
Lipid Droplet Accumulation Independently Predicts Poor Clinical Prognosis in High-Grade Serous Ovarian Carcinoma

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients with HGSOC and Samples
2.2. Materials
2.3. Immunohistochemical Analysis of ADP
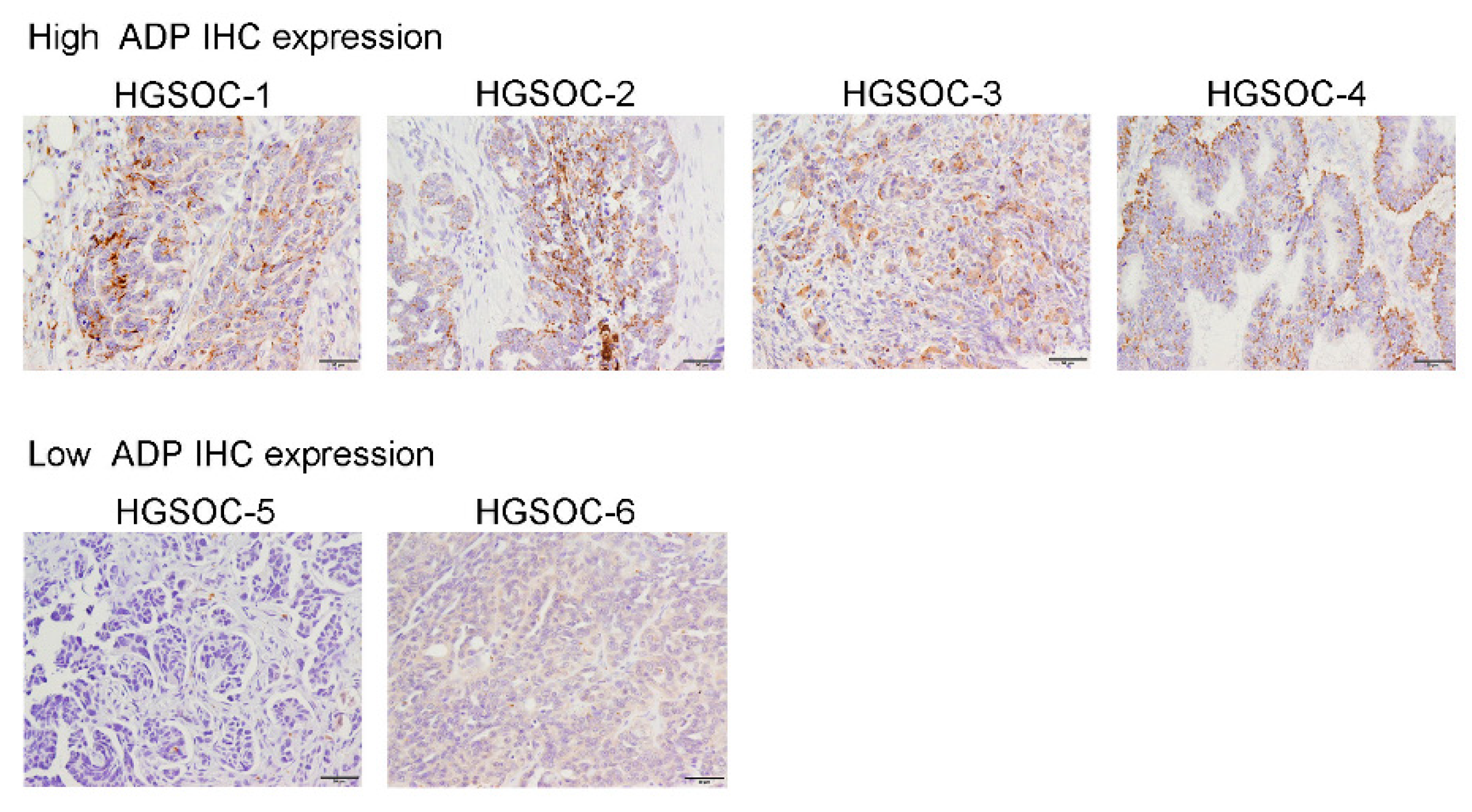
2.4. Scoring of ADP Immunostaining
2.5. Analysis of LD and ADP Expression
2.6. Analysis of Cell Survival-Related Pathways and Cell Proliferation
2.7. Statistical Analyses
3. Results
3.1. Cohort Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All (n = 96) | ADP-Low (n = 61) | ADP-High (n = 35) | p Value | |
|---|---|---|---|---|
| Age, years (mean ± SD) | 61.8 ± 11.2 | 61.8 ± 11.3 | 61.7 ± 11.3 | 0.985 |
| BMI, kg/m2 (mean ± SD) | 22.6 ± 4.4 | 21.9 ± 3.2 | 23.7 ± 5.8 | <0.001 * |
| FIGO stage, n (%) | <0.001 * | |||
| I | 4 (4.2) | 4 (6.6) | 0 (0.0) | |
| II | 7 (7.3) | 6 (9.8) | 1 (2.9) | |
| III | 69 (71.9) | 48 (78.7) | 21 (60.0) | |
| IV | 16 (16.7) | 3 (4.9) | 13 (37.1) | |
| Recurrence or progression, n (%) | 68 (70.8) | 36 (59.0) | 32 (91.4) | <0.001 * |
| Platinum sensitive, n (%) | 31 (45.6) | 16 (44.4) | 15 (46.9) | 0.999 |
| Platinum resistant, n (%) | 37 (54.4) | 20 (55.6) | 17 (53.1) | |
| Treatment, n (%) | ||||
| Neoadjuvant chemotherapy | 40 (41.7) | 20 (32.8) | 20 (57.1) | 0.0309 * |
| “Surgery alone” and adjuvant chemotherapy | 56 (58.3) | 41 (67.2) | 15 (42.9) | |
| Death at the observation time point, n (%) | 37 (38.5) | 17 (27.9) | 20 (57.1) | 0.008 * |
| PFS, days [median (range)] | 413 (16–5055) | 971 (17–5055) | 388 (16–1266) | 0.004 * |
| OS, days [median (range)] | 776 (16–5055) | 1250 (17–5055) | 847 (16–3662) | 0.061 |
3.2. Impact of ADP Accumulation on HGSOC Prognosis
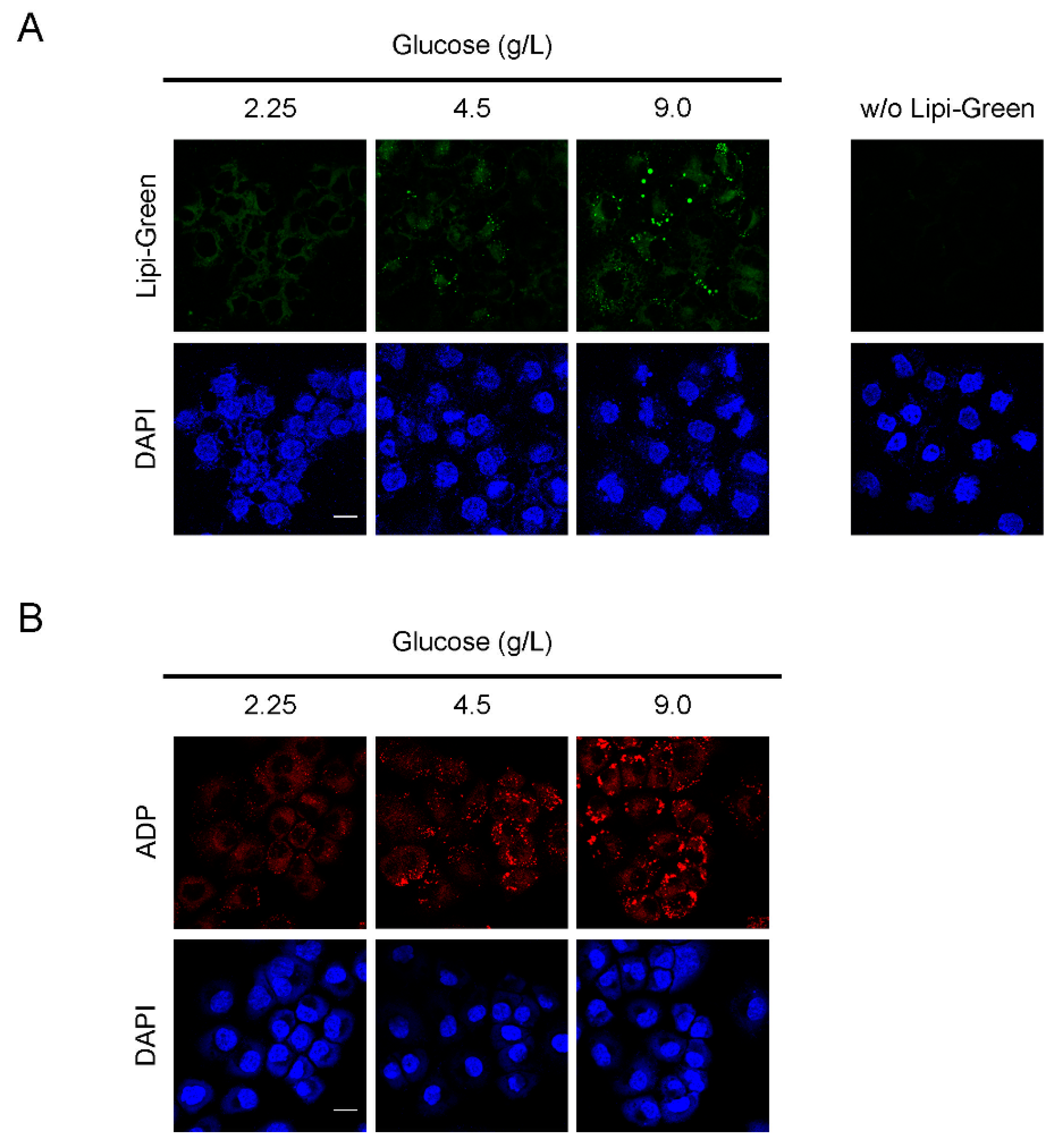
3.3. LD Formation and ADP Expression in OVCAR-3 Cells
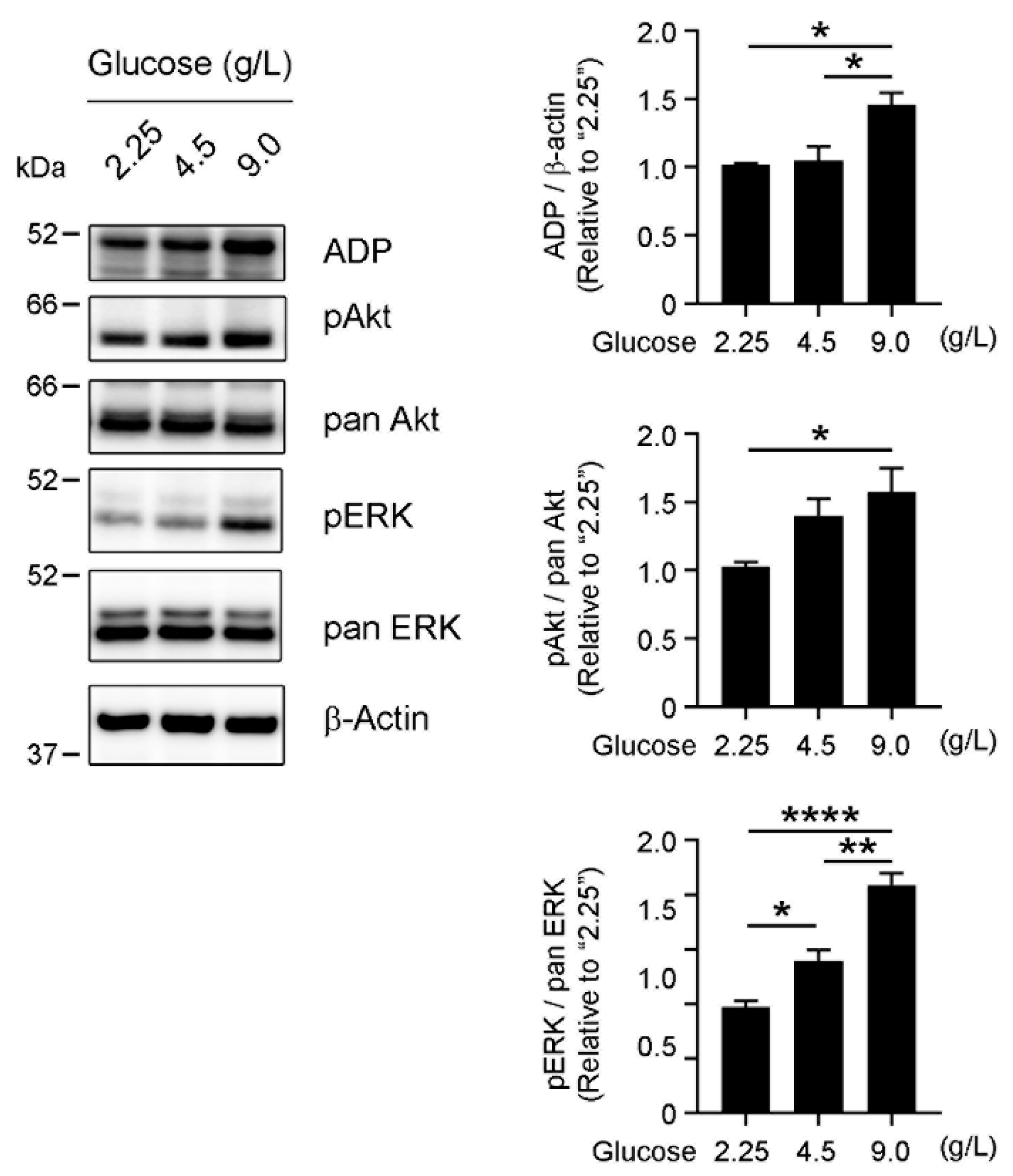
3.4. Glucose-Dependent Activation of Survival-Related Pathways in OVCAR-3 Cells
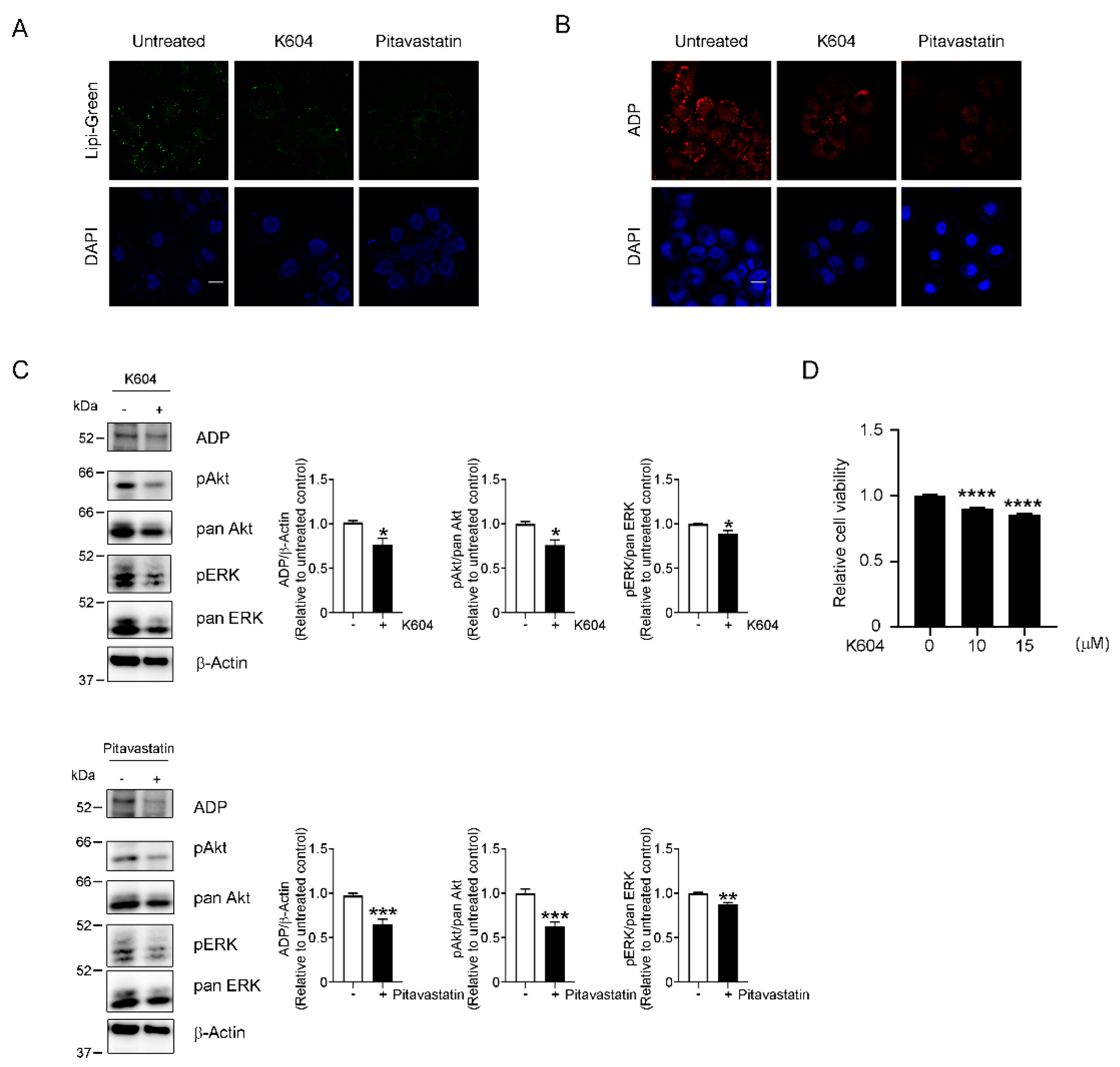
3.5. LD-Dependent Activation of Survival-Related Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Roffo, A.H. Heliotropism of Cholesterol in Relation to Skin Cancer. Am. J. Cancer 1933, 17, 42–57. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, M.; Bloor, W.R. Lipid content of tumors. J. Clin. Invest. 1932, 11, 677–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboumrad, M.H.; Horn, R.C., Jr.; Fine, G. Lipid-secreting mammary carcinoma. Report of a case associated with Paget’s disease of the nipple. Cancer 1963, 16, 521–525. [Google Scholar] [CrossRef]
- Wright, D.H. Lipid content of malignant lymphomas. J. Clin. Pathol. 1968, 21, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, B.; Ackerman, D.; Sanchez, D.J.; Li, B.; Ochocki, J.D.; Grazioli, A.; Bobrovnikova-Marjon, E.; Diehl, J.A.; Keith, B.; Simon, M.C. HIF2alpha-Dependent Lipid Storage Promotes Endoplasmic Reticulum Homeostasis in Clear-Cell Renal Cell Carcinoma. Cancer Discov. 2015, 5, 652–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebhard, R.L.; Clayman, R.V.; Prigge, W.F.; Figenshau, R.; Staley, N.A.; Reesey, C.; Bear, A. Abnormal cholesterol metabolism in renal clear cell carcinoma. J. Lipid Res. 1987, 28, 1177–1184. [Google Scholar] [CrossRef]
- Ostler, D.A.; Prieto, V.G.; Reed, J.A.; Deavers, M.T.; Lazar, A.J.; Ivan, D. Adipophilin expression in sebaceous tumors and other cutaneous lesions with clear cell histology: An immunohistochemical study of 117 cases. Mod. Pathol. 2010, 23, 567–573. [Google Scholar] [CrossRef] [Green Version]
- Milman, T.; Schear, M.J.; Eagle, R.C., Jr. Diagnostic utility of adipophilin immunostain in periocular carcinomas. Ophthalmology 2014, 121, 964–971. [Google Scholar] [CrossRef]
- Plaza, J.A.; Mackinnon, A.; Carrillo, L.; Prieto, V.G.; Sangueza, M.; Suster, S. Role of immunohistochemistry in the diagnosis of sebaceous carcinoma: A clinicopathologic and immunohistochemical study. Am. J. Dermatopathol. 2015, 37, 809–821. [Google Scholar] [CrossRef]
- Fujimoto, M.; Yoshizawa, A.; Sumiyoshi, S.; Sonobe, M.; Menju, T.; Hirata, M.; Momose, M.; Date, H.; Haga, H. Adipophilin expression in lung adenocarcinoma is associated with apocrine-like features and poor clinical prognosis: An immunohistochemical study of 328 cases. Histopathology 2017, 70, 232–241. [Google Scholar] [CrossRef]
- Fujimoto, M.; Matsuzaki, I.; Yamamoto, Y.; Yoshizawa, A.; Warigaya, K.; Iwahashi, Y.; Kojima, F.; Furukawa, F.; Murata, S.I. Adipophilin expression in cutaneous malignant melanoma. J. Cutan. Pathol. 2017, 44, 228–236. [Google Scholar] [CrossRef]
- Fujimoto, M.; Matsuzaki, I.; Nishitsuji, K.; Yamamoto, Y.; Murakami, D.; Yoshikawa, T.; Fukui, A.; Mori, Y.; Nishino, M.; Takahashi, Y.; et al. Adipophilin expression in cutaneous malignant melanoma is associated with high proliferation and poor clinical prognosis. Lab. Investig. 2020, 100, 727–737. [Google Scholar] [CrossRef]
- Conte, M.; Franceschi, C.; Sandri, M.; Salvioli, S. Perilipin 2 and Age-Related Metabolic Diseases: A New Perspective. Trends Endocrinol. Metab. 2016, 27, 893–903. [Google Scholar] [CrossRef]
- Straub, B.K.; Gyoengyoesi, B.; Koenig, M.; Hashani, M.; Pawella, L.M.; Herpel, E.; Mueller, W.; Macher-Goeppinger, S.; Heid, H.; Schirmacher, P. Adipophilin/perilipin-2 as a lipid droplet-specific marker for metabolically active cells and diseases associated with metabolic dysregulation. Histopathology 2013, 62, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, A.J.; Tudrej, P.; Kujawa, K.A.; Lisowska, K.M. Advances in ovarian cancer therapy. Cancer Chemother. Pharmacol. 2018, 81, 17–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Leroy, B.; Anderson, M.; Soussi, T. TP53 mutations in human cancer: Database reassessment and prospects for the next decade. Hum. Mutat. 2014, 35, 672–688. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Vang, R.; Shih Ie, M.; Kurman, R.J. Ovarian low-grade and high-grade serous carcinoma: Pathogenesis, clinicopathologic and molecular biologic features, and diagnostic problems. Adv. Anat. Pathol. 2009, 16, 267–282. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.K.; Davis, D.A.; Tomar, S.; Roy, L.; Gurler, H.; Xie, J.; Lantvit, D.D.; Cardenas, H.; Fang, F.; Liu, Y.; et al. In vivo tumor growth of high-grade serous ovarian cancer cell lines. Gynecol. Oncol. 2015, 138, 372–377. [Google Scholar] [CrossRef] [Green Version]
- Warburg, O. On respiratory impairment in cancer cells. Science 1956, 124, 269–270. [Google Scholar] [CrossRef]
- Hager, M.H.; Solomon, K.R.; Freeman, M.R. The role of cholesterol in prostate cancer. Curr. Opin. Clin. Nutr. Metab Care 2006, 9, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Tatenaka, Y.; Kato, H.; Ishiyama, M.; Sasamoto, K.; Shiga, M.; Nishitoh, H.; Ueno, Y. Monitoring Lipid Droplet Dynamics in Living Cells by Using Fluorescent Probes. Biochemistry 2019, 58, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Domcke, S.; Sinha, R.; Levine, D.A.; Sander, C.; Schultz, N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat. Commun. 2013, 4, 2126. [Google Scholar] [CrossRef] [PubMed]
- Altomare, D.A.; Testa, J.R. Perturbations of the AKT signaling pathway in human cancer. Oncogene 2005, 24, 7455–7464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, H.; Vikis, H.G.; Guan, K.L. Mechanisms of regulating the Raf kinase family. Cell Signal. 2003, 15, 463–469. [Google Scholar] [CrossRef]
- Ringerike, T.; Blystad, F.D.; Levy, F.O.; Madshus, I.H.; Stang, E. Cholesterol is important in control of EGF receptor kinase activity but EGF receptors are not concentrated in caveolae. J. Cell Sci. 2002, 115, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Lin, J.; Lu, M.L.; Solomon, K.R.; Freeman, M.R. Cholesterol-rich lipid rafts mediate akt-regulated survival in prostate cancer cells. Cancer Res. 2002, 62, 2227–2231. [Google Scholar]
- Liu, Y. Fatty acid oxidation is a dominant bioenergetic pathway in prostate cancer. Prostate Cancer Prostatic Dis. 2006, 9, 230–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikenoya, M.; Yoshinaka, Y.; Kobayashi, H.; Kawamine, K.; Shibuya, K.; Sato, F.; Sawanobori, K.; Watanabe, T.; Miyazaki, A. A selective ACAT-1 inhibitor, K-604, suppresses fatty streak lesions in fat-fed hamsters without affecting plasma cholesterol levels. Atherosclerosis 2007, 191, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Ohmoto, T.; Nishitsuji, K.; Yoshitani, N.; Mizuguchi, M.; Yanagisawa, Y.; Saito, H.; Sakashita, N. K604, a specific acylCoA:cholesterol acyltransferase 1 inhibitor, suppresses proliferation of U251MG glioblastoma cells. Mol. Med. Rep. 2015, 12, 6037–6042. [Google Scholar] [CrossRef] [PubMed]
- Siperstein, M.D.; Fagan, V.M. Deletion of the Cholesterol-Negative Feedback System in Liver Tumors. Cancer Res. 1964, 24, 1108–1115. [Google Scholar] [PubMed]
- Kajinami, K.; Takekoshi, N.; Saito, Y. Pitavastatin: Efficacy and safety profiles of a novel synthetic HMG-CoA reductase inhibitor. Cardiovasc. Drug Rev. 2003, 21, 199–215. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Ishikawa, E.; Odaka, H.; Miki, N.; Tawada, H.; Ikeda, H. TMP-153, a novel ACAT inhibitor, inhibits cholesterol absorption and lowers plasma cholesterol in rats and hamsters. Atherosclerosis 1995, 113, 71–78. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Odaka, H.; Itokawa, S.; Ishikawa, E.; Tomari, Y.; Ikeda, H. TMP-153, a novel ACAT inhibitor, lowers plasma cholesterol through its hepatic action in golden hamsters. Atherosclerosis 1995, 118, 145–153. [Google Scholar] [CrossRef]
- Ross, A.C.; Go, K.J.; Heider, J.G.; Rothblat, G.H. Selective inhibition of acyl coenzyme A:cholesterol acyltransferase by compound 58-035. J. Biol. Chem. 1984, 259, 815–819. [Google Scholar] [CrossRef]
- Tomoda, H.; Ito, M.; Tabata, N.; Masuma, R.; Yamaguchi, Y.; Omura, S. Amidepsines, inhibitors of diacylglycerol acyltransferase produced by Humicola sp. FO-2942. I. Production, isolation and biological properties. J. Antibiot. 1995, 48, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Tomoda, H.; Tabata, N.; Ito, M.; Omura, S. Amidepsines, inhibitors of diacylglycerol acyltransferase produced by Humicola sp. FO-2942. II. Structure elucidation of amidepsines A, B and C. J. Antibiot. 1995, 48, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Schulze, A.; Harris, A.L. How cancer metabolism is tuned for proliferation and vulnerable to disruption. Nature 2012, 491, 364–373. [Google Scholar] [CrossRef]
- Ji, Z.; Shen, Y.; Feng, X.; Kong, Y.; Shao, Y.; Meng, J.; Zhang, X.; Yang, G. Deregulation of Lipid Metabolism: The Critical Factors in Ovarian Cancer. Front. Oncol. 2020, 10, 593017. [Google Scholar] [CrossRef]
- Ward, P.S.; Thompson, C.B. Metabolic reprogramming: A cancer hallmark even warburg did not anticipate. Cancer Cell 2012, 21, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Kuhajda, F.P. Fatty-acid synthase and human cancer: New perspectives on its role in tumor biology. Nutrition 2000, 16, 202–208. [Google Scholar] [CrossRef]
- Menendez, J.A.; Lupu, R. Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat. Rev. Cancer 2007, 7, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Medes, G.; Thomas, A.; Weinhouse, S. Metabolism of neoplastic tissue. IV. A study of lipid synthesis in neoplastic tissue slices in vitro. Cancer Res. 1953, 13, 27–29. [Google Scholar] [PubMed]
- Altinkaya, S.O.; Nergiz, S.; Kucuk, M.; Yuksel, H. Apelin levels are higher in obese patients with endometrial cancer. J. Obstet. Gynaecol. Res. 2015, 41, 294–300. [Google Scholar] [CrossRef]
- Mitsiades, C.S.; Mitsiades, N.; Koutsilieris, M. The Akt pathway: Molecular targets for anti-cancer drug development. Curr. Cancer Drug Targets 2004, 4, 235–256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, X.; Li, C.; Wu, X.; Chen, Y.; Li, W.; Li, X.; Gong, F.; Tang, Q.; Jiang, D. SNCG promotes the progression and metastasis of high-grade serous ovarian cancer via targeting the PI3K/AKT signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 79. [Google Scholar] [CrossRef]
- Cheaib, B.; Auguste, A.; Leary, A. The PI3K/Akt/mTOR pathway in ovarian cancer: Therapeutic opportunities and challenges. Chin. J. Cancer 2015, 34, 4–16. [Google Scholar] [CrossRef]
- Adam, R.M.; Mukhopadhyay, N.K.; Kim, J.; Di Vizio, D.; Cinar, B.; Boucher, K.; Solomon, K.R.; Freeman, M.R. Cholesterol sensitivity of endogenous and myristoylated Akt. Cancer Res. 2007, 67, 6238–6246. [Google Scholar] [CrossRef] [Green Version]
- Reis-Sobreiro, M.; Roue, G.; Moros, A.; Gajate, C.; de la Iglesia-Vicente, J.; Colomer, D.; Mollinedo, F. Lipid raft-mediated Akt signaling as a therapeutic target in mantle cell lymphoma. Blood Cancer J. 2013, 3, e118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.; Pansino, F.; Baker, M.; Rice, G.; Quinn, M. Association between alphavbeta6 integrin expression, elevated p42/44 kDa MAPK, and plasminogen-dependent matrix degradation in ovarian cancer. J. Cell. Biochem. 2002, 84, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, R.; Wagoner, H.A.; Zeng, P.; Hammond, J.R.; Hannon, T.S.; Meyers, J.L.; Pescovitz, O.H. Mechanisms regulating the constitutive activation of the extracellular signal-regulated kinase (ERK) signaling pathway in ovarian cancer and the effect of ribonucleic acid interference for ERK1/2 on cancer cell proliferation. Mol. Endocrinol. 2004, 18, 2570–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Cheng, J.X. Direct visualization of de novo lipogenesis in single living cells. Sci. Rep. 2014, 4, 6807. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Bozza, P.T.; Tzizik, D.M.; Gray, J.P.; Cassara, J.; Dvorak, A.M.; Weller, P.F. Co-compartmentalization of MAP kinases and cytosolic phospholipase A2 at cytoplasmic arachidonate-rich lipid bodies. Am. J. Pathol. 1998, 152, 759–769. [Google Scholar]
- Yu, W.; Cassara, J.; Weller, P.F. Phosphatidylinositide 3-kinase localizes to cytoplasmic lipid bodies in human polymorphonuclear leukocytes and other myeloid-derived cells. Blood 2000, 95, 1078–1085. [Google Scholar] [CrossRef]
- Cruz, A.L.S.; Barreto, E.A.; Fazolini, N.P.B.; Viola, J.P.B.; Bozza, P.T. Lipid droplets: Platforms with multiple functions in cancer hallmarks. Cell Death Dis. 2020, 11, 105. [Google Scholar] [CrossRef] [Green Version]
- Butler, L.M.; Perone, Y.; Dehairs, J.; Lupien, L.E.; de Laat, V.; Talebi, A.; Loda, M.; Kinlaw, W.B.; Swinnen, J.V. Lipids and cancer: Emerging roles in pathogenesis, diagnosis and therapeutic intervention. Adv. Drug Deliv. Rev. 2020, 159, 245–293. [Google Scholar] [CrossRef]





| Parameter | PFS Univariate | PFS Multivariate | OS Univariate | OS Multivariate | ||||
|---|---|---|---|---|---|---|---|---|
| HR (95% CI) | p Value | HR (95% CI) | p Value | HR (95% CI) | p-Value | HR (95% CI) | p Value | |
| Patient age | 1.00 (0.98–1.02) | 0.8154 | 1.00 (0.98–1.02) | 0.9725 | 1.01 (0.98–1.04) | 0.6564 | 1.01 (0.98–1.04) | 0.5481 |
| BMI | 1.02 (0.96–1.07) | 0.5684 | 1.00 (0.94–1.05) | 0.9447 | 1.02 (0.95–1.09) | 0.5134 | 1.00 (0.93–1.07) | 0.9483 |
| Stage (I-II vs. III-IV) | 6.28 (1.54–25.70) | 0.0005 * | 5.58 (1.35–23.01) | 0.0017 * | 2.43 (0.58–10.09) | 0.1630 | 1.98 (0.47–8.34) | 0.3058 |
| NAC (+ vs. −) | 1.62 (0.99–2.62) | 0.0523 | 1.06 (0.63–1.77) | 0.8238 | 1.49 (0.78–2.84) | 0.2289 | 1.09 (0.55–2.18) | 0.8016 |
| ADP (high vs. low) | 2.15 (1.30–3.54) | 0.0027 * | 1.96 (1.17–3.27) | 0.0098 * | 2.24 (1.15–4.35) | 0.0158 * | 2.18 (1.10–4.34) | 0.0245 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwahashi, N.; Ikezaki, M.; Fujimoto, M.; Komohara, Y.; Fujiwara, Y.; Yamamoto, M.; Mizoguchi, M.; Matsubara, K.; Watanabe, Y.; Matsuzaki, I.; et al. Lipid Droplet Accumulation Independently Predicts Poor Clinical Prognosis in High-Grade Serous Ovarian Carcinoma. Cancers 2021, 13, 5251. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205251
Iwahashi N, Ikezaki M, Fujimoto M, Komohara Y, Fujiwara Y, Yamamoto M, Mizoguchi M, Matsubara K, Watanabe Y, Matsuzaki I, et al. Lipid Droplet Accumulation Independently Predicts Poor Clinical Prognosis in High-Grade Serous Ovarian Carcinoma. Cancers. 2021; 13(20):5251. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205251
Chicago/Turabian StyleIwahashi, Naoyuki, Midori Ikezaki, Masakazu Fujimoto, Yoshihiro Komohara, Yukio Fujiwara, Madoka Yamamoto, Mika Mizoguchi, Kentaro Matsubara, Yudai Watanabe, Ibu Matsuzaki, and et al. 2021. "Lipid Droplet Accumulation Independently Predicts Poor Clinical Prognosis in High-Grade Serous Ovarian Carcinoma" Cancers 13, no. 20: 5251. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205251