
Mutation in Genes Encoding Key Functional Groups Additively Increase Mortality in Patients with BRAFV600E-Mutant Advanced Papillary Thyroid Carcinoma

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Patients and Tissue Samples
2.2. DNA Extraction and Preparation
2.3. Targeted Next-Generated Sequencing (NGS) and Analysis Process
2.4. Functional Groups and Patient Grouping
2.5. Primary Outcome
2.6. Statistical Analysis
3. Results
3.1. Clinical Characteristics of Patients
3.2. Mutational Profile
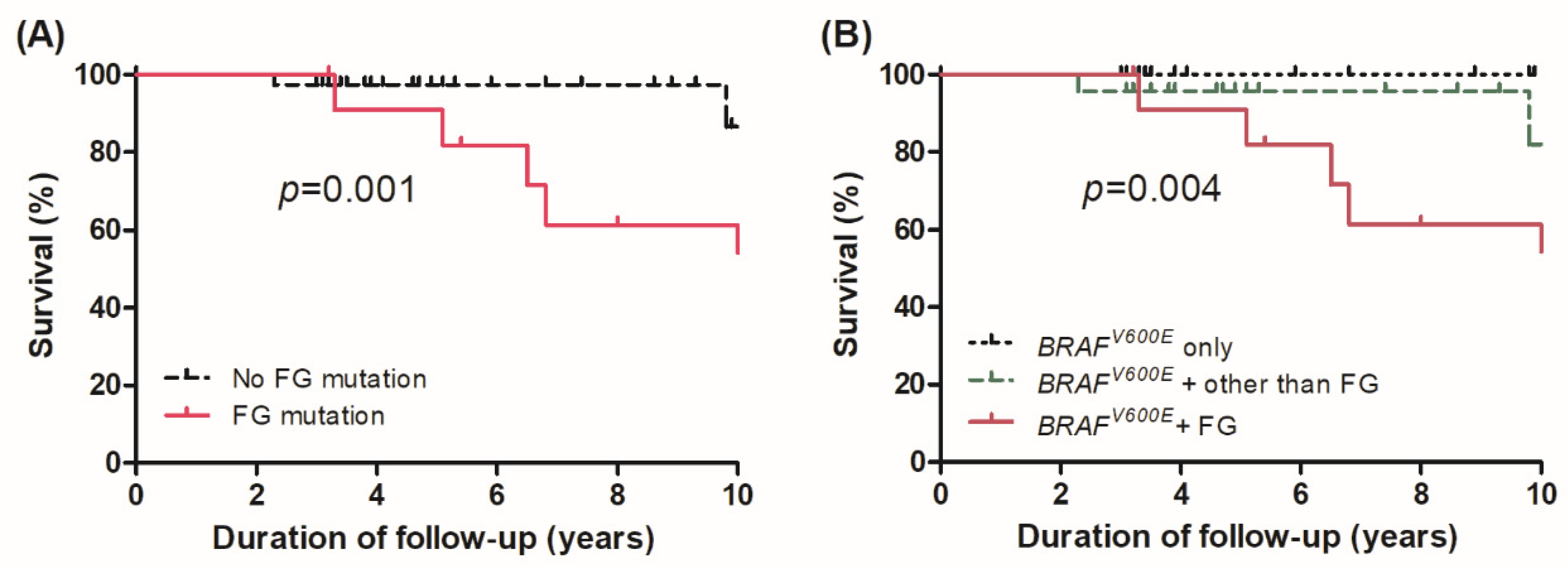
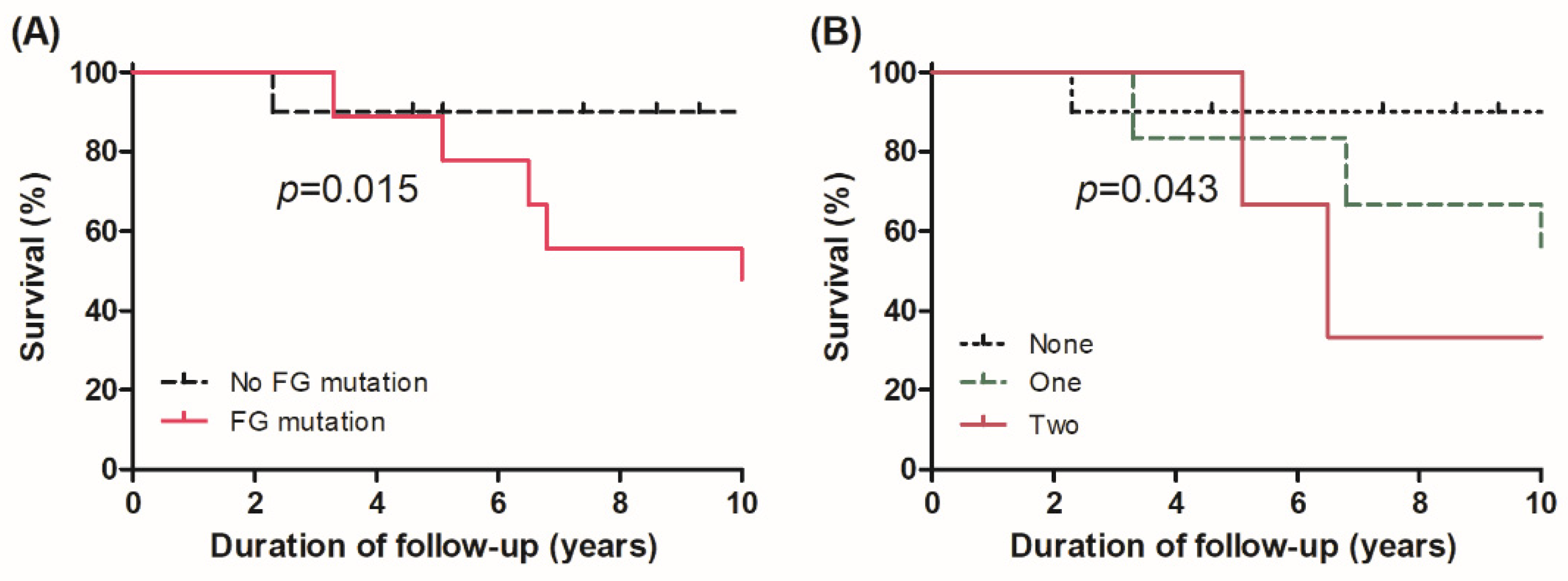
3.3. Survival in Different Genomic Subgroups
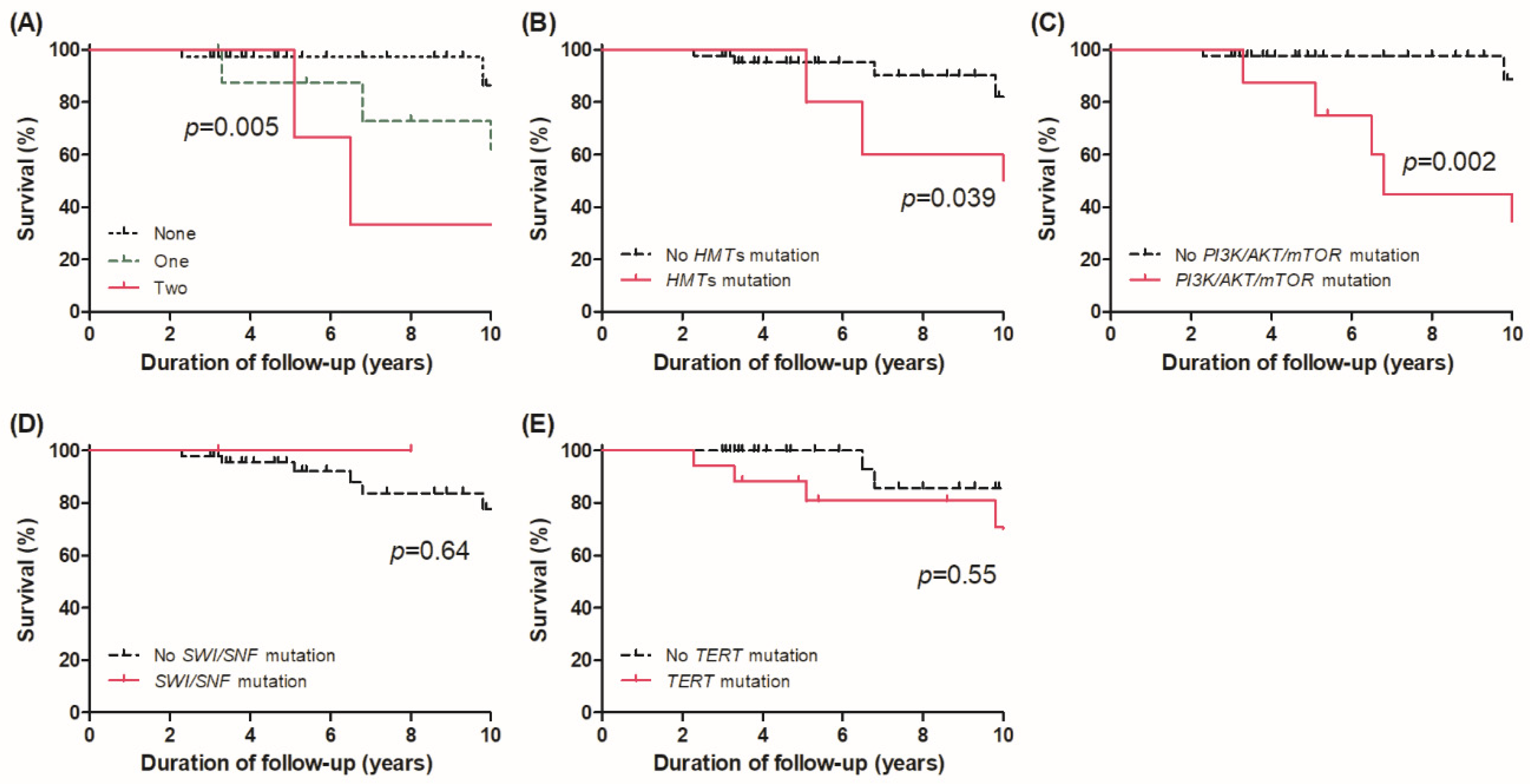
3.4. Effect of Alterations in FGs on Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xing, M.; Liu, R.; Liu, X.; Murugan, A.K.; Zhu, G.; Zeiger, M.A.; Pai, S.; Bishop, J. BRAF V600E and TERT promoter mutations cooperatively identify the most aggressive papillary thyroid cancer with highest recurrence. J. Clin. Oncol. 2014, 32, 2718–2726. [Google Scholar] [CrossRef] [Green Version]
- Fagin, J.A.; Wells, S.A., Jr. Biologic and Clinical Perspectives on Thyroid Cancer. N. Engl. J. Med. 2016, 375, 1054–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, M.; da Rocha, A.G.; Vinagre, J.; Batista, R.; Peixoto, J.; Tavares, C.; Celestino, R.; Almeida, A.; Salgado, C.; Eloy, C.; et al. TERT promoter mutations are a major indicator of poor outcome in differentiated thyroid carcinomas. J. Clin. Endocrinol. Metab. 2014, 99, E754–E765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, E.T.; Nikiforova, M.N.; Zhu, Z.; Knauf, J.A.; Nikiforov, Y.E.; Fagin, J.A. High prevalence of BRAF mutations in thyroid cancer: Genetic evidence for constitutive activation of the RET/PTC-RAS-BRAF signaling pathway in papillary thyroid carcinoma. Cancer Res. 2003, 63, 1454–1457. [Google Scholar]
- Xing, M.; Westra, W.H.; Tufano, R.P.; Cohen, Y.; Rosenbaum, E.; Rhoden, K.J.; Carson, K.A.; Vasko, V.; Larin, A.; Tallini, G.; et al. BRAF mutation predicts a poorer clinical prognosis for papillary thyroid cancer. J. Clin. Endocrinol. Metab. 2005, 90, 6373–6379. [Google Scholar] [CrossRef] [Green Version]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Viola, D.; Elisei, R.; Bendlova, B.; Yip, L.; Mian, C.; Vianello, F.; Tuttle, M.R.; et al. Association between BRAF V600E mutation and mortality in patients with papillary thyroid cancer. JAMA 2013, 309, 1493–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzaferri, E.L.; Jhiang, S.M. Long-term impact of initial surgical and medical therapy on papillary and follicular thyroid cancer. Am. J. Med. 1994, 97, 418–428. [Google Scholar] [CrossRef]
- Ito, Y.; Miyauchi, A.; Kihara, M.; Fukushima, M.; Higashiyama, T.; Miya, A. Overall Survival of Papillary Thyroid Carcinoma Patients: A Single-Institution Long-Term Follow-Up of 5897 Patients. World J. Surg. 2018, 42, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.Y.; Kim, W.G.; Kim, W.B.; Shong, Y.K. Current status and future perspectives in differentiated thyroid cancer. Endocrinol. Metab. 2014, 29, 217–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landa, I.; Ibrahimpasic, T.; Boucai, L.; Sinha, R.; Knauf, J.A.; Shah, R.H.; Dogan, S.; Ricarte-Filho, J.C.; Krishnamoorthy, G.P.; Xu, B.; et al. Genomic and transcriptomic hallmarks of poorly differentiated and anaplastic thyroid cancers. J. Clin. Investig. 2016, 126, 1052–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, E.; Song, D.E.; Ahn, J.; Kim, T.Y.; Kim, W.B.; Shong, Y.K.; Jeon, M.J.; Kim, W.G. Genetic profile of advanced thyroid cancers in relation to distant metastasis. Endocr. Relat. Cancer 2020, 27, 285–293. [Google Scholar] [CrossRef]
- Jin, M.; Song, D.E.; Ahn, J.; Song, E.; Lee, Y.M.; Sung, T.Y.; Kim, T.Y.; Kim, W.B.; Shong, Y.K.; Jeon, M.J.; et al. Genetic Profiles of Aggressive Variants of Papillary Thyroid Carcinomas. Cancers 2021, 13, 892. [Google Scholar] [CrossRef]
- Agrawal, N.; Akbani, R.; Aksoy, B.A.; Ally, A.; Arachchi, H.; Asa, S.L.; Auman, J.T.; Balasundaram, M.; Balu, S.; Baylin, S.B.; et al. Integrated genomic characterization of papillary thyroid carcinoma. Cell 2014, 159, 676–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahimpasic, T.; Xu, B.; Landa, I.; Dogan, S.; Middha, S.; Seshan, V.; Deraje, S.; Carlson, D.L.; Migliacci, J.; Knauf, J.A.; et al. Genomic Alterations in Fatal Forms of Non-Anaplastic Thyroid Cancer: Identification of MED12 and RBM10 as Novel Thyroid Cancer Genes Associated with Tumor Virulence. Clin. Cancer Res. 2017, 23, 5970–5980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, M. BRAF mutation in thyroid cancer. Endocr. Relat. Cancer 2005, 12, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.H.; Park, Y.J.; Lim, J.A.; Ahn, H.Y.; Lee, E.K.; Lee, Y.J.; Kim, K.W.K.; Hahn, S.K.; Youn, Y.K.; Kim, K.H.; et al. The association of the BRAF(V600E) mutation with prognostic factors and poor clinical outcome in papillary thyroid cancer: A meta-analysis. Cancer 2012, 118, 1764–1773. [Google Scholar] [CrossRef] [PubMed]
- Saqcena, M.; Leandro-Garcia, L.J.; Maag, J.L.V.; Tchekmedyian, V.; Krishnamoorthy, G.P.; Tamarapu, P.P.; Tiedje, V.; Reuter, V.; Knauf, J.A.; de Stanchina, E.; et al. SWI/SNF Complex Mutations Promote Thyroid Tumor Progression and Insensitivity to Redifferentiation Therapies. Cancer Discov. 2021, 11, 1158–1175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hou, P.; Ji, M.; Guan, H.; Studeman, K.; Jensen, K.; Casko, V.; El-Naggar, A.K.; Xing, M. Highly prevalent genetic alterations in receptor tyrosine kinases and phosphatidylinositol 3-kinase/akt and mitogen-activated protein kinase pathways in anaplastic and follicular thyroid cancers. J. Clin. Endocrinol. Metab. 2008, 93, 3106–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricarte-Filho, J.C.; Ryder, M.; Chitale, D.A.; Rivera, M.; Heguy, A.; Ladanyi, M.; Janakiraman, M.; Solit, D.; Knauf, J.A.; Tuttle, R.M.; et al. Mutational profile of advanced primary and metastatic radioactive iodine-refractory thyroid cancers reveals distinct pathogenetic roles for BRAF, PIK3CA, and AKT1. Cancer Res. 2009, 69, 4885–4893. [Google Scholar] [CrossRef] [Green Version]
- Pappa, T.; Ahmadi, S.; Marqusee, E.; Johnson, H.L.; Nehs, M.A.; Cho, N.L.; Barletta, J.A.; Lorch, J.H.; Doherty, G.M.; Lindeman, N.I.; et al. Oncogenic Mutations in PI3K/AKT/mTOR Pathway Effectors Associate with Worse Prognosis in BRAF(V600E)-Driven Papillary Thyroid. Cancer Patients Clin. Cancer Res. 2021, 27, 4256–4264. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.S.; Park, Y.J. Mechanisms of TERT Reactivation and Its Interaction with BRAFV600E. Endocrinol. Metab. 2020, 35, 515–525. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total (n = 50) | Mutations in FGs (−) (n = 37) | Mutations in FGs (+) (n = 13) | p-Value |
|---|---|---|---|---|
| Pathology | 0.026 | |||
| Classic PTC | 19 (38.0%) | 10 (27.0%) | 9 (69.2%) | |
| TV PTC | 23 (46.0%) | 20 (54.1%) | 3 (23.1%) | |
| CCV PTC | 8 (16.0%) | 7 (18.9%) | 1 (7.7%) | |
| Age (years) | 49.5 (38.0–59.5) | 48.0 (35.0–55.0) | 58.9 (48.9–68.4) | 0.025 |
| Sex (female) | 29 (58.0%) | 23 (62.2%) | 6 (46.2%) | 0.497 |
| Primary tumor size (cm) | 2.3 (1.5–3.2) | 2.1 (1.4–2.8) | 3.0 (1.5–3.8) | 0.341 |
| Extrathyroidal extension | 0.535 | |||
| Microscopic | 23 (46%) | 17 (45.9%) | 6 (46.2%) | |
| Gross | 19 (38%) | 15 (40.5%) | 4 (30.8%) | |
| Multifocality (yes) | 21 (42.0%) | 16 (43.2%) | 5 (38.5%) | 0.801 |
| LN metastases (yes) | 0.482 | |||
| N1a | 15 (30.0%) | 12 (32.4%) | 3 (23.1%) | |
| N1b | 30 (60.0%) | 22 (59.4%) | 8 (61.5%) | |
| Distant metastases | 0.017 | |||
| Synchronous | 6 (12.0%) | 4 (10.8%) | 2 (15.4%) | |
| Metachronous | 13 (26.0%) | 6 (16.2%) | 7 (53.8%) | |
| AJCC TNM 8th stage | 0.248 | |||
| I | 33 (66.0%) | 27 (73.0%) | 6 (46.2%) | |
| II | 12 (24.0%) | 7 (18.9%) | 5 (38.5%) | |
| III | 1 (2.0%) | 1 (2.7%) | 0 (0.0%) | |
| IV | 4 (8.0%) | 2 (5.4%) | 2 (15.4%) | |
| Follow-up duration (years) | 5.3 (3.4–9.8) | 4.9 (3.4–9.3) | 6.8 (5.1–13.6) | 0.219 |
| Overall mortality | 11 (22.0%) | 3 (8.1%) | 8 (61.5%) | <0.001 |
| Risk Factors | Adjusted HR (95% CI) * | p-Value |
|---|---|---|
| Mutation in functional groups | ||
| Presence vs. absence | 11.65 (1.39–97.58) | 0.024 |
| Number of mutations in functional groups | ||
| One site vs. none | 11.53 (2.69–49.4) | <0.001 |
| Two sites vs. none | 60.12 (10.63–340.2) | <0.001 |
| Comparisons in groups | ||
| Group 2 vs. Group 1 | 1.20 (0.14–10.04) | 0.867 |
| Group 3 vs. Group 1 | 13.96 (1.67–116.9) | 0.015 |
| Group 3 vs. Group 2 | 11.65 (1.39–97.58) | 0.024 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, E.; Jin, M.; Jang, A.; Jeon, M.J.; Song, D.E.; Yoo, H.J.; Kim, W.B.; Shong, Y.K.; Kim, W.G. Mutation in Genes Encoding Key Functional Groups Additively Increase Mortality in Patients with BRAFV600E-Mutant Advanced Papillary Thyroid Carcinoma. Cancers 2021, 13, 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13225846
Song E, Jin M, Jang A, Jeon MJ, Song DE, Yoo HJ, Kim WB, Shong YK, Kim WG. Mutation in Genes Encoding Key Functional Groups Additively Increase Mortality in Patients with BRAFV600E-Mutant Advanced Papillary Thyroid Carcinoma. Cancers. 2021; 13(22):5846. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13225846
Chicago/Turabian StyleSong, Eyun, Meihua Jin, Ahreum Jang, Min Ji Jeon, Dong Eun Song, Hye Jin Yoo, Won Bae Kim, Young Kee Shong, and Won Gu Kim. 2021. "Mutation in Genes Encoding Key Functional Groups Additively Increase Mortality in Patients with BRAFV600E-Mutant Advanced Papillary Thyroid Carcinoma" Cancers 13, no. 22: 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13225846