
Copy Neutral LOH Affecting the Entire Chromosome 6 Is a Frequent Mechanism of HLA Class I Alterations in Cancer

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Tumor Cell Lines and Tumors
2.2. Bladder Tumor Tissue Immunohistochemistry and Microdissection
2.3. Analysis of HLA Genomic Copy Number Alterations by Single Nucleotide Polymorphism (SNP) Arrays
3. Results
Analysis of LOH-6 and LOH-15 in Human Cancer Cell Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garrido, F.; Cabrera, T.; Concha, A.; Glew, S.; Ruiz-Cabello, F.; Stern, P.L. Natural history of HLA expression during tumour development. Immunol. Today 1993, 14, 491–499. [Google Scholar] [CrossRef]
- Browning, M.; Petronzelli, F.; Bicknell, D.; Krausa, P.; Rowan, A.; Tonks, S.; Murray, N.; Bodmer, J.; Bodmer, W. Mechanisms of loss of HLA class I expression on colorectal tumor cells. Tissue Antigens 1996, 47, 364–371. [Google Scholar] [CrossRef]
- Koopman, L.A.; Corver, W.E.; van der Slik, A.R.; Giphart, M.J.; Fleuren, G.J. Multiple genetic alterations cause frequent and heterogeneous human histocompatibility leukocyte antigen class I loss in cervical cancer. J. Exp. Med. 2000, 191, 961–976. [Google Scholar] [CrossRef]
- Romero, J.M.; Jimenez, P.; Cabrera, T.; Cozar, J.M.; Pedrinaci, S.; Tallada, M.; Garrido, F.; Ruiz-Cabello, F. Coordinated downregulation of the antigen presentation machinery and HLA class I/beta2-microglobulin complex is responsible for HLA-ABC loss in bladder cancer. Int. J. Cancer 2005, 113, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.; Canton, J.; Collado, A.; Cabrera, T.; Serrano, A.; Real, L.M.; Garcia, A.; Ruiz-Cabello, F.; Garrido, F. Chromosome loss is the most frequent mechanism contributing to HLA haplotype loss in human tumors. Int. J. Cancer 1999, 83, 91–97. [Google Scholar] [CrossRef]
- Serrano, A.; Tanzarella, S.; Lionello, I.; Mendez, R.; Traversari, C.; Ruiz-Cabello, F.; Garrido, F. Rexpression of HLA class I antigens and restoration of antigen-specific CTL response in melanoma cells following 5-aza-2’-deoxycytidine treatment. Int. J. Cancer 2001, 94, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.J.; Ruiz-Cabello, F.; Skoudy, A.; Berrozpe, G.; Jimenez, P.; Serrano, A.; Real, F.X.; Garrido, F. Loss of an HLA haplotype in pancreas cancer tissue and its corresponding tumor derived cell line. Tissue Antigens 1996, 47, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Maleno, I.; Cabrera, C.M.; Cabrera, T.; Paco, L.; Lopez-Nevot, M.A.; Collado, A.; Ferron, A.; Garrido, F. Distribution of HLA class I altered phenotypes in colorectal carcinomas: High frequency of HLA haplotype loss associated with loss of heterozygosity in chromosome region 6p21. Immunogenetics 2004, 56, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Maleno, I.; Romero, J.M.; Cabrera, T.; Paco, L.; Aptsiauri, N.; Cozar, J.M.; Tallada, M.; Lopez-Nevot, M.A.; Garrido, F. LOH at 6p21.3 region and HLA class I altered phenotypes in bladder carcinomas. Immunogenetics 2006, 58, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Perea, F.; Sanchez-Palencia, A.; Gomez-Morales, M.; Bernal, M.; Concha, A.; Garcia, M.M.; Gonzalez-Ramirez, A.R.; Kerick, M.; Martin, J.; Garrido, F.; et al. HLA class I loss and PD-L1 expression in lung cancer: Impact on T-cell infiltration and immune escape. Oncotarget 2017, 9, 4120–4133. [Google Scholar] [CrossRef] [Green Version]
- Paschen, A.; Mendez, R.M.; Jimenez, P.; Sucker, A.; Ruiz-Cabello, F.; Song, M.; Garrido, F.; Schadendorf, D. Complete loss of HLA class I antigen expression on melanoma cells: A result of successive mutational events. Int. J. Cancer 2003, 103, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Restifo, N.P.; Marincola, F.M.; Kawakami, Y.; Taubenberger, J.; Yannelli, J.R.; Rosenberg, S.A. Loss of functional beta 2-microglobulin in metastatic melanomas from five patients receiving immunotherapy. J. Natl. Cancer Inst. 1996, 88, 100–108. [Google Scholar] [CrossRef]
- Del Campo, A.B.; Kyte, J.A.; Carretero, J.; Zinchencko, S.; Mendez, R.; Gonzalez-Aseguinolaza, G.; Ruiz-Cabello, F.; Aamdal, S.; Gaudernack, G.; Garrido, F.; et al. Immune escape of cancer cells with beta2-microglobulin loss over the course of metastatic melanoma. Int. J. Cancer 2014, 134, 102–113. [Google Scholar] [CrossRef]
- Garrido, F.; Cabrera, T.; Aptsiauri, N. “Hard” and “soft” lesions underlying the HLA class I alterations in cancer cells: Implications for immunotherapy. Int. J. Cancer 2010, 127, 249–256. [Google Scholar] [CrossRef]
- Bernal, M.; Ruiz-Cabello, F.; Concha, A.; Paschen, A.; Garrido, F. Implication of the beta2-microglobulin gene in the generation of tumor escape phenotypes. Cancer Immunol. Immunother. 2012, 61, 1359–1371. [Google Scholar] [CrossRef]
- Maleno, I.; Aptsiauri, N.; Cabrera, T.; Gallego, A.; Paschen, A.; Lopez-Nevot, M.A.; Garrido, F. Frequent loss of heterozygosity in the beta2-microglobulin region of chromosome 15 in primary human tumors. Immunogenetics 2011, 63, 65–71. [Google Scholar] [CrossRef]
- Chowell, D.; Morris, L.G.T.; Grigg, C.M.; Weber, J.K.; Samstein, R.M.; Makarov, V.; Kuo, F.; Kendall, S.M.; Requena, D.; Riaz, N.; et al. Patient HLA class I genotype influences cancer response to checkpoint blockade immunotherapy. Science 2018, 359, 582–587. [Google Scholar] [CrossRef] [Green Version]
- Carretero, R.; Cabrera, T.; Gil, H.; Saenz-Lopez, P.; Maleno, I.; Aptsiauri, N.; Cozar, J.M.; Garrido, F. Bacillus Calmette-Guerin immunotherapy of bladder cancer induces selection of human leukocyte antigen class I-deficient tumor cells. Int. J. Cancer 2011, 129, 839–846. [Google Scholar] [CrossRef]
- Zaretsky, J.M.; Garcia-Diaz, A.; Shin, D.S.; Escuin-Ordinas, H.; Hugo, W.; Hu-Lieskovan, S.; Torrejon, D.Y.; Abril-Rodriguez, G.; Sandoval, S.; Barthly, L.; et al. Mutations Associated with Acquired Resistance to PD-1 Blockade in Melanoma. N. Engl. J. Med. 2016, 375, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Gettinger, S.; Choi, J.; Hastings, K.; Truini, A.; Datar, I.; Sowell, R.; Wurtz, A.; Dong, W.; Cai, G.; Melnick, M.A.; et al. Impaired HLA Class I Antigen Processing and Presentation as a Mechanism of Acquired Resistance to Immune Checkpoint Inhibitors in Lung Cancer. Cancer Discov. 2017, 7, 1420–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sade-Feldman, M.; Jiao, Y.J.; Chen, J.H.; Rooney, M.S.; Barzily-Rokni, M.; Eliane, J.P.; Bjorgaard, S.L.; Hammond, M.R.; Vitzthum, H.; Blackmon, S.M.; et al. Resistance to checkpoint blockade therapy through inactivation of antigen presentation. Nat. Commun. 2017, 8, 1136. [Google Scholar] [CrossRef]
- Montesion, M.; Murugesan, K.; Jin, D.X.; Sharaf, R.; Sanchez, N.; Guria, A.; Minker, M.; Li, G.; Fisher, V.; Sokol, E.S.; et al. Somatic HLA Class I Loss Is a Widespread Mechanism of Immune Evasion Which Refines the Use of Tumor Mutational Burden as a Biomarker of Checkpoint Inhibitor Response. Cancer Discov. 2021, 11, 282–292. [Google Scholar] [CrossRef]
- Luo, G.; Santoro, I.M.; McDaniel, L.D.; Nishijima, I.; Mills, M.; Youssoufian, H.; Vogel, H.; Schultz, R.A.; Bradley, A. Cancer predisposition caused by elevated mitotic recombination in Bloom mice. Nat. Genet. 2000, 26, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Kumar, Y.; Yang, J.; Hu, T.; Chen, L.; Xu, Z.; Xu, L.; Hu, X.X.; Tang, G.; Wang, J.M.; Li, Y.; et al. Massive interstitial copy-neutral loss-of-heterozygosity as evidence for cancer being a disease of the DNA-damage response. BMC Med. Genom. 2015, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Thiagalingam, S.; Laken, S.; Willson, J.K.; Markowitz, S.D.; Kinzler, K.W.; Vogelstein, B.; Lengauer, C. Mechanisms underlying losses of heterozygosity in human colorectal cancers. Proc. Natl. Acad. Sci. USA 2001, 98, 2698–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, T.; Mendez, R.; Roberts, C.H.; Ruiz-Cabello, F.; Dodi, I.A.; Lopez Nevot, M.A.; Paco, L.; Maleno, I.; Marsh, S.G.; Pawelec, G.; et al. High frequency of homozygosity of the HLA region in melanoma cell lines reveals a pattern compatible with extensive loss of heterozygosity. Cancer Immunol. Immunother. 2005, 54, 141–148. [Google Scholar] [CrossRef]
- Mendez, R.; Rodriguez, T.; Del Campo, A.; Monge, E.; Maleno, I.; Aptsiauri, N.; Jimenez, P.; Pedrinaci, S.; Pawelec, G.; Ruiz-Cabello, F.; et al. Characterization of HLA class I altered phenotypes in a panel of human melanoma cell lines. Cancer Immunol. Immunother. 2008, 57, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Montes, P.; Kerick, M.; Bernal, M.; Hernandez, F.; Jimenez, P.; Garrido, P.; Marquez, A.; Jurado, M.; Martin, J.; Garrido, F.; et al. Genomic loss of HLA alleles may affect the clinical outcome in low-risk myelodysplastic syndrome patients. Oncotarget 2018, 9, 36929–36944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, F.; Santos-Aguado, J.; Borche, L.; Places, L.; Domenech, N.; Gaya, A.; Vilella, R.; Vives, J. Identification of the amino acid residues defining an intralocus determinant in the alpha 1 domain of HLA-A molecules. Immunogenetics 1989, 30, 50–53. [Google Scholar] [CrossRef]
- Lozano, F.; Borche, L.; Places, L.; Alberola-Ila, J.; Gaya, A.; Vilella, R.; Vives, J. Biochemical and serological characterization of a public antigenic determinant present on HLA-B molecules. Tissue Antigens 1990, 35, 193–195. [Google Scholar] [CrossRef]
- Doherty, P.C.; Zinkernagel, R.M. Enhanced immunological surveillance in mice heterozygous at the H-2 gene complex. Nature 1975, 256, 50–52. [Google Scholar] [CrossRef]
- Garrido, F.; Ruiz-Cabello, F.; Cabrera, T.; Perez-Villar, J.J.; Lopez-Botet, M.; Duggan-Keen, M.; Stern, P.L. Implications for immunosurveillance of altered HLA class I phenotypes in human tumours. Immunol. Today 1997, 18, 89–95. [Google Scholar] [CrossRef]
- McGranahan, N.; Rosenthal, R.; Hiley, C.T.; Rowan, A.J.; Watkins, T.B.K.; Wilson, G.A.; Birkbak, N.J.; Veeriah, S.; Van Loo, P.; Herrero, J.; et al. Allele-Specific HLA Loss and Immune Escape in Lung Cancer Evolution. Cell 2017, 171, 1259–1271.e11. [Google Scholar] [CrossRef] [Green Version]
- Yeung, J.T.; Hamilton, R.L.; Ohnishi, K.; Ikeura, M.; Potter, D.M.; Nikiforova, M.N.; Ferrone, S.; Jakacki, R.I.; Pollack, I.F.; Okada, H. LOH in the HLA class I region at 6p21 is associated with shorter survival in newly diagnosed adult glioblastoma. Clin. Cancer Res. 2013, 19, 1816–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harima, Y.; Harima, K.; Sawada, S.; Tanaka, Y.; Arita, S.; Ohnishi, T. Loss of heterozygosity on chromosome 6p21.2 as a potential marker for recurrence after radiotherapy of human cervical cancer. Clin. Cancer Res. 2000, 6, 1079–1085. [Google Scholar] [PubMed]
- Facoetti, A.; Nano, R.; Zelini, P.; Morbini, P.; Benericetti, E.; Ceroni, M.; Campoli, M.; Ferrone, S. Human leukocyte antigen and antigen processing machinery component defects in astrocytic tumors. Clin. Cancer Res. 2005, 11, 8304–8311. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Carrot-Zhang, J.; Zhao, Y.; Hu, H.; Freeman, S.S.; Yu, S.; Ha, G.; Taylor, A.M.; Berger, A.C.; Westlake, L.; et al. Genomic and immune profiling of pre-invasive lung adenocarcinoma. Nat. Commun. 2019, 10, 5472. [Google Scholar] [CrossRef] [PubMed]
- Iorgulescu, J.B.; Braun, D.; Oliveira, G.; Keskin, D.B.; Wu, C.J. Acquired mechanisms of immune escape in cancer following immunotherapy. Genome Med. 2018, 10, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carretero, R.; Romero, J.M.; Ruiz-Cabello, F.; Maleno, I.; Rodriguez, F.; Camacho, F.M.; Real, L.M.; Garrido, F.; Cabrera, T. Analysis of HLA class I expression in progressing and regressing metastatic melanoma lesions after immunotherapy. Immunogenetics 2008, 60, 439–447. [Google Scholar] [CrossRef]
- Mendez, R.; Ruiz-Cabello, F.; Rodriguez, T.; Del Campo, A.; Paschen, A.; Schadendorf, D.; Garrido, F. Identification of different tumor escape mechanisms in several metastases from a melanoma patient undergoing immunotherapy. Cancer Immunol. Immunother. 2007, 56, 88–94. [Google Scholar] [CrossRef]
- Lopez, S.; Lim, E.L.; Horswell, S.; Haase, K.; Huebner, A.; Dietzen, M.; Mourikis, T.P.; Watkins, T.B.K.; Rowan, A.; Dewhurst, S.M.; et al. Interplay between whole-genome doubling and the accumulation of deleterious alterations in cancer evolution. Nat. Genet. 2020, 52, 283–293. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
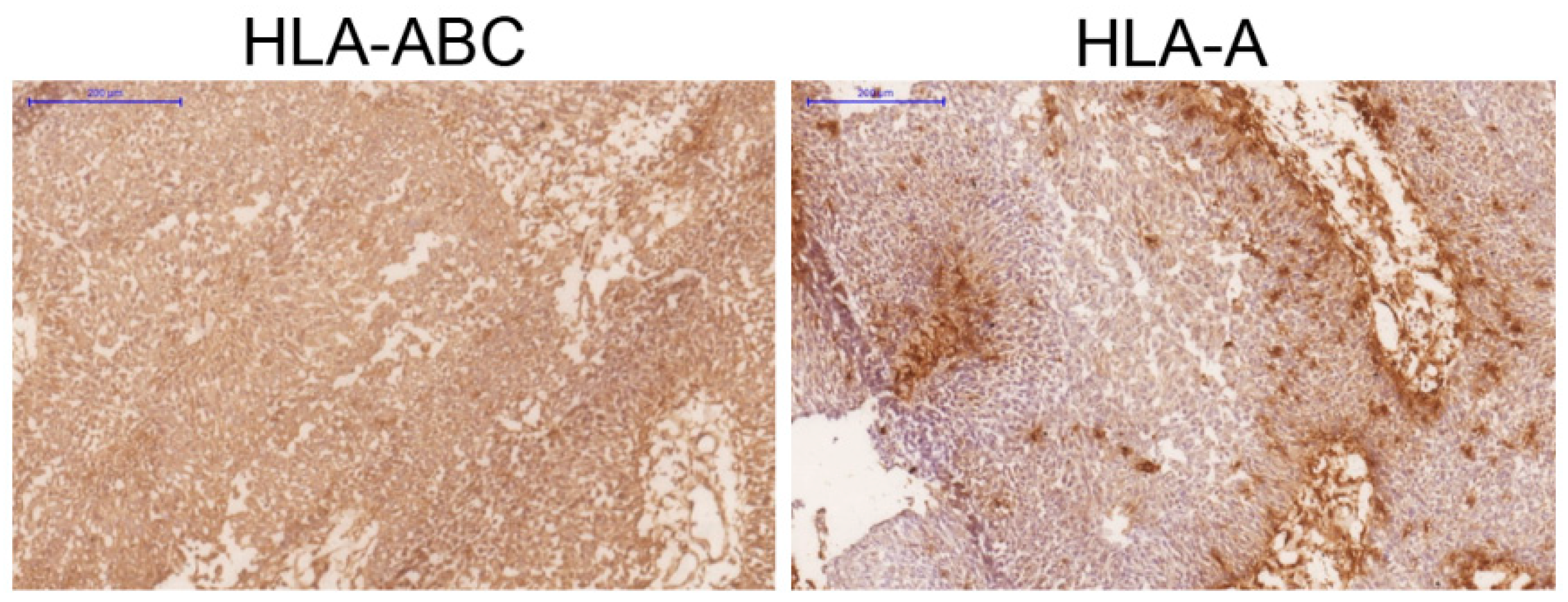{kind=link}
{kind=link}
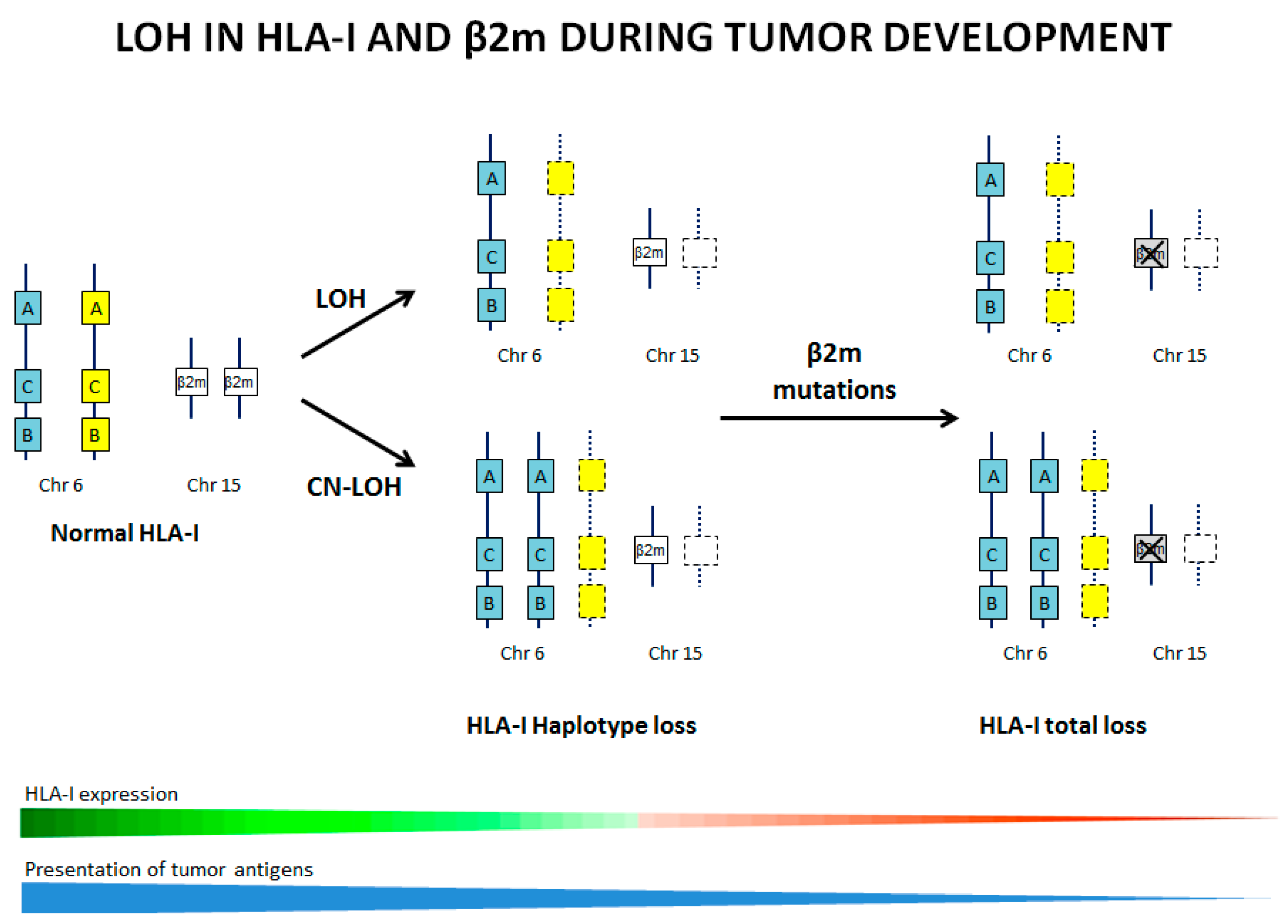{kind=link}
| Histological Type | Sample | SNP-A Chr6—Range | Size (Mb) | LOH of the HLA Region | SNP-A Chr15—Range | Size (Mb) | LOH of the B2M Gene |
|---|---|---|---|---|---|---|---|
| Melanoma (CCL) | |||||||
| E037 | CN-LOH 6pterqter | YES | CN-LOH 15q11.2qter * | YES | |||
| E040 | CN-LOH6pterp12.1,6pterq24.3 * | 55, 148 * | YES | normal | NO | ||
| E055 | CN-LOH 6pterqter *,6q12qter | 104 | YES | normal | NO | ||
| E058 | CN-LOH 6pterqter | YES | CN-LOH 15q22.2qter * | 41.8 * | NO | ||
| E062 | CN-LOH 6pterqter | YES | CN-LOH 15q11.2q23 * | 50 * | YES | ||
| E064 | CN-LOH 6pterqter * | YES | normal | NO | |||
| E070 | CN-LOH 6pterp21.3 | 33.4 | YES | normal | NO | ||
| E071 | CN-LOH 6pterqter | YES | normal | NO | |||
| E081 | CN-LOH 6pterqter | YES | normal | NO | |||
| E019 | CN-LOH 6pterqter *,6q15.1qter | 177 | YES | CN-LOH 15q14q22.2 | 25 | YES | |
| E124 | CN-LOH 6pterq12 | 65 | YES | CN-LOH 15q11.2qter * | YES | ||
| M010 | CN-LOH 6pterqter | YES | CN-LOH 15q11.2qter | YES | |||
| Ando | CN-LOH 6pterqter | YES | CN-LOH 15q11.2qter* | YES | |||
| Breast (CCL) | |||||||
| MDA-MB4-435 | CN-LOH 6pterqter | YES | CN-LOH 15q21.1q21.3 *,15q22.2qter * | 10 *,41 * | NO | ||
| IMIM-MA2 | CN-LOH 6pterqter | YES | CN-LOH 15q11.2q21.2 | 30 | YES | ||
| MCF-7 | CN-LOH 6p21.31q25.2 * | 122 * | NO | CN LOH 15q11.2q23 * | 50 * | YES | |
| MDA-MB-231 | Normal | NO | No data | No data | |||
| Leukemia (CCL) | |||||||
| X31364 | CN-LOH 6pterp21.31 | 34 | YES | normal | NO | ||
| X28751 | CN-LOH 6pterp12.3 | 47 | YES | normal | NO | ||
| X29708 | CN-LOH 6pterp21.31 | 36 | YES | normal | NO | ||
| X30279 | CN-LOH 6pterp21.2 | 38 | YES | normal | NO | ||
| Lung (CCL) | |||||||
| SKMES | CN-LOH 6pterqter *,6pterp12.2 | 52 | YES | CN-LOH 15q11.2qter * | YES | ||
| SKLU-1 | CN-LOH 6pterqter | YES | CN-LOH 15q11.2qter | YES | |||
| CALU6 | CN-LOH 6pterqter | YES | CN-LOH 15q11.2qter * | YES | |||
| A427 | CN-LOH 6pterqter | YES | normal | NO | |||
| Bladder (CCL) | |||||||
| RT4 (HTB-2) | CN-LOH 6q21q27 * | 49 | NO | CN-LOH 15q11.2qter * | YES | ||
| 5637 (HTB-9) | CN-LOH 6q16.3qter * | 66 * | NO | CN-LOH 15q11.2qter * | YES | ||
| TCCSUP (HTB-5) | CN-LOH 6pterp22.3,6q12q25.2 | 22, 87 | NO | CN-LOH 15q11.2qter * | YES | ||
| WILL | CN-LOH 6pterqter | YES | CN-LOH 15q11.2q14 *,15q21.3q26.2 * | 16 *, 42 * | NO | ||
| J82 (HTB-1) | CN-LOH 6pterqter | YES | normal | NO | |||
| T24 (HTB-4) | CN-LOH 6pterqter | YES | CN-LOH 15q11.2qter * | YES | |||
| Bladder (TT) | |||||||
| V039 | CN-LOH 6pterqter * | YES | normal | NO | |||
| V164 | CN-LOH 6pterp21.1 | 49.5 | YES | normal | NO | ||
| V24 | CN-LOH 6pterp21.2 | 39 | YES | normal | NO | ||
| V315 | CN-LOH 6q13qter | 95.1 | NO | CN-LOH 15q11.2qter | YES | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, M.A.; Perea, F.; Vilchez, J.R.; Rodríguez, T.; Anderson, P.; Garrido, F.; Ruiz-Cabello, F.; Aptsiauri, N. Copy Neutral LOH Affecting the Entire Chromosome 6 Is a Frequent Mechanism of HLA Class I Alterations in Cancer. Cancers 2021, 13, 5046. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205046
Garrido MA, Perea F, Vilchez JR, Rodríguez T, Anderson P, Garrido F, Ruiz-Cabello F, Aptsiauri N. Copy Neutral LOH Affecting the Entire Chromosome 6 Is a Frequent Mechanism of HLA Class I Alterations in Cancer. Cancers. 2021; 13(20):5046. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205046
Chicago/Turabian StyleGarrido, Maria Antonia, Francisco Perea, Jose Ramon Vilchez, Teresa Rodríguez, Per Anderson, Federico Garrido, Francisco Ruiz-Cabello, and Natalia Aptsiauri. 2021. "Copy Neutral LOH Affecting the Entire Chromosome 6 Is a Frequent Mechanism of HLA Class I Alterations in Cancer" Cancers 13, no. 20: 5046. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205046