
Chromosomal Rearrangements and Altered Nuclear Organization: Recent Mechanistic Models in Cancer

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
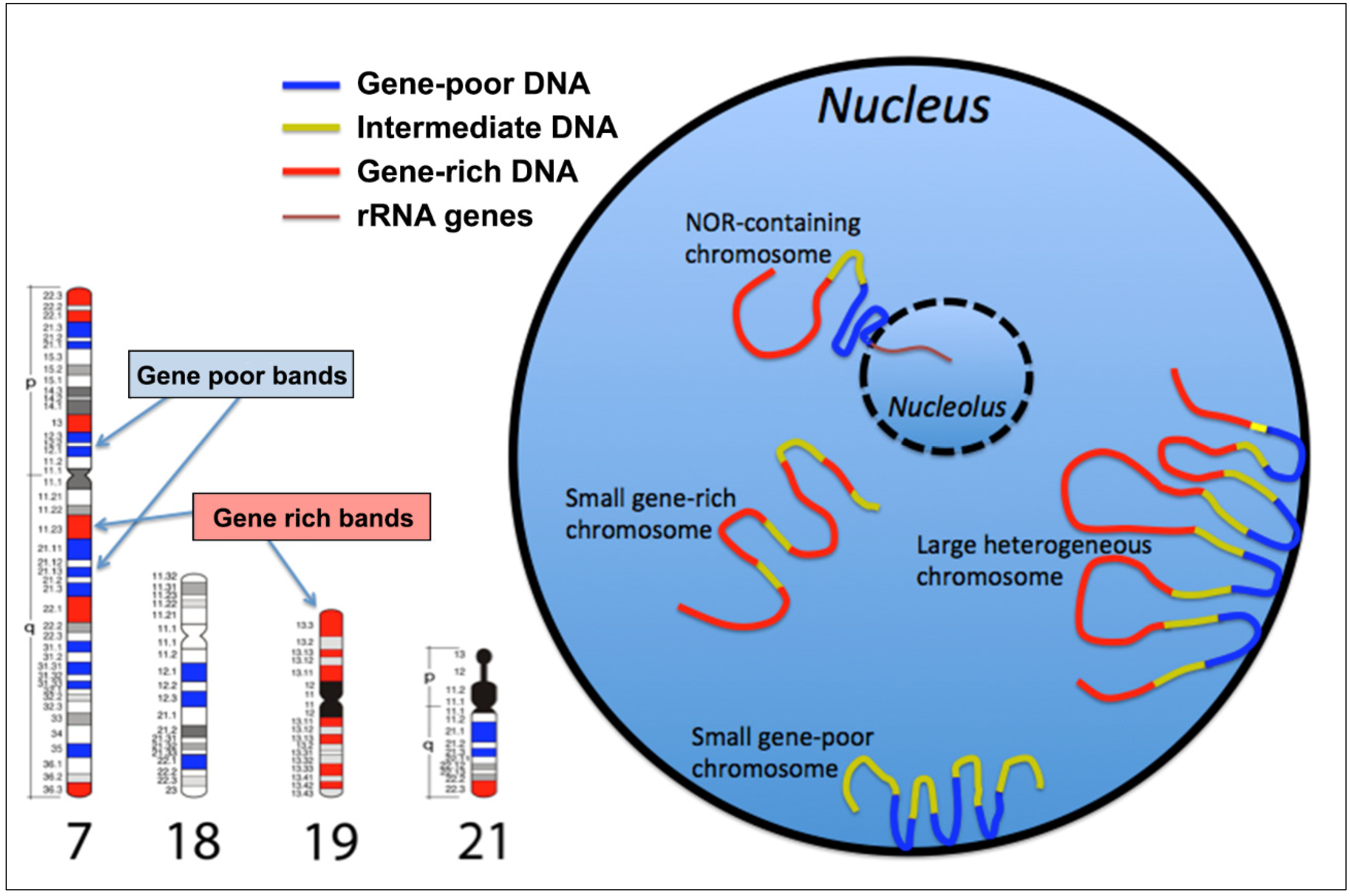
2. The Chromosome Organization in the Cell Nuclei
3. Genome Organization inside the Chromosome Territories
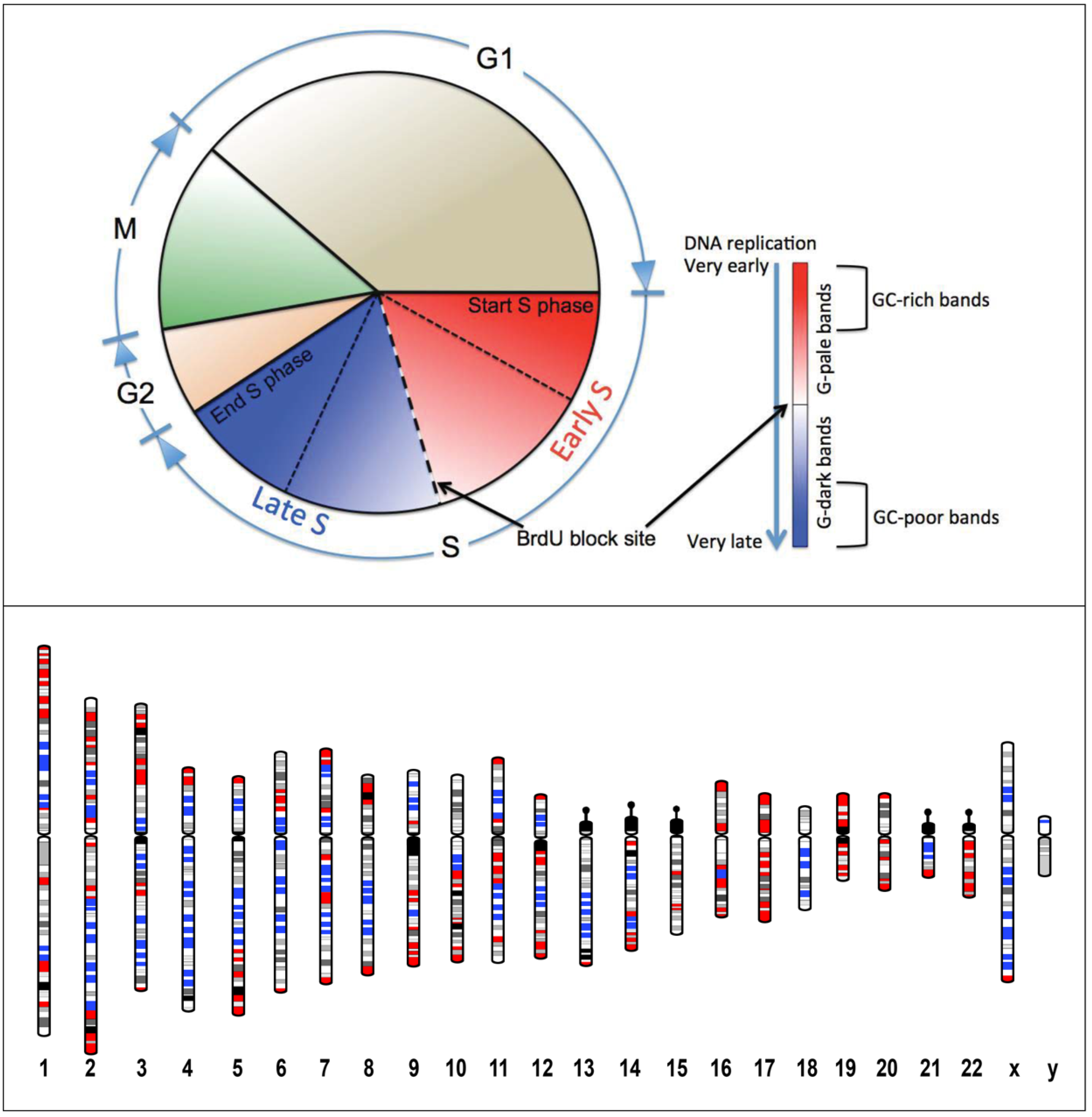
4. Nuclear Architecture, Replication Timing, and Cancer Diseases
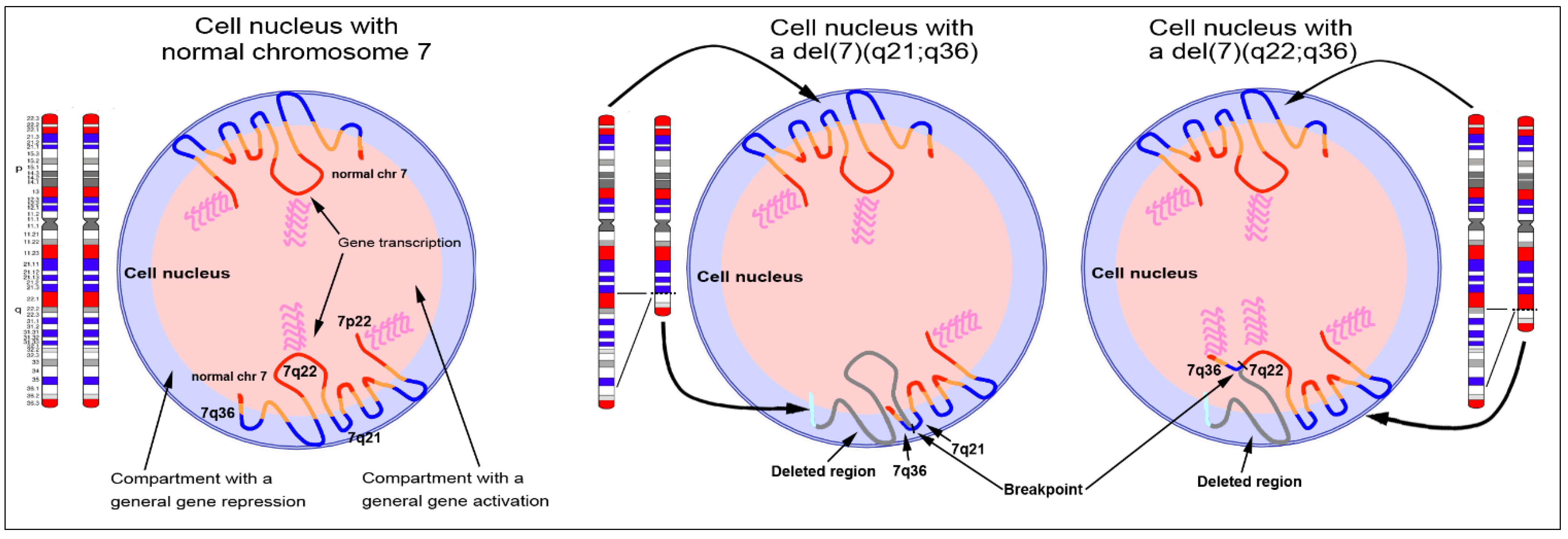
5. Chromosomal Rearrangements and Nuclear Gene Repositioning in Cancers
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rocha, E.P.C. The Organization of the Bacterial Genome. Annu. Rev. Genet. 2008, 42, 211–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridger, J.M.; Arican-Gotkas, H.D.; Foster, H.A.; Godwin, L.S.; Harvey, A.; Kill, I.R.; Knight, M.; Mehta, I.S.; Ahmed, M.H. The Non-random Repositioning of Whole Chromosomes and Individual Gene Loci in Interphase Nuclei and Its Relevance in Disease, Infection, Aging, and Cancer. Adv. Exp. Med. Biol. 2014, 263–279. [Google Scholar]
- Boyle, S.; Gilchrist, S.; Bridger, J.M.; Mahy, N.L.; Ellis, J.A.; Bickmore, W.A. The spatial organization of human chromosomes within the nuclei of normal and emerin-mutant cells. Hum. Mol. Genet. 2001, 10, 211–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolzer, A.; Kreth, G.; Solovei, I.; Koehler, D.; Saracoglu, K.; Fauth, C.; Müller, S.; Eils, R.; Cremer, C.; Speicher, M.R.; et al. Three dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol. 2005, 3, 826–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccone, S.; Federico, C.; Bernardi, G. Localization of the gene-richest and the gene poorest isochores in the interphase nuclei of mammals and birds. Gene 2002, 300, 169–178. [Google Scholar] [CrossRef]
- Federico, C.; Cantarella, C.D.; Di Mare, P.; Tosi, S.; Saccone, S. The radial arrangement of the human chromosome 7 in the lymphocyte cell nucleus is associated with chromosomal band gene density. Chromosoma 2008, 117, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Meaburn, K.J.; Newbold, R.F.; Bridger, J.M. Positioning of human chromosomes in murine cell hybrids according to synteny. Chromosoma 2008, 117, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Mayer, R.; Brero, A.; von Hase, J.; Schroeder, T.; Cremer, T.; Dietzel, S. Common themes and cell type specific variations of higher order chromatin arrangements in the mouse. BMC Cell Biol. 2005, 6, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, H.; Mùller, S.; Neusser, M.; von Hase, J.; Calcagno, E.; Cremer, M.; Solovei, S.; Cremer, C.; Cremer, T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc. Natl. Acad. Sci. USA 2002, 99, 4424–4429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, C.; Pappalardo, A.M.; Ferrito, V.; Tosi, S.; Saccone, S. Genomic properties of chromosomal bands are linked to evolutionary rearrangements and new centromere formation in primates. Chromosome Res. 2017, 25, 261–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kociucka, B.; Sosnowski, J.; Kubiak, A.; Nowak, A.; Pawlak, P.; Szczerbal, I. Three-Dimensional Positioning of B Chromosomes in Fibroblast Nuclei of the Red Fox and the Chinese Raccoon Dog. Cytogenet. Genome Res. 2013, 139, 243–249. [Google Scholar]
- Foster, H.A.; Griffin, D.K.; Bridger, J.M. Interphase chromosome positioning in in vitro porcine cells and ex vivo porcine tissues. BMC Cell Biol. 2012, 13, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, C.; Saccone, S.; Andreozzi, L.; Motta, S.; Russo, V.; Carels, N.; Bernardi, G. The pig genome: Compositional analysis and identification of the gene-richest regions in chromosomes and nuclei. Gene 2004, 34, 245–251. [Google Scholar] [CrossRef]
- Habermann, F.A.; Cremer, M.; Walter, J.; Kreth, G.; von Hase, J.; Bauer, K.; Wienberg, J.; Cremer, C.; Cremer, T.; Solovei, I. Arrangements of macro- and microchromosomes in chicken cells. Chromosome Res. 2001, 9, 569–584. [Google Scholar] [CrossRef]
- Federico, C.; Cantarella, C.D.; Scavo, C.; Saccone, S.; Bed’Hom, B.; Bernardi, G. Avian genomes: Different karyotypes but a similar distribution of the GC-richest chromosome regions at interphase. Chromosome Res. 2005, 3, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Federico, C.; Scavo, C.; Cantarella, C.D.; Motta, S.; Saccone, S.; Bernardi, G. Gene-rich and gene-poor chromosomal regions have different locations in the interphase nuclei of cold-blooded vertebrates. Chromosoma 2006, 115, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Croft, J.; Bridger, J.M.; Boyle, S.; Perry, P.; Teague, P.; Bickmore, W.A. Differences in the localization and morphology of chromosomes in the human nucleus. J. Cell Biol. 1999, 145, 1119–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, H.A.; Abeydeera, L.R.; Griffin, D.K.; Bridger, J.M. Non-random chromosome positioning in mammalian sperm nuclei, with migration of the sex chromosomes during late spermatogenesis. J. Cell Sci. 2005, 118, 1811–1820. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J.; Rippe, K.; Dekker, M.; Kleckner, N. Capturing Chromosome Conformation. Science 2002, 295, 1306–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guelen, L.; Pagie, L.; Brasset, E.; Meuleman, W.; Faza, M.B.; Talhout, W.; Eussen, B.H.; De Klein, A.; Wessels, L.; De Laat, W.; et al. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 2008, 453, 948–951. [Google Scholar] [CrossRef] [PubMed]
- Bourne, G.; Moir, C.; Bikkul, U.; Ahmed Hassan, M.; Kill, I.R.; Eskiw, C.H.; Tosi, S. Interphase chromosome behavior in normal and diseased cells. In Human Interphase Chromosomes; Yurov, Y., Vorsanova, S., Iourov, I., Eds.; Springer: New York, NY, USA, 2013; pp. 9–33. [Google Scholar]
- Xing, Y.; Johnson, C.V.; Moen, P.T.; McNeil, J.A.; Lawrence, J. Nonrandom gene organization: Structural arrangements of specific pre-mRNA transcription and splicing with SC-35 domains. J. Cell Biol. 1995, 131, 1635–1647. [Google Scholar] [CrossRef] [Green Version]
- Mahy, N.L.; Perry, P.E.; Gilchrist, S.; Baldock, R.A.; Bickmore, W.A. Spatial organization of active and inactive genes and noncoding DNA within chromosome territories. J. Cell Biol. 2002, 157, 579–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Antoni, S.; Mattina, T.; Di Mare, P.; Federico, C.; Motta, S.; Saccone, S. Altered replication timing of the HIRA/Tuple1 locus in the DiGeorge and Velocardiofacial syndromes. Gene 2004, 333, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Ballabio, E.; Cantarella, C.D.; Federico, C.; Di Mare, P.; Hall, G.; Harbott, J.; Hughes, J.; Saccone, S.; Tosi, S. Ectopic expression of the HLXB9 gene is associated with an altered nuclear position in t(7;12) leukaemias. Leukemia 2009, 23, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Leotta, C.G.; Federico, C.; Brundo, M.V.; Tosi, S.; Saccone, S. HLXB9 gene expression, and nuclear location during in vitro neuronal differentiation in the SK-N-BE neuroblastoma cell line. PLoS ONE 2014, 9, e105481. [Google Scholar]
- Gulino, G.M.; Bruno, F.; Sturiale, V.; Brancato, D.; Ragusa, D.; Tosi, S.; Saccone, S.; Federico, C. From FISH to Hi-C: The chromatin architecture of the chromosomal region 7q36.3, frequently rearranged in leukemic cells, is evolutionary conserved. Int. J. Mol. Sci. 2021, 22, 2338. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Foster, H.A.; Bridger, J.M. The spatial repositioning of adipogenesis genes is correlated with their expression status in a porcine mesenchymal stem cell adipogenesis model system. Chromosoma 2009, 118, 647–663. [Google Scholar] [CrossRef]
- Szczerbal, I.; Bridger, J.M. Association of adipogenic genes with SC-35 domains during porcine adipogenesis. Chromosome Res. 2010, 18, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Arican-Goktas, H.D.; Ittiprasert, W.; Bridger, J.M.; Knight, M. Differential Spatial Repositioning of Activated Genes in Biomphalaria glabrata Snails Infected with Schistosoma mansoni. PLoS Negl. Trop. Dis. 2014, 8, e3013. [Google Scholar] [CrossRef] [Green Version]
- Cremer, T.; Cremer, C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2001, 2, 292–301. [Google Scholar] [CrossRef]
- Malhas, A.; Lee, C.F.; Sanders, R.; Saunders, N.J.; Vaux, D.J. Defects in lamin B1 expression or processing affect interphase chromosome position and gene expression. J. Cell Biol. 2007, 176, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Saccone, S.; De Sario, A.; Della Valle, G.; Bernardi, G. The highest gene concentrations in the human genome are in telomeric bands of metaphase chromosomes. Proc. Natl. Acad. Sci. USA 1992, 89, 4913–4917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccone, S.; De Sario, A.; Wiegant, J.; Raap, A.K.; Della Valle, G.; Bernardi, G. Correlations between isochores and chromosomal bands in the human genome. Proc. Natl. Acad. Sci. USA 1993, 90, 11929–11933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccone, S.; Federico, C.; Solovei, I.; Croquette, M.F.; Della Valle, G.; Bernardi, G. Identification of the gene-richest bands in human prometaphase chromosomes. Chromosome Res. 1999, 7, 379–386. [Google Scholar] [CrossRef]
- Federico, C.; Andreozzi, L.; Saccone, S.; Bernardi, G. Gene density in the Giemsa bands of human chromosomes. Chromosome Res. 2000, 8, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Bickmore, W.A.; van Steensel, B. Genome Architecture: Domain Organization of Interphase Chromosomes. Cell 2013, 152, 1270–1284. [Google Scholar] [CrossRef] [Green Version]
- Simonis, M.; Klous, P.; Splinter, E.; Moshkin, Y.; Willemsen, R.; de Wit, E.; van Steensel, B.; de Laat, W. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C). Nat. Genet. 2006, 38, 1348–1354. [Google Scholar] [CrossRef]
- Dostie, J.; Richmond, T.A.; Arnaout, R.A.; Selzer, R.R.; Lee, W.L.; Honan, T.A.; Rubio, E.D.; Krumm, A.; Lamb, J.; Nusbaum, C.; et al. Chromosome Conformation Capture Carbon Copy (5C): A massively parallel solution for mapping interactions between genomic elements. Genome Res. 2006, 16, 1299–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieberman-Aiden, E.; van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive Mapping of Long-Range Interactions Reveals Folding Principles of the Human Genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belton, J.-M.; McCord, R.P.; Gibcus, J.H.; Naumova, N.; Zhan, Y.; Dekker, J. Hi-C: A comprehensive technique to capture the conformation of genomes. Methods 2012, 58, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shavit, Y.; Lio’, P. CytoHiC: A cytoscape plugin for visual comparison of Hi-C networks. Bioinformatics 2013, 29, 1206–1207. [Google Scholar] [CrossRef] [Green Version]
- Botta, M.; Haider, S.; Leung, I.X.Y.; Lio’, P.; Mozziconacci, J. Intra- and inter-chromosomal interactions correlate with CTCF binding genome wide. Mol. Syst. Biol. 2010, 6, 426. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, K.; Bernardi, G. An Isochore Framework Underlies Chromatin Architecture. PLoS ONE 2017, 12, e0168023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anania, C.; Lupianez, D.G. Order and disorder: Abnormal 3D chromatin organization in human disease. Brief. Funct. Genom. 2020, 19, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Huang, S. CTCF mediated genome organization and leukemogenesis. Leukemia 2020, 34, 2295–2304. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Su, J.-H.; Beliveau, B.J.; Bintu, B.; Moffitt, J.R.; Wu, C.; Zhuang, X. Spatial organization of chromatin domains and compartments in single chromosomes. Science 2016, 353, 598–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, T.J.; Lando, D.; Basu, S.; Atkinson, L.P.; Cao, Y.; Lee, S.F.; Leeb, M.; Wohlfahrt, K.J.; Boucher, W.; O’Shaughnessy-Kirwan, A.; et al. 3D structures of individual mammalian genomes studied by single-cell Hi-C. Nature 2017, 544, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, G. Chromosome Architecture and Genome Organization. PLoS ONE 2015, 10, e0143739. [Google Scholar] [CrossRef] [Green Version]
- Kurz, A.; Lampel, S.; Nickolenko, J.E.; Bradl, J.; Benner, A.; Zirbel, R.M.; Cremer, T.; Lichter, P. Active and inactive genes localize preferentially in the periphery of chromosome territories. J. Cell Biol. 1996, 135, 1195–1205. [Google Scholar] [CrossRef]
- Dietzel, S.; Schiebel, K.; Little, G.; Edelmann, P.; Rappold, G.A.; Eils, R.; Cremer, C.; Cremer, T. The 3D Positioning of ANT2 and ANT3 Genes within Female X Chromosome Territories Correlates with Gene Activity. Exp. Cell Res. 1999, 252, 363–375. [Google Scholar] [CrossRef]
- Scheuermann, M.O.; Tajbakhsh, J.; Kurz, A.; Saracoglu, K.; Eils, R.; Lichter, P. Topology of genes and nontranscribed sequences in human interphase nuclei. Exp. Cell Res. 2004, 301, 266–279. [Google Scholar] [CrossRef] [PubMed]
- Galiovà, G.; Bàrtovà, E.; Kozubek, S. Nuclear topography of beta-like globin gene cluster in IL-3-stimulated human leukemic K-562 cells. Blood Cells Mol. Dis. 2004, 33, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, J.; Luz, H.; Bornfleth, H.; Lampel, S.; Cremer, C.; Lichter, P. Spatial Distribution of GC- and AT-Rich DNA Sequences within Human Chromosome Territories. Exp. Cell Res. 2000, 255, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Torabi, K.; Wangsa, D.; Ponsa, I.; Brown, M.; Bosch, A.; Vila-Casadesùs, M.; Karpova, T.S.; Calvo, M.; Castells, A.; Miró, R.; et al. Transcription-dependent radial distribution of TCF7L2 regulated genes in chromosome territories. Chromosoma 2017, 126, 655–667. [Google Scholar] [CrossRef]
- Peng, A.Y.T.; Kolhe, J.A.; Behrens, L.D.; Freeman, B.C. Genome organization: Tag it, move it, place it. Curr. Op. Cell Biol. 2021, 68, 90–97. [Google Scholar] [CrossRef]
- Gandhi, M.S.; Stringer, J.R.; Nikiforova, M.N.; Medvedovic, M.; Nikiforov, Y.E. Gene position within chromosome territories correlates with their involvement in distinct rearrangement types in thyroid cancer cells. Genes Chromosomes Cancer 2009, 48, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Verschure, P.J.; van der Kraan, I.; Enserink, J.M.; Moné, M.J.; Manders, E.M.M.; van Driel, R. Large-scale Chromatin Organization and the Localization of Proteins Involved in Gene Expression in Human Cells. J. Histochem. Cytochem. 2002, 50, 1303–1312. [Google Scholar] [CrossRef] [Green Version]
- Branco, M.R.; Pombo, A. Intermingling of Chromosome Territories in Interphase Suggests Role in Translocations and Transcription-Dependent Associations. PLoS Biol. 2006, 4, e138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpi, E.V.; Chevret, E.; Jones, T.; Vatcheva, R.; Williamson, J.; Beck, S.; Campbell, R.D.; Goldsworthy, M.; Powis, S.H.; Ragoussis, J.; et al. Large-scale chromatin organization of the major histocompatibility complex and other regions of human chromosome 6 and its response to interferon in interphase nuclei. J. Cell Sci. 2000, 113, 1565–1576. [Google Scholar] [CrossRef]
- Williams, R.R.E.; Broad, S.; Sheer, D.; Ragoussis, J. Subchromosomal Positioning of the Epidermal Differentiation Complex (EDC) in Keratinocyte and Lymphoblast Interphase Nuclei. Exp. Cell Res. 2002, 272, 163–175. [Google Scholar] [CrossRef]
- Khanna, N.; Hu, Y.; Belmont, A.S. HSP70 Transgene Directed Motion to Nuclear Speckles Facilitates Heat Shock Activation. Curr. Biol. 2014, 24, 1138–1144. [Google Scholar] [CrossRef] [Green Version]
- Lahbib-Mansais, Y.; Barasc, H.; Marti-Marimon, M.; Mompart, F.; Iannuccelli, E.; Robelin, D.; Riquet, J.; Yerle-Bouissou, M. Expressed alleles of imprinted IGF2, DLK1 and MEG3 colocalize in 3Dpreserved nuclei of porcine fetal cells. BMC Cell Biol. 2016, 17, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, S.P.; Dutta, A. DNA Replication in Eukaryotic Cells. Annu. Rev. Biochem. 2002, 71, 333–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Mulia, J.C.; Gilbert, D.M. Replication timing and transcriptional control: Beyond cause and effect—Part III. Curr Opin. Cell Biol. 2016, 40, 168–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagin, V.O.; Casas-Delucchi, C.S.; Reinhart, M.; Schermelleh, L.; Markaki, Y.; Maiser, A.; Bolius, J.J.; Bensimon, A.; Fillies, M.; Domaing, P.; et al. 4D Visualization of replication foci in mammalian cells corresponding to individual replicons. Nat. Commun. 2016, 7, 11231. [Google Scholar] [CrossRef] [PubMed]
- Meister, P.; Taddei, A.; Gasser, S.M. In and out of the Replication Factory. Cell 2006, 125, 1233–1235. [Google Scholar] [CrossRef] [Green Version]
- Ryba, T.; Hiratani, I.; Lu, J.; Itoh, M.; Kulik, M.; Zhang, J.; Schulz, T.C.; Robins, A.J.; Dalton, S.; Gilbert, D.M. Evolutionarily conserved replication timing profiles predict long-range chromatin interactions and distinguish closely related cell types. Genome Res. 2010, 20, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Zink, D. The temporal program of DNA replication: New insights into old questions. Chromosoma 2006, 115, 273–287. [Google Scholar] [CrossRef]
- Drouin, R.; Lemieux, N.; Richer, C.-L. Chromosome condensation from prophase to late metaphase: Relationship to chromosome bands and their replication time. Cytogenet. Genome Res. 1991, 57, 91–99. [Google Scholar] [CrossRef]
- Rhind, N.; Gilbert, D.M. DNA Replication Timing. Cold Spring Harb. Perspect. Biol. 2013, 5, a010132. [Google Scholar] [CrossRef] [Green Version]
- Woodfine, K.; Fiegler, H.; Beare, D.M.; Collins, J.E.; McCann, O.T.; Young, B.D.; Debernardi, S.; Mott, R.; Dunham, I.; Carter, N.P. Replication timing of the human genome. Hum. Mol. Genet. 2004, 13, 191–202. [Google Scholar] [CrossRef]
- Dutrillaux, B.; Couturier, J.; Richer, C.-L.; Viegas-Pequinot, E. Sequence of DNA replication in 277 R- and Q-bands of human chromosomes using a BrdU treatment. Chromosoma 1976, 58, 51–61. [Google Scholar] [CrossRef]
- Federico, C.; Saccone, S.; Bernardi, G. The gene-richest bands of human chromosomes replicate at the onset of the S-phase. Cytogenet. Genome Res. 1998, 80, 83–88. [Google Scholar]
- Grasser, F.; Neusser, M.; Fiegler, H.; Thormeyer, T.; Cremer, M.; Carter, N.P.; Cremer, T.; Müller, S. Replication-timing-correlated spatial chromatin arrangements in cancer and in primate interphase nuclei. J. Cell Sci. 2008, 121, 1876–1886. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.; Paolella, G.; Ramos, C.; Lamond, A.L. Spatial organization of large-scale chromatin domains in the nucleus: A magnified view of single chromosome territories. Cell Biol. 1997, 139, 1597–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadoni, N.; Langer, S.; Fauth, C.; Bernardi, G.; Cremer, T.; Turner, B.M.; Zink, D. Nuclear organization of mammalian genomes: Polar chromosome territories build up functionally distinct higher order compartments. J. Cell Biol. 1999, 146, 1211–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donley, N.; Thayer, M.J. DNA replication timing, genome stability and cancer. Semin. Cancer Biol. 2013, 23, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Boggs, B.A.; Chinault, A.C. Analysis of replication timing properties of human Xchromosomal loci by fluorescence in situ hybridization. Proc. Natl. Acad. Sci. USA 1994, 91, 6083–6087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chess, A.; Simon, I.; Cedar, H.; Axel, R. Allelic inactivation regulates olfactory receptor gene expression. Cell 1994, 78, 823–834. [Google Scholar] [CrossRef]
- Knoll, J.H.M.; Cheng, S.-D.; Lalande, M. Allele specificity of DNA replication timing in the Angelman/Prader-Willi syndrome imprinted chromosomal region. Nat. Genet. 1994, 6, 41–46. [Google Scholar] [CrossRef]
- Simon, I.; Tenzen, T.; Reubinoff, B.E.; Hillman, D.; McCarrey, J.R.; Cedar, H. Asynchronous replication of imprinted genes is established in the gametes and maintained during development. Nature 1999, 401, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Reish, O.; Orlovski, A.; Mashevitz, M.; Sher, C.; Libman, V.; Rosenblat, M.; Avivi, L. Modified allelic replication in lymphocytes of patients with neurofibromatosis type 1. Cancer Genet. Cytogenet. 2003, 143, 133–139. [Google Scholar] [CrossRef]
- Laish, I.; Mannasse-Green, B.; Hadary, R.; Konikoff, F.M.; Amiel, A.; Kitay-Cohen, Y. Aneuploidy and asynchronous replication in non-alcholic fatty liver disease and cryptogenic cirrhosis. Gene 2016, 593, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Amiel, A.; Elis, A.; Blumenthal, D.; Gaber, E.; Fejgin, M.D.; Dubinsky, R.; Lishner, M. Modified order of allelic replication in lymphoma patients at different disease stages. Cancer Genet. Cytogenet. 2001, 125, 156–160. [Google Scholar] [CrossRef]
- Korenstein-Ilan, A.; Amiel, A.; Lalezari, S.; Lishner, M.; Avivi, L. Allele-specific replication associated with aneuploidy in blood cells of patients with hematologic malignancies. Cancer Genet. Cytogenet. 2002, 139, 97–103. [Google Scholar] [CrossRef]
- Dotan, Z.A.; Dotan, A.; Ramon, J.; Avivi, L. Aberrant allele-specific replication, independent of parental origin, in blood cells of cancer patients. BMC Cancer 2008, 8, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinberg-Rashi, H.; Cytron, S.; Gelman-Kohan, Z.; Litmanovitch, T.; Avivi, L. Replication timing aberrations and aneuploidy in peripheral blood lymphocytes of breast cancer patients. Neoplasia 2010, 12, 668–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagler, R.H.; Gray, S.W.; Romantan, A.; Kelly, B.J.; DeMichele, A.; Armstrong, K.; Schwartz, J.S.; Hornik, R.C. Differences in information seeking among breast, prostate, and colorectal cancer patients: Results from a population-based survey. Patient Educ. Couns. 2010, 81, S54–S62. [Google Scholar] [CrossRef] [Green Version]
- Ryba, T.; Battaglia, D.; Chang, B.H.; Shirley, J.W.; Buckley, Q.; Pope, B.D.; Devidas, M.; Druker, B.J.; Gilbert, D.M. Abnormal developmental control of replication-timing domains in pediatric acute lymphoblastic leukemia. Genome Res. 2012, 22, 1833–1844. [Google Scholar] [CrossRef] [Green Version]
- Fritz, A.; Sinha, S.; Marella, N.; Berezney, R. Alterations in replication timing of cancer related genes in malignant human breast cancer cells. J. Cell Biochem. 2013, 114, 1074–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Han, M.; Qi, L.S. Engineering 3D genome organization. Nat. Rev. Genet. 2021, 22, 343–360. [Google Scholar] [CrossRef]
- Harewood, L.; Fraser, P. The impact of chromosomal rearrangements on regulation of gene expression. Hum. Mol. Genet. 2014, 23, R76–R82. [Google Scholar] [CrossRef] [Green Version]
- Gheldof, N.; Witwicki, R.; Migliavacca, E.; Leleu, M.; Didelot, G.; Harewood, L.; Rougemont, J.; Reymond, A. Structural variation-associated expression changes are paralleled by chromatin architecture modifications. PLoS ONE 2013, 8, e79973. [Google Scholar]
- Ballabio, A.; Gieselmann, V. Lysosomal disorders: From storage to cellular damage. Biochim. Biophys. Acta 2009, 1793, 684–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harewood, L.; Schutz, F.; Boyle, S.; Perry, P.; Delorenzi, M.; Bickmore, W.A.; Reymond, A. The effect of translocation-induced nuclear reorganization on gene expression. Genome Res. 2010, 20, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Federico, C.; Owoka, T.; Ragusa, D.; Sturiale, V.; Caponnetto, D.; Leotta, C.G.; Bruno, F.; Foster, H.A.; Rigamonti, S.; Giudici, G.; et al. Deletions of chromosome 7q affect nuclear organization and HLXB9 gene expression in hematological disorders. Cancers 2019, 11, 585. [Google Scholar] [CrossRef] [Green Version]
- Taslerovà, R.; Kozubek, S.; Lukàsovà, E.; Jirsovà, P.; Bàrtovà, E.; Kozubek, M. Arrangement of chromosome 11 and 22 territories, EWSR1 and FLI1 genes, and other genetic elements of these chromosomes in human lymphocytes and Ewing sarcoma cells. Hum. Genet. 2003, 112, 143–155. [Google Scholar] [CrossRef]
- Lukàsovà, E.; Kozubek, S.; Kozubek, M.; Kjeronskà, J.; Ryznar, L.; Horàkovà, J.; Horneck, G. Localisation and distance between ABL and BCR genes in interphase nuclei of bone marrow cells of control donors and patients with chronic myeloid leukaemia. Hum. Genet. 1997, 100, 525–535. [Google Scholar]
- Murmann, A.E.; Gao, J.; Encinosa, M.; Gautier, M.; Peter, M.E.; Eils, R.; Lichter, P.; Rowley, J.D. Local gene density predicts the spatial position of genetic loci in the interphase nucleus. Exp. Cell Res. 2005, 311, 14–26. [Google Scholar] [CrossRef]
- Meaburn, K.J.; Misteli, T. Locus-specific and activity-independent gene repositioning during early tumorigenesis. J. Cell Biol. 2008, 180, 39–50. [Google Scholar] [CrossRef]
- Morey, C.; Da Silva, N.R.; Kmita, M.; Duboule, D.; Bickmore, W.A. Ectopic nuclear reorganization driven by a Hoxb1 transgene transposed into Hoxd. J. Cell Sci. 2008, 121, 571–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, C.; Leotta, C.G.; Bruno, F.; Longo, A.M.; Owoka, T.; Tosi, S.; Saccone, S. Nuclear repositioning of the non-translocated HLXB9 allele in the leukaemia cell line GDM-1 harbouring a t(6;7)(q23;q36). Cytogenet. Genome Res. 2017, 153, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Roix, J.J.; McQueen, P.G.; Munson, P.J.; Parada, L.A.; Misteli, T. Spatial proximity of translocation-prone gene loci in human lymphomas. Nat. Genet. 2003, 34, 287–291. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Federico, C.; Bruno, F.; Ragusa, D.; Clements, C.S.; Brancato, D.; Henry, M.P.; Bridger, J.M.; Tosi, S.; Saccone, S. Chromosomal Rearrangements and Altered Nuclear Organization: Recent Mechanistic Models in Cancer. Cancers 2021, 13, 5860. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13225860
Federico C, Bruno F, Ragusa D, Clements CS, Brancato D, Henry MP, Bridger JM, Tosi S, Saccone S. Chromosomal Rearrangements and Altered Nuclear Organization: Recent Mechanistic Models in Cancer. Cancers. 2021; 13(22):5860. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13225860
Chicago/Turabian StyleFederico, Concetta, Francesca Bruno, Denise Ragusa, Craig S. Clements, Desiree Brancato, Marianne P. Henry, Joanna M. Bridger, Sabrina Tosi, and Salvatore Saccone. 2021. "Chromosomal Rearrangements and Altered Nuclear Organization: Recent Mechanistic Models in Cancer" Cancers 13, no. 22: 5860. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13225860