suggested to overcome barriers including easy degradation of RNA molecules and non-specific and off-target delivery. However, as with other nucleotide-based therapeutic approaches, concerns persist over the possibility of toxicity and immunogenicity due to the introduction of foreign genetic materials and delivery vehicles, insufficient therapeutic efficacy, off-target effects and the feasibility of upscaling production for eventual clinical applications [184]. These concerns remain particularly valid, with the phase 1 study of MRX34, a liposomal miR-34a mimic, closed early due to serious immune-mediated adverse effects that resulted in four patient deaths [185]. Furthermore, the complex regulation of multiple pathways by miRNAs may make it challenging to clearly delineate the boundary between their effects on normal versus cancer cells.
To address these concerns, several novel approaches have been developed. Instead of using synthetic miRNAs, Wang et al. suggests the use of a miRNA prodrug which can be bioengineered on a large scale in Escherichia coli, using a recombinant tRNA fusion pre-miR-34a [125]. Using a similar method, another team created an miR-127 prodrug, which was processed to mature miR-127-3p in TNBC cells and demonstrated suppressed primary tumour growth and spontaneous metastasis in vivo [186]. Separately, concerns over delivery-associated toxicity may be addressed using vehicle-free delivery systems. Orellana et al. developed a strategy to conjugate a miR-34a with folate for delivery in breast tumours [187]. Stability of the miRNA mimic was achieved by modifying the passenger miRNA strand with 2′-O-methyl RNA bases to increase nuclease resistance without impairing argonaute loading. As the folate receptor is overexpressed in breast cancers [188], this approach of miRNA delivery showed selective targeting of the breast tumour with slowing of tumour growth, offering an alternative to current methods of vehicle-based miRNA delivery. Further enhancements including sugar modifications and backbone modifications that improve cellular uptake and binding specificity of miRNAs have also been explored [189,190]. Other promising areas in RNA interference include the combination of miRNAs with chemotherapy drugs, which could show synergistic effects in breast cancer therapy [191]. Furthermore, chemical modifications to guide strand selection and delivery to reduce off-target activity could also be performed [124].
Thus, while challenges remain ahead of adoption in clinical trials, miRNA-based approaches for breast cancer therapy are definitely promising. The twin pillars of any potential miRNA-based candidates—efficacy and safety—should be rigorously validated to bring their full benefit to patients.
Author Contributions
A.Y.T.T. is the lead study contributor for this review article. L.W. contributed to the structure design and to supervision. X.X., M.T.N.L., A.L.-A.W., Q.Z., L.W. and B.-C.G. contributed to critically revising the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Boon-Cher Goh is supported by the National Medical Research Council, Singapore (NMRC/CSA-SI/0006/2016 and NMRC/CG/M005/2017_NCIS). Lingzhi Wang is supported by Joint NCIS and NUS Cancer Program Seed Funding Grants, National University of Singapore. Andrea L.-A. Wong is supported by the National Medical Research Council, Singapore (NMRC/TA17nov003).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 3′UTR | 3′ untranslated region |
| ADAM15 | A disintegrin and metalloproteinase 15 |
| Akt | Protein kinase B |
| AMPK | 5’ AMP-activated protein kinase |
| AuNP | Gold nanoparticle |
| Bcl-2 | B-cell lymphoma 2 |
| C/EBPβ | CCAAT-enhancer binding protein beta |
| CAF | Cancer-associated fibroblast |
| CCR6 | Chemokine receptor 6 |
| CD4 | Cluster of differentiation 4 |
| CD8 | Cluster of differentiation 8 |
| CDH1 | Cadherin 1 |
| CSF1 | Colony-stimulating factor 1 |
| ECM | Extracellular matrix |
| EGFR | Epidermal growth factor receptor |
| EMT | Epithelial-mesenchymal transition |
| ERK | Extracellular signal-regulated kinases |
| ERRγ | Estrogen-related receptor gamma |
| FHOD1 | FH1/FH2 domain-containing protein 1 |
| FOXO3a | Forkhead box O3 |
| FOXP3 | Forkhead box P3 |
| GABPA | GA-binding protein alpha chain |
| GION | gold-iron oxide nanoparticles |
| GLUT1 | Glucose transporter 1 |
| HER2 | Human epidermal growth factor receptor 2 |
| HIF1α | Hypoxia-inducible factor 1-alpha |
| HK2 | Hexokinase 2 |
| iRGD | 9-amino acid cyclic peptide (sequence: CRGDKGPDC) |
| LATS2 | Large tumor suppressor kinase 2 |
| LDHA | Lactate dehydrogenase A |
| lncRNA | Long non-coding RNA |
| LZTFL1 | Leucine zipper transcription factor-like 1 |
| MDSC | Myeloid-derived suppressor cell |
| MEK | Mitogen-activated protein kinase |
| miRNA | microRNA |
| mRNA | Messenger RNA |
| MSN | Mesoporous silica nanoparticle |
| NF | Normal fibroblast |
| NOTCH-1 | Notch homolog 1 |
| PDCD4 | Programmed cell death protein 4 |
| PDGF | Platelet derived growth factor |
| PDH | Pyruvate dehydrogenase |
| PDHX | Pyruvate dehydrogenase protein X |
| PDK1 | Phosphoinositide-dependent kinase 1 |
| PEG | Polyethylene glycol |
| PEI | Polyethylenimine |
| PGC-1ß | (PPAR)γ coactivator-1ß |
| PI3K | Phosphoinositide 3-kinase |
| PIK3R1 | PI3K regulatory subunit alpha |
| PKM2 | pyruvate kinase isoenzyme type M2 |
| PLGA | Poly(d,l-lactide-co-glycolide) |
| PLL | Poly-L-lysine |
| PPM1F | Protein Phosphatase, Mg2+/Mn2+ Dependent 1F |
| PTEN | Phosphatase and tensin homolog deleted on chromosome 10 |
| RHoC | Ras homolog gene family, member C |
| RISC | RNA-induced silencing complex |
| ROS | Reactive oxygen species |
| SEC23a | Sec23 homolog A |
| SEMA4C | Semaphorin 4C |
| SLUG | Zinc finger protein SNAI2 |
| SNAIL | Zinc finger protein SNAI1 |
| SPION | Superparamagnetic iron oxide nanoparticle |
| STARD13 | StAR-related lipid transfer domain protein 13 |
| STAT3 | Signal transducer and activator of transcription 3 |
| TCF4 | Transcription factor 4 |
| TEV | Tumour cell-derived extracellular vesicles |
| TGF-ß | Transforming growth factor beta |
| TME | Tumour microenvironment |
| TNBC | Triple negative breast cancer |
| tRNA | Transfer RNA |
| TRPS1 | Tricho-rhino-phalangeal syndrome type 1 |
| TWIST | Twist-related protein 1 |
| VEGF | Vascular endothelial growth factor |
| VEGFR2 | Vascular endothelial growth factor receptor 2 |
| WIF1 | Wnt inhibitory factor 1 |
| WNT5A | Wnt family member 5A |
| YAP1 | Yes-associated protein 1 |
| ZEB1/2 | Zinc finger E-box-binding homeobox 1/2 |
| ZNF652 | Zinc finger protein 652 |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef]
- Gupta, G.P.; Massagué, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J. The Warburg metabolism fuels tumor metastasis. Cancer Metastasis Rev. 2019, 38, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Rofstad, E.K. Microenvironment-induced cancer metastasis. Int. J. Radiat. Biol. 2000, 76, 589–605. [Google Scholar] [CrossRef]
- Nicoloso, M.S.; Spizzo, R.; Shimizu, M.; Rossi, S.; Calin, G.A. MicroRNAs—The micro steering wheel of tumour metastases. Nat. Rev. Cancer 2009, 9, 293–302. [Google Scholar] [CrossRef]
- Lee, I.; Ajay, S.S.; Yook, J.I.; Kim, H.S.; Hong, S.H.; Kim, N.H.; Dhanasekaran, S.M.; Chinnaiyan, A.M.; Athey, B.D. New class of microRNA targets containing simultaneous 5′-UTR and 3′-UTR interaction sites. Genome Res. 2009, 19, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Imai-Sumida, M.; Tanaka, Y.; Dahiya, R. Interaction and cross-talk between non-coding RNAs. Cell. Mol. Life. Sci. 2018, 75, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Functional interactions among microRNAs and long noncoding RNAs. Semin. Cell Dev. Biol. 2014, 34, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [Green Version]
- Weidner, N.; Semple, J.P.; Welch, W.R.; Folkman, J. Tumor angiogenesis and metastasis—Correlation in invasive breast carcinoma. N. Engl. J. Med. 1991, 324, 1–8. [Google Scholar] [CrossRef]
- Bielenberg, D.R.; Zetter, B.R. The contribution of angiogenesis to the process of metastasis. Cancer J. 2015, 21, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J.A.; Brown, L.F.; Senger, D.R.; Lanir, N.; Van de Water, L.; Dvorak, A.M.; Dvorak, H.F. Pathogenesis of tumor stroma generation: A critical role for leaky blood vessels and fibrin deposition. Biochim. Biophys. Acta 1989, 948, 305–326. [Google Scholar] [CrossRef]
- Welch, D.R.; Hurst, D.R. Defining the hallmarks of metastasis. Cancer Res. 2019, 79, 3011–3027. [Google Scholar] [CrossRef]
- Friedl, P.; Locker, J.; Sahai, E.; Segall, J.E. Classifying collective cancer cell invasion. Nat. Cell Biol. 2012, 14, 777–783. [Google Scholar] [CrossRef]
- Price, J.T.; Thompson, E.W. Mechanisms of tumor invasion and metastasis: Emerging targets for therapy. Expert Opin. Ther. Targets 2002, 6, 217–233. [Google Scholar] [CrossRef]
- Iwatsuki, M.; Mimori, K.; Yokobori, T.; Ishi, H.; Beppu, T.; Nakamori, S.; Baba, H.; Mori, M. Epithelial-mesenchymal transition in cancer development and its clinical significance. Cancer Sci. 2010, 101, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Palma, M.; Biziato, D.; Petrova, T.V. Microenvironmental regulation of tumour angiogenesis. Nat. Rev. Cancer 2017, 17, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Condeelis, J.; Pollard, J.W. Macrophages: Obligate partners for tumor cell migration, invasion, and metastasis. Cell 2006, 124, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.G.; Vignjevic, D.M. Modes of cancer cell invasion and the role of the microenvironment. Curr. Opin. Cell Biol. 2015, 36, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Xu, J.; Lan, H. Tumor-associated macrophages in tumor metastasis: Biological roles and clinical therapeutic applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- Lu, J.; Tan, M.; Cai, Q. The Warburg effect in tumor progression: Mitochondrial oxidative metabolism as an anti-metastasis mechanism. Cancer Lett. 2015, 356, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or tumor suppressor? The duplicity of MicroRNAs in cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- King, H.W.; Michael, M.Z.; Gleadle, J.M. Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer 2012, 12, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Pochampally, R.; Watabe, K.; Lu, Z.; Mo, Y.Y. Exosome-mediated transfer of miR-10b promotes cell invasion in breast cancer. Mol. Cancer 2014, 13, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, N.; Yano, H.; Nishida, T.; Kamura, T.; Kojiro, M. Angiogenesis in cancer. Vasc. Health Risk Manag. 2006, 2, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Fingleton, B.; Rothenberg, M.L.; Matrisian, L.M. Matrix metalloproteinases: Biologic activity and clinical implications. J. Clin. Oncol. 2000, 18, 1135–1149. [Google Scholar] [CrossRef]
- Neufeld, G.; Cohen, T.; Gengrinovitch, S.; Poltorak, Z. Vascular endothelial growth factor (VEGF) and its receptors. FASEB J. 1999, 13, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Fagiani, E.; Christofori, G. Angiopoietins in angiogenesis. Cancer Lett. 2013, 328, 18–26. [Google Scholar] [CrossRef]
- Ma, L.; Young, J.; Prabhala, H.; Pan, E.; Mestdagh, P.; Muth, D.; Teruya-Feldstein, J.; Reinhardt, F.; Onder, T.T.; Valastyan, S.; et al. miR-9, a MYC/MYCN-activated microRNA, regulates E-cadherin and cancer metastasis. Nat. Cell. Biol. 2010, 12, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; He, L.; Richards, E.J.; Challa, S.; Xu, C.X.; Permuth-Wey, J.; Lancaster, J.M.; Coppola, D.; Sellers, T.A.; Djeu, J.Y.; et al. Upregulation of miRNA-155 promotes tumour angiogenesis by targeting VHL and is associated with poor prognosis and triple-negative breast cancer. Oncogene 2014, 33, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luengo-Gil, G.; Gonzalez-Billalabeitia, E.; Perez-Henarejos, S.A.; Navarro Manzano, E.; Chaves-Benito, A.; Garcia-Martinez, E.; Garcia-Garre, E.; Vicente, V.; Ayala de la Pena, F. Angiogenic role of miR-20a in breast cancer. PLoS ONE 2018, 13, e0194638. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Du, W.W.; Yang, W.; Rutnam, Z.J.; Peng, C.; Li, H.; O’Malley, Y.Q.; Askeland, R.W.; Sugg, S.; Liu, M.; et al. MiR-93 enhances angiogenesis and metastasis by targeting LATS2. Cell Cycle 2012, 11, 4352–4365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Tu, Y.; Wan, L.; Bu, L.; Huang, T.; Sun, X.; Wang, K.; Shen, B. In vivo monitoring of angiogenesis inhibition via down-regulation of mir-21 in a VEGFR2-luc murine breast cancer model using bioluminescent imaging. PLoS ONE 2013, 8, e71472. [Google Scholar] [CrossRef]
- Tanimoto, K.; Makino, Y.; Pereira, T.; Poellinger, L. Mechanism of regulation of the hypoxia-inducible factor-1 alpha by the von Hippel-Lindau tumor suppressor protein. EMBO J. 2000, 19, 4298–4309. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Pei, J.; Xia, H.; Ke, H.; Wang, H.; Tao, W. Lats2, a putative tumor suppressor, inhibits G1/S transition. Oncogene 2003, 22, 4398–4405. [Google Scholar] [CrossRef] [Green Version]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar] [CrossRef]
- Gonzalez, D.M.; Medici, D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci. Signal. 2014, 7, re8. [Google Scholar] [CrossRef] [Green Version]
- Chao, Y.L.; Shepard, C.R.; Wells, A. Breast carcinoma cells re-express E-cadherin during mesenchymal to epithelial reverting transition. Mol. Cancer 2010, 9, 179. [Google Scholar] [CrossRef] [Green Version]
- Gunasinghe, N.P.; Wells, A.; Thompson, E.W.; Hugo, H.J. Mesenchymal-epithelial transition (MET) as a mechanism for metastatic colonization in breast cancer. Cancer Metastasis Rev. 2012, 31, 469–478. [Google Scholar] [CrossRef]
- Zeisberg, M.; Neilson, E.G. Biomarkers for epithelial-mesenchymal transitions. J. Clin. Investig. 2009, 119, 1429–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voulgari, A.; Pintzas, A. Epithelial-mesenchymal transition in cancer metastasis: Mechanisms, markers and strategies to overcome drug resistance in the clinic. Biochim. Biophys. Acta 2009, 1796, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Graziano, F. The E-Cadherin gene, structure and function. In Spotlight on Familial and Hereditary Gastric Cancer; Springer: Cham, Switzerland, 2013; pp. 27–33. [Google Scholar]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell. Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felipe Lima, J.; Nofech-Mozes, S.; Bayani, J.; Bartlett, J.M. EMT in breast carcinoma—A review. J. Clin. Med. 2016, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Sato, R.; Semba, T.; Saya, H.; Arima, Y. Concise review: Stem cells and epithelial-mesenchymal transition in cancer: Biological implications and therapeutic targets. Stem Cells 2016, 34, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.S.; Jiang, J.; Liang, X.H.; Tang, Y.L. Links between cancer stem cells and epithelial-mesenchymal transition. Onco Targets Ther. 2015, 8, 2973–2980. [Google Scholar] [CrossRef] [Green Version]
- Clarke, M.F.; Hass, A.T. Cancer stem cells. Rev. Cell Biol. Mol. Med. 2006. [Google Scholar] [CrossRef]
- Liu, X.; Fan, D. The epithelial-mesenchymal transition and cancer stem cells: Functional and mechanistic links. Curr. Pharm. Des. 2015, 21, 1279–1291. [Google Scholar] [CrossRef]
- Wang, H.; Tan, Z.; Hu, H.; Liu, H.; Wu, T.; Zheng, C.; Wang, X.; Luo, Z.; Wang, J.; Liu, S.; et al. microRNA-21 promotes breast cancer proliferation and metastasis by targeting LZTFL1. BMC Cancer 2019, 19, 738. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature 2007, 449, 682–688. [Google Scholar] [CrossRef]
- Stinson, S.; Lackner, M.R.; Adai, A.T.; Yu, N.; Kim, H.J.; O’Brien, C.; Spoerke, J.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. TRPS1 targeting by miR-221/222 promotes the epithelial-to-mesenchymal transition in breast cancer. Sci. Signal. 2011, 4, ra41. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Guan, H.; Fang, L.; Yang, Y.; Zhu, X.; Yuan, J.; Wu, J.; Li, M. MicroRNA-374a activates Wnt/beta-catenin signaling to promote breast cancer metastasis. J. Clin. Investig. 2013, 123, 566–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagpal, N.; Ahmad, H.M.; Chameettachal, S.; Sundar, D.; Ghosh, S.; Kulshreshtha, R. HIF-inducible miR-191 promotes migration in breast cancer through complex regulation of TGFbeta-signaling in hypoxic microenvironment. Sci. Rep. 2015, 5, 9650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Hu, J.; Zou, K.; Ye, M.; Chen, Y.; Wu, C.; Chen, X.; Han, M. Activation of LncRNA TINCR by H3K27 acetylation promotes Trastuzumab resistance and epithelial-mesenchymal transition by targeting MicroRNA-125b in breast Cancer. Mol. Cancer 2019, 18, 3. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, Y.; Lu, X.; Zhao, Z.; Zhu, L.; Chen, S.; Wu, Q.; Chen, C.; Wang, Z. MiR-125b regulates epithelial-mesenchymal transition via targeting Sema4C in paclitaxel-resistant breast cancer cells. Oncotarget 2015, 6, 3268–3279. [Google Scholar] [CrossRef] [Green Version]
- Tang, F.; Zhang, R.; He, Y.; Zou, M.; Guo, L.; Xi, T. MicroRNA-125b induces metastasis by targeting STARD13 in MCF-7 and MDA-MB-231 breast cancer cells. PLoS ONE 2012, 7, e35435. [Google Scholar] [CrossRef]
- Xiang, X.; Zhuang, X.; Ju, S.; Zhang, S.; Jiang, H.; Mu, J.; Zhang, L.; Miller, D.; Grizzle, W.; Zhang, H.G. miR-155 promotes macroscopic tumor formation yet inhibits tumor dissemination from mammary fat pads to the lung by preventing EMT. Oncogene 2011, 30, 3440–3453. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.Y.; Dayan, S.; Wong, S.W.; Kaczmarek, A.; Hope, C.M.; Pederson, S.M.; Arnet, V.; Goodall, G.J.; Russell, D.; Sadlon, T.J.; et al. FOXP3 and miR-155 cooperate to control the invasive potential of human breast cancer cells by down regulating ZEB2 independently of ZEB1. Oncotarget 2018, 9, 27708–27727. [Google Scholar] [CrossRef]
- Johansson, J.; Berg, T.; Kurzejamska, E.; Pang, M.F.; Tabor, V.; Jansson, M.; Roswall, P.; Pietras, K.; Sund, M.; Religa, P.; et al. MiR-155-mediated loss of C/EBPbeta shifts the TGF-beta response from growth inhibition to epithelial-mesenchymal transition, invasion and metastasis in breast cancer. Oncogene 2013, 32, 5614–5624. [Google Scholar] [CrossRef] [Green Version]
- Neilsen, P.M.; Noll, J.E.; Mattiske, S.; Bracken, C.P.; Gregory, P.A.; Schulz, R.B.; Lim, S.P.; Kumar, R.; Suetani, R.J.; Goodall, G.J.; et al. Mutant p53 drives invasion in breast tumors through up-regulation of miR-155. Oncogene 2013, 32, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.V.; Martin, E.C.; Segar, H.C.; Miller, D.F.; Buechlein, A.; Rusch, D.B.; Nephew, K.P.; Burow, M.E.; Collins-Burow, B.M. Dual regulation by microRNA-200b-3p and microRNA-200b-5p in the inhibition of epithelial-to-mesenchymal transition in triple-negative breast cancer. Oncotarget 2015, 6, 16638–16652. [Google Scholar] [CrossRef] [PubMed]
- Jurmeister, S.; Baumann, M.; Balwierz, A.; Keklikoglou, I.; Ward, A.; Uhlmann, S.; Zhang, J.D.; Wiemann, S.; Sahin, O. MicroRNA-200c represses migration and invasion of breast cancer cells by targeting actin-regulatory proteins FHOD1 and PPM1F. Mol. Cell. Biol. 2012, 32, 633–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell. Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Park, S.M.; Gaur, A.B.; Lengyel, E.; Peter, M.E. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008, 22, 894–907. [Google Scholar] [CrossRef] [Green Version]
- Korpal, M.; Lee, E.S.; Hu, G.; Kang, Y. The miR-200 family inhibits epithelial-mesenchymal transition and cancer cell migration by direct targeting of E-cadherin transcriptional repressors ZEB1 and ZEB2. J. Biol. Chem. 2008, 283, 14910–14914. [Google Scholar] [CrossRef] [Green Version]
- Howe, E.N.; Cochrane, D.R.; Richer, J.K. Targets of miR-200c mediate suppression of cell motility and anoikis resistance. Breast Cancer Res. 2011, 13, R45. [Google Scholar] [CrossRef] [Green Version]
- Le, M.T.; Hamar, P.; Guo, C.; Basar, E.; Perdigao-Henriques, R.; Balaj, L.; Lieberman, J. miR-200-containing extracellular vesicles promote breast cancer cell metastasis. J. Clin. Investig. 2014, 124, 5109–5128. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Guo, J.; Wang, Q.; Zhou, J.; Xu, C.; Teng, R.; Chen, Y.; Wei, Q.; Liu, Z.P. LZTFL1 suppresses gastric cancer cell migration and invasion through regulating nuclear translocation of beta-catenin. J. Cancer Res. Clin. Oncol. 2014, 140, 1997–2008. [Google Scholar] [CrossRef]
- Wei, Q.; Chen, Z.H.; Wang, L.; Zhang, T.; Duan, L.; Behrens, C.; Wistuba, I.I.; Minna, J.D.; Gao, B.; Luo, J.H.; et al. LZTFL1 suppresses lung tumorigenesis by maintaining differentiation of lung epithelial cells. Oncogene 2016, 35, 2655–2663. [Google Scholar] [CrossRef] [Green Version]
- Gwak, J.M.; Kim, H.J.; Kim, E.J.; Chung, Y.R.; Yun, S.; Seo, A.N.; Lee, H.J.; Park, S.Y. MicroRNA-9 is associated with epithelial-mesenchymal transition, breast cancer stem cell phenotype, and tumor progression in breast cancer. Breast Cancer Res. Treat. 2014, 147, 39–49. [Google Scholar] [CrossRef]
- Ahmad, A.; Sethi, S.; Chen, W.; Ali-Fehmi, R.; Mittal, S.; Sarkar, F.H. Up-regulation of microRNA-10b is associated with the development of breast cancer brain metastasis. Am. J. Transl. Res. 2014, 6, 384–390. [Google Scholar] [PubMed]
- Han, X.; Yan, S.; Weijie, Z.; Feng, W.; Liuxing, W.; Mengquan, L.; Qingxia, F. Critical role of miR-10b in transforming growth factor-beta1-induced epithelial-mesenchymal transition in breast cancer. Cancer Gene Ther. 2014, 21, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.Y.; Calin, G.A. MicroRNAs miR-221 and miR-222: A new level of regulation in aggressive breast cancer. Genome Med. 2011, 3, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurrapu, S.; Pupo, E.; Franzolin, G.; Lanzetti, L.; Tamagnone, L. Sema4C/PlexinB2 signaling controls breast cancer cell growth, hormonal dependence and tumorigenic potential. Cell Death Differ. 2018, 25, 1259–1275. [Google Scholar] [CrossRef] [Green Version]
- Rajabi, H.; Jin, C.; Ahmad, R.; McClary, C.; Joshi, M.D.; Kufe, D. Mucin 1 oncoprotein expression is suppressed by the miR-125b oncomir. Genes Cancer 2010, 1, 62–68. [Google Scholar] [CrossRef]
- Saydam, O.; Shen, Y.; Wurdinger, T.; Senol, O.; Boke, E.; James, M.F.; Tannous, B.A.; Stemmer-Rachamimov, A.O.; Yi, M.; Stephens, R.M.; et al. Downregulated microRNA-200a in meningiomas promotes tumor growth by reducing E-cadherin and activating the Wnt/beta-catenin signaling pathway. Mol. Cell. Biol. 2009, 29, 5923–5940. [Google Scholar] [CrossRef] [Green Version]
- Sossey-Alaoui, K.; Bialkowska, K.; Plow, E.F. The miR200 family of microRNAs regulates WAVE3-dependent cancer cell invasion. J. Biol. Chem. 2009, 284, 33019–33029. [Google Scholar] [CrossRef] [Green Version]
- Dykxhoorn, D.M.; Wu, Y.; Xie, H.; Yu, F.; Lal, A.; Petrocca, F.; Martinvalet, D.; Song, E.; Lim, B.; Lieberman, J. miR-200 enhances mouse breast cancer cell colonization to form distant metastases. PLoS ONE 2009, 4, e7181. [Google Scholar] [CrossRef] [Green Version]
- Korpal, M.; Ell, B.J.; Buffa, F.M.; Ibrahim, T.; Blanco, M.A.; Celia-Terrassa, T.; Mercatali, L.; Khan, Z.; Goodarzi, H.; Hua, Y.; et al. Direct targeting of Sec23a by miR-200s influences cancer cell secretome and promotes metastatic colonization. Nat. Med. 2011, 17, 1101–1108. [Google Scholar] [CrossRef] [Green Version]
- Madhavan, D.; Zucknick, M.; Wallwiener, M.; Cuk, K.; Modugno, C.; Scharpff, M.; Schott, S.; Heil, J.; Turchinovich, A.; Yang, R.; et al. Circulating miRNAs as surrogate markers for circulating tumor cells and prognostic markers in metastatic breast cancer. Clin Cancer Res. 2012, 18, 5972–5982. [Google Scholar] [CrossRef] [Green Version]
- Liberti, M.V.; Locasale, J.W. The Warburg effect: How does it benefit cancer cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic glycolysis: Meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Lee, E.; Jung, J.; Lee, J.W.; Kim, H.J.; Kim, J.; Yoo, H.J.; Lee, H.J.; Chae, S.Y.; Jeon, S.M.; et al. microRNA-155 positively regulates glucose metabolism via PIK3R1-FOXO3a-cMYC axis in breast cancer. Oncogene 2018, 37, 2982–2991. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; He, L.; Coppola, M.; Guo, J.; Esposito, N.N.; Coppola, D.; Cheng, J.Q. MicroRNA-155 regulates cell survival, growth, and chemosensitivity by targeting FOXO3a in breast cancer. J. Biol. Chem. 2010, 285, 17869–17879. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Zhang, L.F.; Zhang, H.W.; Hu, S.; Lu, M.H.; Liang, S.; Li, B.; Li, Y.; Li, D.; Wang, E.D.; et al. A novel miR-155/miR-143 cascade controls glycolysis by regulating hexokinase 2 in breast cancer cells. EMBO J. 2012, 31, 1985–1998. [Google Scholar] [CrossRef] [Green Version]
- Eastlack, S.C.; Dong, S.; Ivan, C.; Alahari, S.K. Suppression of PDHX by microRNA-27b deregulates cell metabolism and promotes growth in breast cancer. Mol. Cancer 2018, 17, 100. [Google Scholar] [CrossRef]
- Eichner, L.J.; Perry, M.C.; Dufour, C.R.; Bertos, N.; Park, M.; St-Pierre, J.; Giguere, V. miR-378( *) mediates metabolic shift in breast cancer cells via the PGC-1beta/ERRgamma transcriptional pathway. Cell Metab. 2010, 12, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Krist, B.; Florczyk, U.; Pietraszek-Gremplewicz, K.; Jozkowicz, A.; Dulak, J. The Role of miR-378a in Metabolism, Angiogenesis, and Muscle Biology. Int. J. Endocrinol. 2015, 2015, 281756. [Google Scholar] [CrossRef] [Green Version]
- Xia, M.; Feng, S.; Chen, Z.; Wen, G.; Zu, X.; Zhong, J. Non-coding RNAs: Key regulators of aerobic glycolysis in breast cancer. Life Sci. 2020, 250, 117579. [Google Scholar] [CrossRef]
- Dang, C.V.; Le, A.; Gao, P. MYC-induced cancer cell energy metabolism and therapeutic opportunities. Clin. Cancer Res. 2009, 15, 6479–6483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferber, E.C.; Peck, B.; Delpuech, O.; Bell, G.P.; East, P.; Schulze, A. FOXO3a regulates reactive oxygen metabolism by inhibiting mitochondrial gene expression. Cell Death Differ. 2012, 19, 968–979. [Google Scholar] [CrossRef]
- Mathupala, S.P.; Ko, Y.H.; Pedersen, P.L. Hexokinase-2 bound to mitochondria: Cancer’s stygian link to the “Warburg Effect” and a pivotal target for effective therapy. Semin. Cancer Biol. 2009, 19, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFate, T.; Mohyeldin, A.; Lu, H.; Thakar, J.; Henriques, J.; Halim, N.D.; Wu, H.; Schell, M.J.; Tsang, T.M.; Teahan, O.; et al. Pyruvate dehydrogenase complex activity controls metabolic and malignant phenotype in cancer cells. J. Biol. Chem. 2008, 283, 22700–22708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Handschin, C.; Spiegelman, B.M. Metabolic control through the PGC-1 family of transcription coactivators. Cell. Metab. 2005, 1, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Valastyan, S.; Weinberg, R.A. Tumor metastasis: Molecular insights and evolving paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Saidou, J.; Watabe, K. Cancer associated fibroblasts (CAFs) in tumor microenvironment. Front. Biosci. 2010, 15, 166–179. [Google Scholar] [CrossRef] [Green Version]
- Schoepp, M.; Strose, A.J.; Haier, J. Dysregulation of miRNA expression in Cancer Associated Fibroblasts (CAFs) and its consequences on the tumor microenvironment. Cancers 2017, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Donnarumma, E.; Fiore, D.; Nappa, M.; Roscigno, G.; Adamo, A.; Iaboni, M.; Russo, V.; Affinito, A.; Puoti, I.; Quintavalle, C.; et al. Cancer-associated fibroblasts release exosomal microRNAs that dictate an aggressive phenotype in breast cancer. Oncotarget 2017, 8, 19592–19608. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wei, H.; Wang, J.; Li, L.; Chen, A.; Li, Z. MicroRNA-181d-5p-containing exosomes derived from CAFs promote EMT by regulating CDX2/HOXA5 in breast cancer. Mol. Ther. Nucleic Acids 2020, 19, 654–667. [Google Scholar] [CrossRef]
- Vu, L.T.; Peng, B.; Zhang, D.X.; Ma, V.; Mathey-Andrews, C.A.; Lam, C.K.; Kiomourtzis, T.; Jin, J.; McReynolds, L.; Huang, L.; et al. Tumor-secreted extracellular vesicles promote the activation of cancer-associated fibroblasts via the transfer of microRNA-125b. J. Extracell. Vesicles 2019, 8, 1599680. [Google Scholar] [CrossRef] [Green Version]
- Alkasalias, T.; Moyano-Galceran, L.; Arsenian-Henriksson, M.; Lehti, K. Fibroblasts in the tumor microenvironment: Shield or spear? Int. J. Mol. Sci. 2018, 19, 1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baroni, S.; Romero-Cordoba, S.; Plantamura, I.; Dugo, M.; D’Ippolito, E.; Cataldo, A.; Cosentino, G.; Angeloni, V.; Rossini, A.; Daidone, M.G.; et al. Exosome-mediated delivery of miR-9 induces cancer-associated fibroblast-like properties in human breast fibroblasts. Cell Death Dis. 2016, 7, e2312. [Google Scholar] [CrossRef]
- Du, Y.E.; Tu, G.; Yang, G.; Li, G.; Yang, D.; Lang, L.; Xi, L.; Sun, K.; Chen, Y.; Shu, K.; et al. MiR-205/YAP1 in activated fibroblasts of breast tumor promotes VEGF-independent angiogenesis through STAT3 signaling. Theranostics 2017, 7, 3972–3988. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wu, X.; Zhou, W.; Fong, M.Y.; Cao, M.; Liu, J.; Liu, X.; Chen, C.H.; Fadare, O.; Pizzo, D.P.; et al. Cancer-cell-secreted exosomal miR-105 promotes tumour growth through the MYC-dependent metabolic reprogramming of stromal cells. Nat. Cell. Biol. 2018, 20, 597–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, A.C.; Ebersberger, S.; Fink, A.F.; Lampe, S.; Weigert, A.; Schmid, T.; Ebersberger, I.; Syed, S.N.; Brune, B. Apoptotic tumor cell-derived microRNA-375 uses CD36 to alter the tumor-associated macrophage phenotype. Nat. Commun. 2019, 10, 1135. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Rong, Y.; Teng, Y.; Zhuang, X.; Samykutty, A.; Mu, J.; Zhang, L.; Cao, P.; Yan, J.; Miller, D.; et al. Exosomes miR-126a released from MDSC induced by DOX treatment promotes lung metastasis. Oncogene 2017, 36, 639–651. [Google Scholar] [CrossRef] [Green Version]
- Rong, Y.; Yuan, C.H.; Qu, Z.; Zhou, H.; Guan, Q.; Yang, N.; Leng, X.H.; Bu, L.; Wu, K.; Wang, F.B. Doxorubicin resistant cancer cells activate myeloid-derived suppressor cells by releasing PGE2. Sci. Rep. 2016, 6, 23824. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Cai, X.; Chen, X.; Liang, H.; Zhang, Y.; Li, J.; Wang, Z.; Chen, X.; Zhang, W.; Yokoyama, S.; et al. Tumor-secreted miR-214 induces regulatory T cells: A major link between immune evasion and tumor growth. Cell Res. 2014, 24, 1164–1180. [Google Scholar] [CrossRef]
- Qin, A.; Wen, Z.; Zhou, Y.; Li, Y.; Li, Y.; Luo, J.; Ren, T.; Xu, L. MicroRNA-126 regulates the induction and function of CD4(+) Foxp3(+) regulatory T cells through PI3K/AKT pathway. J. Cell. Mol. Med. 2013, 17, 252–264. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, C.; Li, Y.; Zhao, J.; Chen, C.; Zhou, Y.; Tao, Y.; Guo, M.; Qin, N.; Ren, T.; et al. MiR-21 controls in situ expansion of CCR6(+) regulatory T cells through PTEN/AKT pathway in breast cancer. Immunol. Cell Biol. 2015, 93, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoszewski, R.; Sikorski, A.F. Editorial focus: Understanding off-target effects as the key to successful RNAi therapy. Cell Mol. Biol. Lett. 2019, 24, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.P.; Ho, P.Y.; Chen, Q.X.; Addepalli, B.; Limbach, P.A.; Li, M.M.; Wu, W.J.; Jilek, J.L.; Qiu, J.X.; Zhang, H.J.; et al. Bioengineering novel chimeric microRNA-34a for prodrug cancer therapy: High-yield expression and purification, and structural and functional characterization. J. Pharmacol. Exp. Ther. 2015, 354, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, Z.; Gemeinhart, R.A. Progress in microRNA delivery. J. Control Release 2013, 172, 962–974. [Google Scholar] [CrossRef] [Green Version]
- Malam, Y.; Loizidou, M.; Seifalian, A.M. Liposomes and nanoparticles: Nanosized vehicles for drug delivery in cancer. Trends Pharm. Sci. 2009, 30, 592–599. [Google Scholar] [CrossRef]
- Miele, E.; Spinelli, G.P.; Miele, E.; Di Fabrizio, E.; Ferretti, E.; Tomao, S.; Gulino, A. Nanoparticle-based delivery of small interfering RNA: Challenges for cancer therapy. Int. J. Nanomed. 2012, 7, 3637–3657. [Google Scholar] [CrossRef] [Green Version]
- Moghimi, S.M.; Hunter, A.C.; Murray, J.C. Long-circulating and target-specific nanoparticles: Theory to practice. Pharmacol. Rev. 2001, 53, 283–318. [Google Scholar]
- Gabizon, A.; Catane, R.; Uziely, B.; Kaufman, B.; Safra, T.; Cohen, R.; Martin, F.; Huang, A.; Barenholz, Y. Prolonged circulation time and enhanced accumulation in malignant exudates of doxorubicin encapsulated in polyethylene-glycol coated liposomes. Cancer Res. 1994, 54, 987–992. [Google Scholar]
- Ganju, A.; Khan, S.; Hafeez, B.B.; Behrman, S.W.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. miRNA nanotherapeutics for cancer. Drug Discov. Today 2017, 22, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Rajendran, V.; Kulshreshtha, R.; Ghosh, P.C. Enhanced efficacy of anti-miR-191 delivery through stearylamine liposome formulation for the treatment of breast cancer cells. Int. J. Pharm. 2017, 530, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Lujan, H.; Griffin, W.C.; Taube, J.H.; Sayes, C.M. Synthesis and characterization of nanometer-sized liposomes for encapsulation and microRNA transfer to breast cancer cells. Int. J. Nanomed. 2019, 14, 5159–5173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, H.; Zhang, S.; Wang, B.; Cui, S.; Yan, J. Toxicity of cationic lipids and cationic polymers in gene delivery. J. Control Release 2006, 114, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Mok, H.; Zhang, M. Superparamagnetic iron oxide nanoparticle-based delivery systems for biotherapeutics. Expert Opin. Drug Deliv. 2013, 10, 73–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarockyte, G.; Daugelaite, E.; Stasys, M.; Statkute, U.; Poderys, V.; Tseng, T.C.; Hsu, S.H.; Karabanovas, V.; Rotomskis, R. Accumulation and toxicity of superparamagnetic iron oxide nanoparticles in cells and experimental animals. Int. J. Mol. Sci. 2016, 17, 1193. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.W.; Noland, C.; Siridechadilok, B.; Taylor, D.W.; Ma, E.; Felderer, K.; Doudna, J.A.; Nogales, E. Structural insights into RNA processing by the human RISC-loading complex. Nat. Struct. Mol. Biol. 2009, 16, 1148–1153. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.S.; Garchow, B.G.; Liu, X.; Yeung, J.; Morris, J.P.T.; Cuellar, T.L.; McManus, M.T.; Kiriakidou, M. Expanded RNA-binding activities of mammalian Argonaute 2. Nucleic Acids Res. 2009, 37, 7533–7545. [Google Scholar] [CrossRef] [Green Version]
- Unal, O.; Akkoc, Y.; Kocak, M.; Nalbat, E.; Dogan-Ekici, A.I.; Yagci Acar, H.; Gozuacik, D. Treatment of breast cancer with autophagy inhibitory microRNAs carried by AGO2-conjugated nanoparticles. J. Nanobiotechnol. 2020, 18, 65. [Google Scholar] [CrossRef]
- Patil, R.M.; Thorat, N.D.; Shete, P.B.; Bedge, P.A.; Gavde, S.; Joshi, M.G.; Tofail, S.A.M.; Bohara, R.A. Comprehensive cytotoxicity studies of superparamagnetic iron oxide nanoparticles. Biochem. Biophys. Rep. 2018, 13, 63–72. [Google Scholar] [CrossRef]
- Teesalu, T.; Sugahara, K.N.; Kotamraju, V.R.; Ruoslahti, E. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration. Proc. Natl. Acad. Sci. USA 2009, 106, 16157–16162. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xie, Y.; Li, J.; Peng, Z.H.; Sheinin, Y.; Zhou, J.; Oupicky, D. Tumor-penetrating nanoparticles for enhanced anticancer activity of combined photodynamic and hypoxia-activated therapy. ACS Nano 2017, 11, 2227–2238. [Google Scholar] [CrossRef] [PubMed]
- Mackowiak, S.A.; Schmidt, A.; Weiss, V.; Argyo, C.; von Schirnding, C.; Bein, T.; Brauchle, C. Targeted drug delivery in cancer cells with red-light photoactivated mesoporous silica nanoparticles. Nano Lett. 2013, 13, 2576–2583. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Lei, Q.; Jia, H.Z.; Wang, S.B.; Yin, W.N.; Chen, W.H.; Cheng, S.X.; Zhang, X.Z. A tumor targeted chimeric peptide for synergistic endosomal escape and therapy by dual-stage light manipulation. Adv. Funct. Mater. 2015, 25, 1248–1257. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Y.; Kilchrist, K.V.; Li, J.; Duvall, C.L.; Oupicky, D. Endosomolytic and tumor-penetrating mesoporous silica nanoparticles for siRNA/miRNA combination cancer therapy. ACS Appl. Mater. Interfaces 2020, 12, 4308–4322. [Google Scholar] [CrossRef]
- Fu, C.; Liu, T.; Li, L.; Liu, H.; Chen, D.; Tang, F. The absorption, distribution, excretion and toxicity of mesoporous silica nanoparticles in mice following different exposure routes. Biomaterials 2013, 34, 2565–2575. [Google Scholar] [CrossRef]
- Ding, Y.; Jiang, Z.; Saha, K.; Kim, C.S.; Kim, S.T.; Landis, R.F.; Rotello, V.M. Gold nanoparticles for nucleic acid delivery. Mol. Ther. 2014, 22, 1075–1083. [Google Scholar] [CrossRef] [Green Version]
- Rosi, N.L.; Giljohann, D.A.; Thaxton, C.S.; Lytton-Jean, A.K.; Han, M.S.; Mirkin, C.A. Oligonucleotide-modified gold nanoparticles for intracellular gene regulation. Science 2006, 312, 1027–1030. [Google Scholar] [CrossRef]
- Ekin, A.; Karatas, O.F.; Culha, M.; Ozen, M. Designing a gold nanoparticle-based nanocarrier for microRNA transfection into the prostate and breast cancer cells. J. Gene Med. 2014, 16, 331–335. [Google Scholar] [CrossRef]
- Ramchandani, D.; Lee, S.K.; Yomtoubian, S.; Han, M.S.; Tung, C.H.; Mittal, V. Nanoparticle delivery of miR-708 mimetic impairs breast cancer metastasis. Mol. Cancer Ther. 2019, 18, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.S.; Hung, Y.C.; Liau, I.; Huang, G.S. Assessment of the in vivo toxicity of gold nanoparticles. Nanoscale Res. Lett. 2009, 4, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Lasagna-Reeves, C.; Gonzalez-Romero, D.; Barria, M.A.; Olmedo, I.; Clos, A.; Sadagopa Ramanujam, V.M.; Urayama, A.; Vergara, L.; Kogan, M.J.; Soto, C. Bioaccumulation and toxicity of gold nanoparticles after repeated administration in mice. Biochem. Biophys. Res. Commun. 2010, 393, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Breunig, M.; Lungwitz, U.; Liebl, R.; Goepferich, A. Breaking up the correlation between efficacy and toxicity for nonviral gene delivery. Proc. Natl. Acad. Sci USA 2007, 104, 14454–14459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffert, D.; Wagner, E. Gene therapy progress and prospects: Synthetic polymer-based systems. Gene Ther. 2008, 15, 1131–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lungwitz, U.; Breunig, M.; Blunk, T.; Gopferich, A. Polyethylenimine-based non-viral gene delivery systems. Eur. J. Pharm. Biopharm. 2005, 60, 247–266. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, X. MicroRNA delivery with bioreducible polyethylenimine as a non-viral vector for breast cancer gene therapy. Macromol. Biosci. 2019, 19, e1800445. [Google Scholar] [CrossRef]
- Gao, S.; Tian, H.; Guo, Y.; Li, Y.; Guo, Z.; Zhu, X.; Chen, X. miRNA oligonucleotide and sponge for miRNA-21 inhibition mediated by PEI-PLL in breast cancer therapy. Acta Biomater. 2015, 25, 184–193. [Google Scholar] [CrossRef]
- Devulapally, R.; Paulmurugan, R. Polymer nanoparticles for drug and small silencing RNA delivery to treat cancers of different phenotypes. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2014, 6, 40–60. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhang, J.; Wang, Y.; Chen, M. Hyaluronic acid-coated PEI-PLGA nanoparticles mediated co-delivery of doxorubicin and miR-542-3p for triple negative breast cancer therapy. Nanomedicine 2016, 12, 411–420. [Google Scholar] [CrossRef]
- Devulapally, R.; Sekar, N.M.; Sekar, T.V.; Foygel, K.; Massoud, T.F.; Willmann, J.K.; Paulmurugan, R. Polymer nanoparticles mediated codelivery of antimiR-10b and antimiR-21 for achieving triple negative breast cancer therapy. ACS Nano 2015, 9, 2290–2302. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.; Sun, W.; Kissel, T. Chitosan-based formulations for delivery of DNA and siRNA. Adv. Drug Deliv. Rev. 2010, 62, 12–27. [Google Scholar] [CrossRef]
- Lee, M.; Nah, J.W.; Kwon, Y.; Koh, J.J.; Ko, K.S.; Kim, S.W. Water-soluble and low molecular weight chitosan-based plasmid DNA delivery. Pharm. Res. 2001, 18, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Corsi, K.; Chellat, F.; Fernandes, J.C. Mesenchymal stem cells, MG63 and HEK293 transfection using chitosan-DNA nanoparticles. Biomaterials 2003, 24, 1255–1264. [Google Scholar] [CrossRef]
- Huang, M.; Khor, E.; Lim, L.Y. Uptake and cytotoxicity of chitosan molecules and nanoparticles: Effects of molecular weight and degree of deacetylation. Pharm. Res. 2004, 21, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Santos-Carballal, B.; Aaldering, L.J.; Ritzefeld, M.; Pereira, S.; Sewald, N.; Moerschbacher, B.M.; Gotte, M.; Goycoolea, F.M. Physicochemical and biological characterization of chitosan-microRNA nanocomplexes for gene delivery to MCF-7 breast cancer cells. Sci. Rep. 2015, 5, 13567. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Cao, M.; Zhang, J.; Hu, K.; Yin, Z.; Zhou, Z.; Xiao, X.; Yang, Y.; Sheng, W.; Wu, Y.; et al. Hyaluronic acid-chitosan nanoparticles for co-delivery of MiR-34a and doxorubicin in therapy against triple negative breast cancer. Biomaterials 2014, 35, 4333–4344. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Oliva, N.; Atilano, M.; Song, H.S.; Artzi, N. Self-assembled RNA-triple-helix hydrogel scaffold for microRNA modulation in the tumour microenvironment. Nat. Mater. 2016, 15, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Akao, Y.; Iio, A.; Itoh, T.; Noguchi, S.; Itoh, Y.; Ohtsuki, Y.; Naoe, T. Microvesicle-mediated RNA molecule delivery system using monocytes/macrophages. Mol. Ther. 2011, 19, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Bose, R.J.C.; Uday Kumar, S.; Zeng, Y.; Afjei, R.; Robinson, E.; Lau, K.; Bermudez, A.; Habte, F.; Pitteri, S.J.; Sinclair, R.; et al. Tumor cell-derived extracellular vesicle-coated nanocarriers: An efficient theranostic platform for the cancer-specific delivery of anti-miR-21 and imaging agents. ACS Nano 2018, 12, 10817–10832. [Google Scholar] [CrossRef]
- Ohno, S.; Takanashi, M.; Sudo, K.; Ueda, S.; Ishikawa, A.; Matsuyama, N.; Fujita, K.; Mizutani, T.; Ohgi, T.; Ochiya, T.; et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol. Ther. 2013, 21, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.D.; Koo, B.H.; Kim, Y.H.; Jeon, O.H.; Kim, D.S. Exosome release of ADAM15 and the functional implications of human macrophage-derived ADAM15 exosomes. FASEB J. 2012, 26, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Didiot, M.C.; Hall, L.M.; Coles, A.H.; Haraszti, R.A.; Godinho, B.M.; Chase, K.; Sapp, E.; Ly, S.; Alterman, J.F.; Hassler, M.R.; et al. Exosome-mediated delivery of hydrophobically modified siRNA for huntingtin mRNA silencing. Mol. Ther. 2016, 24, 1836–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, C.; Tian, J.; Wang, Z.; Gao, Y.; Wu, X.; Ding, X.; Qiang, L.; Li, G.; Han, Z.; Yuan, Y.; et al. Functional exosome-mediated co-delivery of doxorubicin and hydrophobically modified microRNA 159 for triple-negative breast cancer therapy. J. Nanobiotechnol. 2019, 17, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usman, W.M.; Pham, T.C.; Kwok, Y.Y.; Vu, L.T.; Ma, V.; Peng, B.; Chan, Y.S.; Wei, L.; Chin, S.M.; Azad, A.; et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat. Commun. 2018, 9, 2359. [Google Scholar] [CrossRef] [PubMed]
- Ebert, M.S.; Sharp, P.A. MicroRNA sponges: Progress and possibilities. RNA 2010, 16, 2043–2050. [Google Scholar] [CrossRef] [Green Version]
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 2007, 4, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Xing, L.; Zheng, X.; Sun, Y.; Wang, X.; Chen, J. The circRNA circAGFG1 acts as a sponge of miR-195-5p to promote triple-negative breast cancer progression through regulating CCNE1 expression. Mol. Cancer 2019, 18, 4. [Google Scholar] [CrossRef]
- Wu, J.; Jiang, Z.; Chen, C.; Hu, Q.; Fu, Z.; Chen, J.; Wang, Z.; Wang, Q.; Li, A.; Marks, J.R.; et al. CircIRAK3 sponges miR-3607 to facilitate breast cancer metastasis. Cancer Lett. 2018, 430, 179–192. [Google Scholar] [CrossRef]
- Mandal, C.C.; Ghosh-Choudhury, T.; Dey, N.; Choudhury, G.G.; Ghosh-Choudhury, N. miR-21 is targeted by omega-3 polyunsaturated fatty acid to regulate breast tumor CSF-1 expression. Carcinogenesis 2012, 33, 1897–1908. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Liu, J.; Song, Q.; Yang, X.; Wang, D.; Liu, W.; Shi, J.; Zhang, Z. DNA Nanosponge for adsorption and clearance of intracellular miR-21 and enhanced antitumor chemotherapy. ACS Appl. Mater. Interfaces 2019, 11, 46604–46613. [Google Scholar] [CrossRef]
- Lei, K.; Du, W.; Lin, S.; Yang, L.; Xu, Y.; Gao, Y.; Xu, B.; Tan, S.; Xu, Y.; Qian, X.; et al. 3B, a novel photosensitizer, inhibits glycolysis and inflammation via miR-155-5p and breaks the JAK/STAT3/SOCS1 feedback loop in human breast cancer cells. Biomed. Pharmacother. 2016, 82, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Costales, M.G.; Aikawa, H.; Li, Y.; Childs-Disney, J.L.; Abegg, D.; Hoch, D.G.; Pradeep Velagapudi, S.; Nakai, Y.; Khan, T.; Wang, K.W.; et al. Small-molecule targeted recruitment of a nuclease to cleave an oncogenic RNA in a mouse model of metastatic cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 2406–2411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Umeh-Garcia, M.; Simion, C.; Ho, P.Y.; Batra, N.; Berg, A.L.; Carraway, K.L.; Yu, A.; Sweeney, C. A novel bioengineered miR-127 prodrug suppresses the growth and metastatic potential of triple-negative breast cancer cells. Cancer Res. 2020, 80, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellana, E.A.; Tenneti, S.; Rangasamy, L.; Lyle, L.T.; Low, P.S.; Kasinski, A.L. FolamiRs: Ligand-targeted, vehicle-free delivery of microRNAs for the treatment of cancer. Sci. Transl. Med. 2017, 9, eaam9327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, J.; Tacha, D.E.; Li, P.; Bremer, R.E.; Chen, H.; Wei, B.; Xiao, X.; Da, J.; Skinner, K.; et al. Folate receptor alpha associated with triple-negative breast cancer and poor prognosis. Arch. Pathol. Lab. Med. 2014, 138, 890–895. [Google Scholar] [CrossRef]
- Hosseinahli, N.; Aghapour, M.; Duijf, P.H.G.; Baradaran, B. Treating cancer with microRNA replacement therapy: A literature review. J. Cell Physiol. 2018, 233, 5574–5588. [Google Scholar] [CrossRef]
- Lima, J.F.; Cerqueira, L.; Figueiredo, C.; Oliveira, C.; Azevedo, N.F. Anti-miRNA oligonucleotides: A comprehensive guide for design. RNA Biol. 2018, 15, 338–352. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Sarkar, B.K.; Lee, S.S. The novel strategies for next-generation cancer treatment: miRNA combined with chemotherapeutic agents for the treatment of cancer. Oncotarget 2018, 9, 10164–10174. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
miRNAs involved in potentiating various stages and events in breast cancer metastasis: (i) angiogenesis, (ii) EMT, migration and invasion, (iii) the Warburg effect, and (iv) the tumour microenvironment.
Figure 1.
miRNAs involved in potentiating various stages and events in breast cancer metastasis: (i) angiogenesis, (ii) EMT, migration and invasion, (iii) the Warburg effect, and (iv) the tumour microenvironment.

Figure 2.
Metastatic cascade in cancer.

Figure 3.
Crosstalk between breast cancer cells and stromal cells in the tumour microenvironment.

Figure 4.
Summary of approaches taken in miRNA-based therapies for breast cancer.


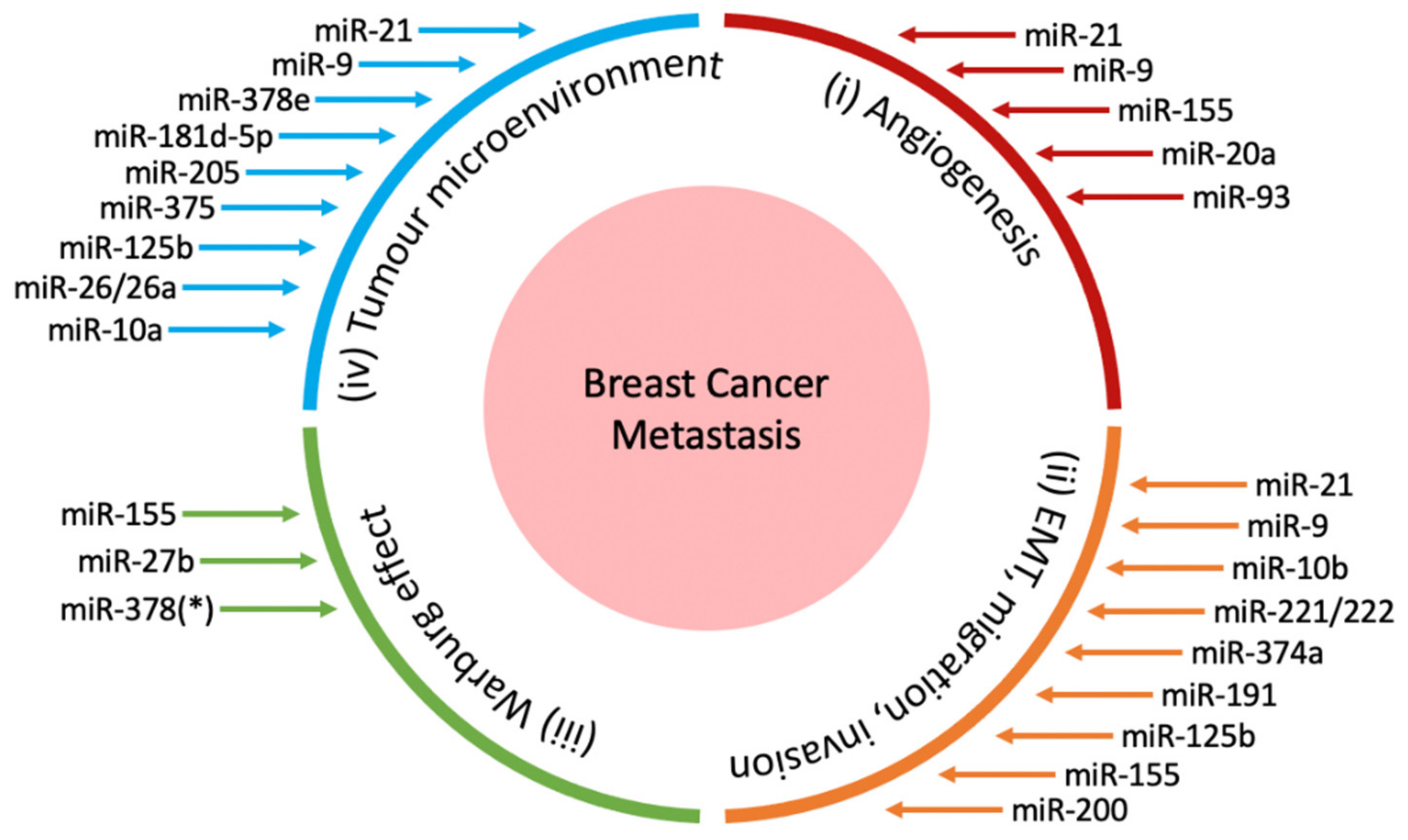{kind=link}
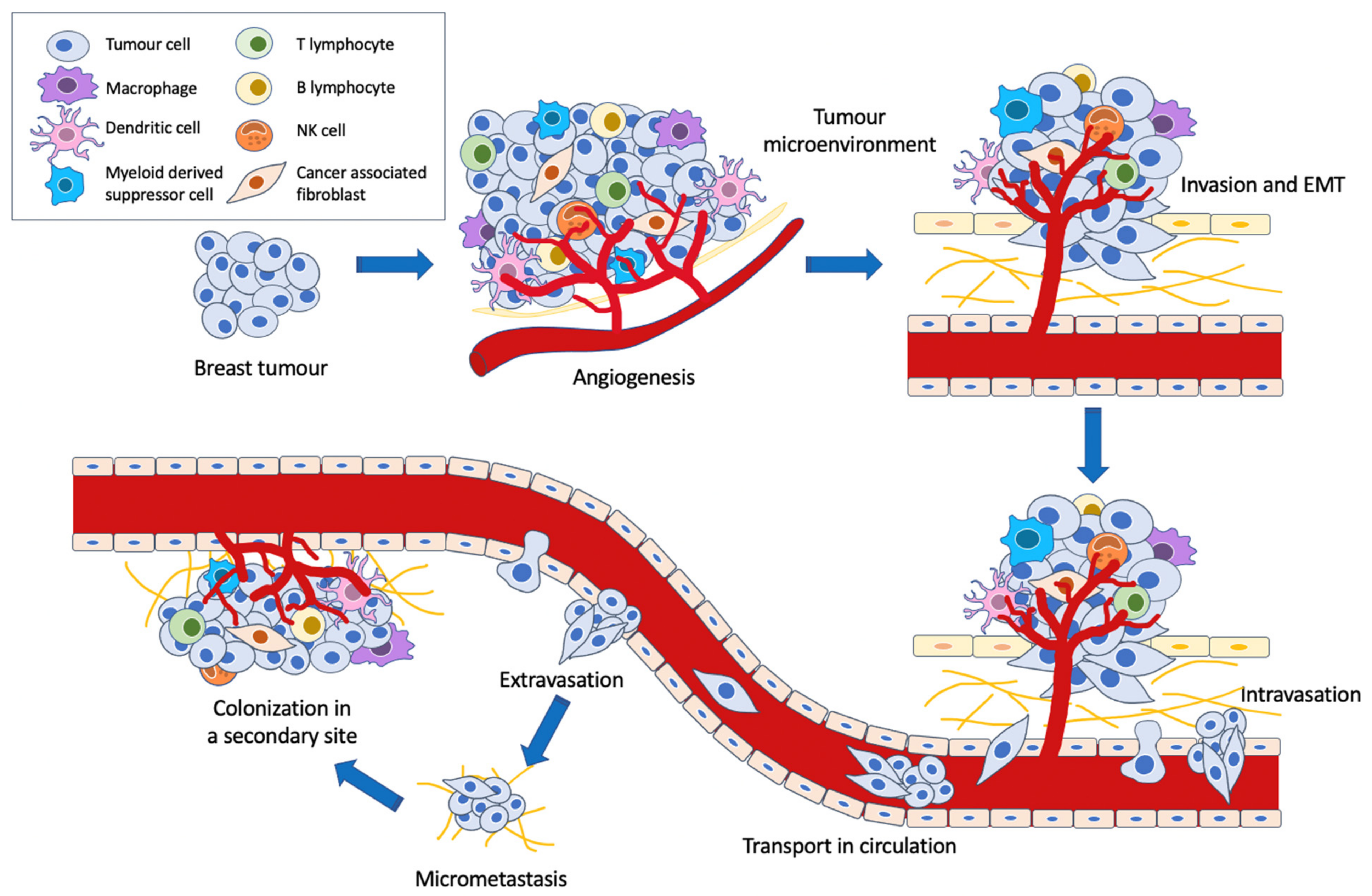{kind=link}
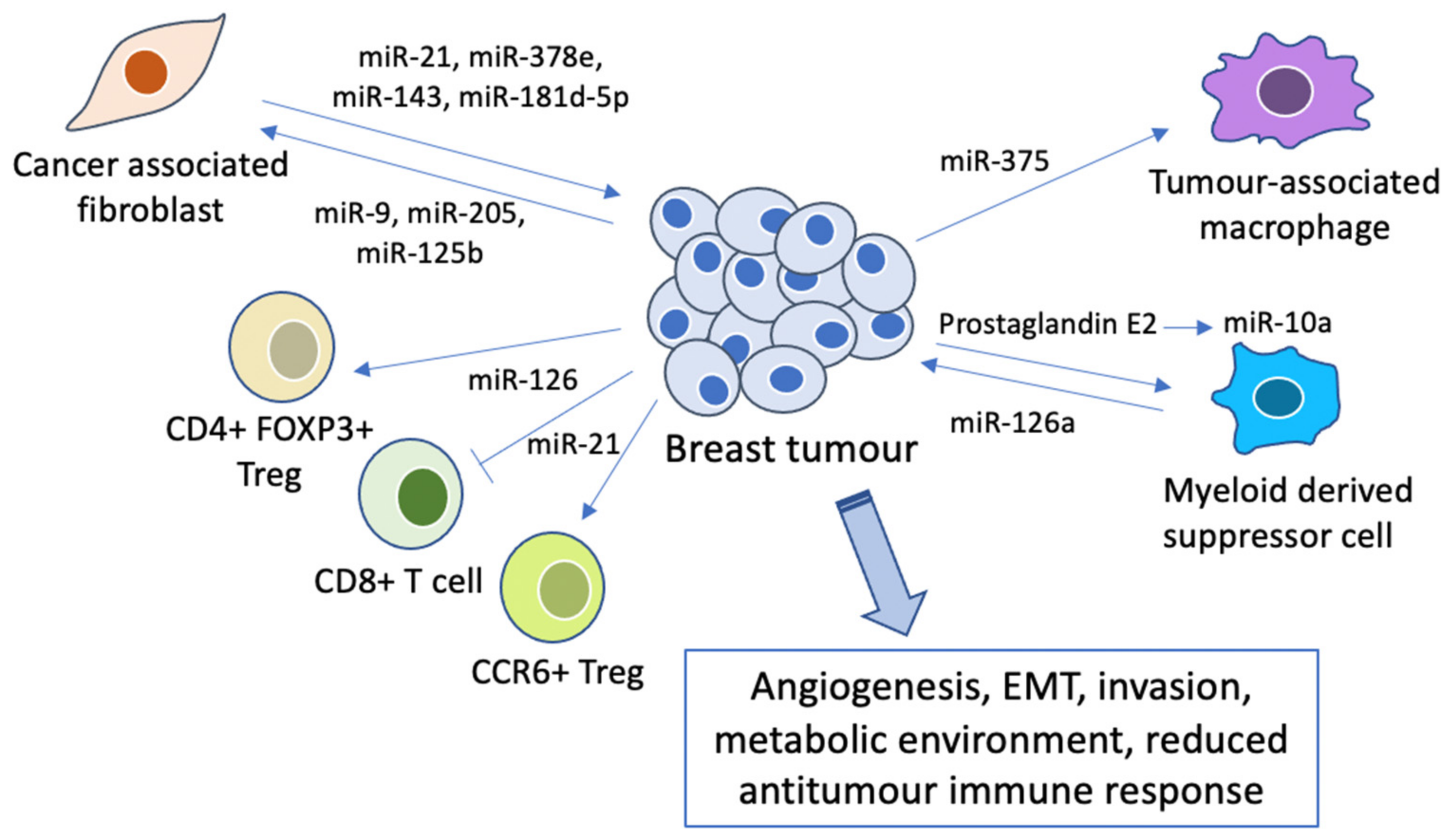{kind=link}
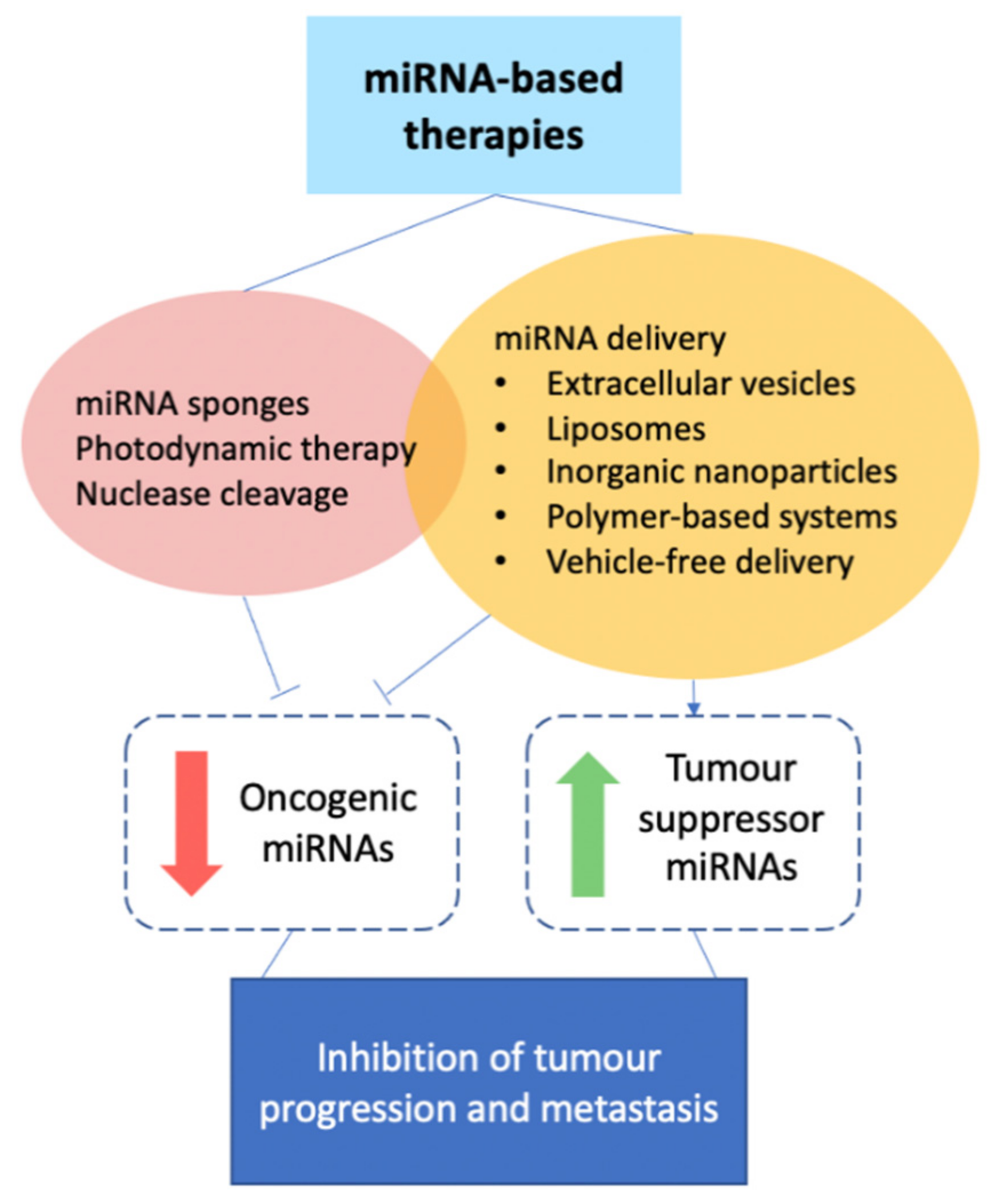{kind=link}
Table 1.
miRNAs involved in promoting angiogenesis.
| miRNA | Expression in Breast Cancer | Target(s) | Experimental Design | Reference(s) | |
|---|---|---|---|---|---|
| Oncogenic miRNAs | miR-9 | Upregulated | CDH1/ß-catenin/VEGF | In vitro & in vivo | [40] |
| miR-155 | Upregulated | VHL | In vitro & in vivo | [41] | |
| miR-20a | Upregulated | VEGFA | In vitro | [42] | |
| miR-93 | Upregulated | LATS2 | In vitro & in vivo | [43] | |
| miR-21 | Upregulated | VEGF/VEGFR2/HIF1α | In vitro & in vivo | [44] |
Table 2.
miRNAs involved in promoting EMT, invasion and migration.
| miRNA | Expression in Breast Cancer | Target(s) | Experimental Design | Reference(s) | |
|---|---|---|---|---|---|
| Oncogenic miRNAs | miR-21 | Upregulated | LZTFL1, PTEN | In vitro & in vivo | [60] |
| miR-9 | Upregulated | CDH1 | In vitro & in vivo | [40] | |
| miR-10b | Upregulated | Homeobox D10 | In vitro & in vivo | [61] | |
| miR-221/222 | Upregulated | TRPS1 | In vitro | [62] | |
| miR-374a | Upregulated | WIF1, PTEN, WNT5A | In vitro & in vivo | [63] | |
| miR-191 | Upregulated | TGFß2 | In vitro | [64] | |
| Both tumour suppressors & oncogenic miRNAs | miR-125b | DownregulatedUpregulated | SNAIL-1, SEMA4C STARD13 | In vitro In vitro & in vivo | [65,66,67] |
| miR-155 | DownregulatedUpregulated | TCF4, ZEB2 C/EBPß, ZNF652 | In vitro & in vivo In vitro & in vivo | [68,69,70,71] | |
| miR-200 | DownregulatedUpregulated | ZEB1, ZEB2, FHOD1, PPM1F ZEB2, SEC23a, CDH1 | In vitro In vitro & in vivo | [72,73,74,75,76,77,78] |
Table 3.
miRNAs regulating the Warburg effect.
| miRNA | Expression in Breast Cancer | Target(s) | Experimental Design | Reference(s) | |
|---|---|---|---|---|---|
| Oncogenic miRNAs | miR-155 | Upregulated | PIK3R1, FOXO3a, STAT3, C/EBPβ | In vitro & in vivo | [95,96,97] |
| miR-27b | Upregulated | PDHX | In vitro | [98] | |
| miR-378(*) # | Upregulated | ERRγ and GABPA | In vitro | [99] |
# miR-378(*) may also be known as miR-378a-5p [100].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Teo, A.Y.T.; Xiang, X.; Le, M.T.; Wong, A.L.-A.; Zeng, Q.; Wang, L.; Goh, B.-C. Tiny miRNAs Play a Big Role in the Treatment of Breast Cancer Metastasis. Cancers 2021, 13, 337. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13020337
AMA Style
Teo AYT, Xiang X, Le MT, Wong AL-A, Zeng Q, Wang L, Goh B-C. Tiny miRNAs Play a Big Role in the Treatment of Breast Cancer Metastasis. Cancers. 2021; 13(2):337. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13020337
Chicago/Turabian StyleTeo, Andrea York Tiang, Xiaoqiang Xiang, Minh TN Le, Andrea Li-Ann Wong, Qi Zeng, Lingzhi Wang, and Boon-Cher Goh. 2021. "Tiny miRNAs Play a Big Role in the Treatment of Breast Cancer Metastasis" Cancers 13, no. 2: 337. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13020337
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.