
miRNAs in the Regulation of Cancer Immune Response: Effect of miRNAs on Cancer Immunotherapy

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. miRNAs—Biogenesis, Biochemistry and Functions
2.1. Oncogenic miRNAs
2.2. miRNAs as Biomarkers
3. Innate and Adaptive Immunity
4. Cancer Immune Escape
5. Immuno-miRNAs: Central Regulators of Immunity
6. Immune Cell Pathways Regulated by miRNAs in Cancer
6.1. Regulation of Monocytes and Macrophages by miRNA
6.2. Regulation of Natural Killer Cells (NK Cells) by miRNA
6.3. Regulation of T Helper Cells and Cytotoxic T Cells by miRNA
6.4. Regulation of Immune-Checkpoint Molecules by miRNA
7. Effect of miRNAs on Cancer Immunotherapy
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thun, M.J.; DeLancey, J.O.; Center, M.M.; Jemal, A.; Ward, E.M. The Global Burden of Cancer: Priorities for Prevention. Carcinogenesis 2010, 31, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Hassanpour, S.H.; Dehghani, M. Review of Cancer from Perspective of Molecular. J. Cancer Res. Pract. 2017, 4, 127–129. [Google Scholar] [CrossRef]
- American Cancer Society. Global Cancer Facts & Figures. Available online: https://www.cancer.org/research/cancer-facts-statistics/global.html (accessed on 4 February 2021).
- Fisher, R.; Pusztai, L.; Swanton, C. Cancer Heterogeneity: Implications for Targeted Therapeutics. Br. J. Cancer 2013, 108, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaffer, C.L.; Weinberg, R.A. A Perspective on Cancer Cell Metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef]
- Alhmied, F.I.; Alammar, A.H.; Alsultan, B.M.; Alshehri, M.; Pottoo, F.H. Molecular Mechanisms of Thymoquinone as Anticancer Agent. Comb. Chem. High. Throughput Screen 2020. [Google Scholar] [CrossRef]
- Poon, S.L.; McPherson, J.R.; Tan, P.; Teh, B.T.; Rozen, S.G. Mutation Signatures of Carcinogen Exposure: Genome-Wide Detection and New Opportunities for Cancer Prevention. Genome Med. 2014, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Trafialek, J.; Kolanowski, W. Dietary Exposure to Meat-Related Carcinogenic Substances: Is There a Way to Estimate the Risk? Int. J. Food Sci. Nutr. 2014, 65, 774–780. [Google Scholar] [CrossRef]
- Cumberbatch, M.G.K.; Cox, A.; Teare, D.; Catto, J.W.F. Contemporary Occupational Carcinogen Exposure and Bladder Cancer: A Systematic Review and Meta-Analysis. JAMA Oncol. 2015, 1, 1282–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antwi, S.O.; Eckert, E.C.; Sabaque, C.V.; Leof, E.R.; Hawthorne, K.M.; Bamlet, W.R.; Chaffee, K.G.; Oberg, A.L.; Petersen, G.M. Exposure to Environmental Chemicals and Heavy Metals, and Risk of Pancreatic Cancer. Cancer Causes Control. 2015, 26, 1583–1591. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Lu, H.; Wang, X.; Jin, H. MicroRNAs as Potential Biomarkers in Cancer: Opportunities and Challenges. Available online: https://www.hindawi.com/journals/bmri/2015/125094/ (accessed on 4 February 2021).
- Aboelmagd, A.; Rayes, S.M.E.; Gomaa, M.S.; Ali, I.A.I.; Fathalla, W.; Pottoo, F.H.; Khan, F.A.; Khalifa, M.E. The Synthesis and Antiproliferative Activity of New N-Allyl Quinoxalinecarboxamides and Their O-Regioisomers. New J. Chem. 2021, 45, 831–849. [Google Scholar] [CrossRef]
- Aboelmagd, A.; Alotaibi, S.H.; Rayes, S.M.E.; Elsayed, G.M.; Ali, I.A.I.; Fathalla, W.; Pottoo, F.H.; Khan, F.A. Synthesis and Anti Proliferative Activity of New N-Pentylquinoxaline Carboxamides and Their O-Regioisomer. ChemistrySelect 2020, 5, 13439–13453. [Google Scholar] [CrossRef]
- Nawaz, F.; Alam, O.; Perwez, A.; Rizvi, M.A.; Naim, M.J.; Siddiqui, N.; Pottoo, F.H.; Jha, M. 3′-(4-(Benzyloxy)Phenyl)-1′-Phenyl-5-(Heteroaryl/Aryl)-3,4-Dihydro-1′H,2H-[3,4′-Bipyrazole]-2-Carboxamides as EGFR Kinase Inhibitors: Synthesis, Anticancer Evaluation, and Molecular Docking Studies. Arch. Der Pharm. 2020, 353, 1900262. [Google Scholar] [CrossRef] [PubMed]
- Rayes, S.M.E.; Aboelmagd, A.; Gomaa, M.S.; Fathalla, W.; Ali, I.A.; Pottoo, F.H.; Khan, F.A. Newly Synthesized 3-(4-Chloro-Phenyl)-3-Hydroxy-2,2-Dimethyl-Propionic Acid Methyl Ester Derivatives Selectively Inhibit the Proliferation of Colon Cancer Cells. RSC Adv. 2020, 10, 8825–8841. [Google Scholar] [CrossRef]
- Barger, J.F.; Rahman, M.A.; Jackson, D.; Acunzo, M.; Nana-Sinkam, S.P. Extracellular MiRNAs as Biomarkers in Cancer. Food Chem. Toxicol. 2016, 98, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohel, M.M.H. Circulating MicroRNAs as Biomarkers in Cancer Diagnosis. Life Sci. 2020, 248, 117473. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.-L.T.; Schmittgen, T.D.; et al. Detection of MicroRNA Expression in Human Peripheral Blood Microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottoo, F.H.; Javed, M.N.; Rahman, J.U.; Abu-Izneid, T.; Khan, F.A. Targeted Delivery of MiRNA Based Therapeuticals in the Clinical Management of Glioblastoma Multiforme. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Pottoo, F.H.; Barkat, M.A.; Ansari, M.A.; Javed, M.N.; Jamal, Q.M.S.; Kamal, M.A. Nanotechnological Based MiRNA Intervention in the Therapeutic Management of Neuroblastoma. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Sohel, M.H. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. MicroRNAs in Cancer Management. Lancet Oncol. 2012, 13, e249–e258. [Google Scholar] [CrossRef]
- Shafi, S.; Khan, S.; Hoda, F.; Fayaz, F.; Singh, A.; Khan, M.A.; Ali, R.; Pottoo, F.H.; Tariq, S.; Najmi, A.K. Decoding Novel Mechanisms and Emerging Therapeutic Strategies in Breast Cancer Resistance. Curr. Drug Metab. 2020, 21, 199–210. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The Role of MicroRNAs in Human Cancer. Signal Transduct. Target. Ther. 2016, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA Polymerase III Transcribes Human MicroRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA Genes Are Transcribed by RNA Polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Denli, A.M.; Tops, B.B.J.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of Primary MicroRNAs by the Microprocessor Complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-Methyladenosine Marks Primary MicroRNAs for Processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 Complexes Carry a Population of Circulating MicroRNAs Independent of Vesicles in Human Plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Li, M.; Vilborg, A.; Lee, N.; Shu, M.-D.; Yartseva, V.; Šestan, N.; Steitz, J.A. Mammalian 5’-Capped MicroRNA Precursors That Generate a Single MicroRNA. Cell 2013, 155, 1568–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prud’homme, G.J.; Glinka, Y.; Lichner, Z.; Yousef, G.M. Neuropilin-1 Is a Receptor for Extracellular MiRNA and AGO2/MiRNA Complexes and Mediates the Internalization of MiRNAs That Modulate Cell Function. Oncotarget 2016, 7, 68057–68071. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-S.; Maurin, T.; Robine, N.; Rasmussen, K.D.; Jeffrey, K.L.; Chandwani, R.; Papapetrou, E.P.; Sadelain, M.; O’Carroll, D.; Lai, E.C. Conserved Vertebrate Mir-451 Provides a Platform for Dicer-Independent, Ago2-Mediated MicroRNA Biogenesis. Proc. Natl. Acad. Sci. USA 2010. [Google Scholar] [CrossRef] [Green Version]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic MicroRNA Precursors That Bypass Drosha Processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandellini, P.; Giovannetti, E.; Nicassio, F. MicroRNAs in Cancer Management: Big Challenges for Small Molecules. Biomed. Res. Int. 2015, 2015, 982156. [Google Scholar] [CrossRef] [PubMed]
- Otmani, K.; Lewalle, P. Tumor Suppressor MiRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar] [CrossRef] [PubMed]
- Coronnello, C.; Benos, P.V. ComiR: Combinatorial MicroRNA Target Prediction Tool. Nucleic Acids Res. 2013, 41, W159–W164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, Y.; Karsenty, S.; Linial, M. MiRror-Suite: Decoding Coordinated Regulation by MicroRNAs. Database 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths-Jones, S. The MicroRNA Registry. Nucleic Acids Res. 2004, 32, D109–D111. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J.P.J.; Keown, A.A.; Chong, M.M.W. The MiR-17∼92a Cluster of MicroRNAs Is Required for the Fitness of Foxp3+ Regulatory T Cells. PLoS ONE 2014, 9, e88997. [Google Scholar] [CrossRef]
- Kohlhapp, F.; Mitra, A.K.; Lengyel, E.; Peter, M.E. MicroRNAs as mediators and communicators between cancer cells and the tumor microenvironment. Oncogene 2015, 34, 5857–5868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human MicroRNA Genes Are Frequently Located at Fragile Sites and Genomic Regions Involved in Cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, K.B. MicroRNA (MiRNA) in Cancer. Cancer Cell Int. 2015, 15, 38. [Google Scholar] [CrossRef] [Green Version]
- Izumchenko, E.; Chang, X.; Michailidi, C.; Kagohara, L.; Ravi, R.; Paz, K.; Brait, M.; Hoque, M.O.; Ling, S.; Bedi, A.; et al. The TGFβ-MiR200-MIG6 Pathway Orchestrates the EMT-Associated Kinase Switch That Induces Resistance to EGFR Inhibitors. Cancer Res. 2014, 74, 3995–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiocca, E.A.; Lawler, S.E. The Many Functions of MicroRNAs in Glioblastoma. World Neurosurg. 2010, 73, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Godlewski, J.; Bronisz, A.; Nowicki, M.O.; Chiocca, E.A.; Lawler, S. MicroRNA-451: A Conditional Switch Controlling Glioma Cell Proliferation and Migration. Cell Cycle 2010, 9, 2742–2748. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in Cancer: Biomarkers, Functions and Therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Wiemer, E.A.C. The Role of MicroRNAs in Cancer: No Small Matter. Eur. J. Cancer 2007, 43, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Mehrgou, A.; Akouchekian, M. Therapeutic Impacts of MicroRNAs in Breast Cancer by Their Roles in Regulating Processes Involved in This Disease. J. Res. Med. Sci. 2017, 22, 130. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Yu, Z.; Duan, Z.; Kan, Q. Role of MicroRNA-1 in Human Cancer and Its Therapeutic Potentials. Biomed. Res. Int. 2014, 2014, 428371. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Jeon, K.; Lee, J.-T.; Kim, S.; Kim, V.N. MicroRNA Maturation: Stepwise Processing and Subcellular Localization. EMBO J. 2002, 21, 4663–4670. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Frixa, T.; Donzelli, S.; Blandino, G. Oncogenic MicroRNAs: Key Players in Malignant Transformation. Cancers 2015, 7, 2466–2485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. MicroRNAs as Oncogenes and Tumor Suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 Family: A Potential Tumor Suppressor and Therapeutic Candidate in Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wu, L.; Sun, Y.; Yin, Q.; Chen, X.; Liang, S.; Meng, Q.; Long, H.; Li, F.; Luo, C.; et al. Mir-421 in Plasma as a Potential Diagnostic Biomarker for Precancerous Gastric Lesions and Early Gastric Cancer. PeerJ 2019, 7, e7002. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-H.; Lee, C.; Chiu, C.-T.; Chu, Y.-Y.; Cheng, H.-T.; Hsu, J.-T.; Tsou, Y.-K.; Wu, R.-C.; Chen, T.-C.; Chang, N.-C.; et al. Circulating MicroRNA-196a Is an Early Gastric Cancer Biomarker. Oncotarget 2018, 9, 10317–10323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desantis, V.; Solimando, A.G.; Saltarella, I.; Sacco, A.; Giustini, V.; Bento, M.; Lamanuzzi, A.; Melaccio, A.; Frassanito, M.A.; Paradiso, A.; et al. MicroRNAs as a Potential New Preventive Approach in the Transition from Asymptomatic to Symptomatic Multiple Myeloma Disease. Cancers 2021, 13, 3650. [Google Scholar] [CrossRef]
- Marcucci, G.; Mrózek, K.; Radmacher, M.D.; Garzon, R.; Bloomfield, C.D. The Prognostic and Functional Role of MicroRNAs in Acute Myeloid Leukemia. Blood 2011, 117, 1121–1129. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, L.; Dong, D.; Wang, Z.; Ji, W.; Yu, M.; Zhang, F.; Niu, R.; Zhou, Y. MiR-34b/c-5p and the Neurokinin-1 Receptor Regulate Breast Cancer Cell Proliferation and Apoptosis. Cell Prolif. 2019, 52, e12527. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; Gao, J.; Zhang, T.; Li, S.; Luo, A.; Chen, H.; Ding, F.; Wang, X.; Liu, Z. MicroRNA-34 Suppresses Breast Cancer Invasion and Metastasis by Directly Targeting Fra-1. Oncogene 2013, 32, 4294–4303. [Google Scholar] [CrossRef]
- Acunzo, M.; Romano, G.; Palmieri, D.; Laganá, A.; Garofalo, M.; Balatti, V.; Drusco, A.; Chiariello, M.; Nana-Sinkam, P.; Croce, C.M. Cross-Talk between MET and EGFR in Non-Small Cell Lung Cancer Involves MiR-27a and Sprouty2. Proc. Natl. Acad. Sci. USA 2013, 110, 8573–8578. [Google Scholar] [CrossRef] [Green Version]
- Outeiro-Pinho, G.; Barros-Silva, D.; Aznar, E.; Sousa, A.-I.; Vieira-Coimbra, M.; Oliveira, J.; Gonçalves, C.S.; Costa, B.M.; Junker, K.; Henrique, R.; et al. MicroRNA-30a-5pme: A Novel Diagnostic and Prognostic Biomarker for Clear Cell Renal Cell Carcinoma in Tissue and Urine Samples. J. Exp. Clin. Cancer Res. 2020, 39, 98. [Google Scholar] [CrossRef]
- Song, S.; Long, M.; Yu, G.; Cheng, Y.; Yang, Q.; Liu, J.; Wang, Y.; Sheng, J.; Wang, L.; Wang, Z.; et al. Urinary Exosome MiR-30c-5p as a Biomarker of Clear Cell Renal Cell Carcinoma That Inhibits Progression by Targeting HSPA5. J. Cell. Mol. Med. 2019, 23, 6755–6765. [Google Scholar] [CrossRef]
- Li, Y.-J.; Du, L.; Aldana-Masangkay, G.; Wang, X.; Urak, R.; Forman, S.J.; Rosen, S.T.; Chen, Y. Regulation of MiR-34b/c-Targeted Gene Expression Program by SUMOylation. Nucleic Acids Res. 2018, 46, 7108–7123. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Levi, E.; Majumdar, A.P.; Sarkar, F.H. Expression of MiR-34 Is Lost in Colon Cancer Which Can Be Re-Expressed by a Novel Agent CDF. J. Hematol. Oncol. 2012, 5, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Hermeking, H. MiR-34a and MiR-34b/c Suppress Intestinal Tumorigenesis. Cancer Res. 2017, 77, 2746–2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Li, Y.; Daniels, G.; Sfanos, K.; De Marzo, A.; Wei, J.; Li, X.; Chen, W.; Wang, J.; Zhong, X.; et al. LEF1 Targeting EMT in Prostate Cancer Invasion Is Regulated by MiR-34a. Mol. Cancer Res. 2015, 13, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.-L.; Sun, B.-F.; Huang, L.-R.; Yuan, H.-B.; Zhang, S.; Chen, J.; Yu, Z.-J.; Luo, H. Potent Inhibition of MiR-34b on Migration and Invasion in Metastatic Prostate Cancer Cells by Regulating the TGF-β Pathway. Int. J. Mol. Sci. 2017, 18, E2762. [Google Scholar] [CrossRef] [Green Version]
- Stahlhut, C.; Slack, F.J. Combinatorial Action of MicroRNAs Let-7 and MiR-34 Effectively Synergizes with Erlotinib to Suppress Non-Small Cell Lung Cancer Cell Proliferation. Cell Cycle 2015, 14, 2171–2180. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Cheng, J.; Chen, B.; Liu, Q.; Xu, D.; Zhang, Y. Circulating MicroRNA-34 Family Low Expression Correlates with Poor Prognosis in Patients with Non-Small Cell Lung Cancer. J. Thorac. Dis. 2017, 9, 3735–3746. [Google Scholar] [CrossRef] [Green Version]
- Kasinski, A.L.; Slack, F.J. MiRNA-34 Prevents Cancer Initiation and Progression in a Therapeutically Resistant K-Ras and P53-Induced Mouse Model of Lung Adenocarcinoma. Cancer Res. 2012, 72, 5576–5587. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Liu, J.; Chen, J.; Dong, J.; Ma, H.; Liu, Y.; Hu, Z. Methylation-Associated Silencing of MicroRNA-34b in Hepatocellular Carcinoma Cancer. Gene 2014, 543, 101–107. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, L.-S.; Yuan, W.; Wu, Z.; Wang, H.-B.; Xu, T.; Sun, J.-C.; Cheng, K.-F.; Shi, J.-G. Low MiR-34a and MiR-192 Are Associated with Unfavorable Prognosis in Patients Suffering from Osteosarcoma. Am. J. Transl. Res. 2015, 7, 111–119. [Google Scholar]
- Gang, L.; Qun, L.; Liu, W.-D.; Li, Y.-S.; Xu, Y.-Z.; Yuan, D.-T. MicroRNA-34a Promotes Cell Cycle Arrest and Apoptosis and Suppresses Cell Adhesion by Targeting DUSP1 in Osteosarcoma. Am. J. Transl. Res. 2017, 9, 5388–5399. [Google Scholar]
- Xi, L.; Zhang, Y.; Kong, S.; Liang, W. MiR-34 Inhibits Growth and Promotes Apoptosis of Osteosarcoma in Nude Mice through Targetly Regulating TGIF2 Expression. Biosci. Rep. 2018, 38, BSR20180078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Ren, W.; Chen, K. MiR-34a Promotes Apoptosis and Inhibits Autophagy by Targeting HMGB1 in Acute Myeloid Leukemia Cells. Cell. Physiol. Biochem. 2017, 41, 1981–1992. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Wang, H.; Li, L.; Ma, X.; Chen, Y.; Zhou, H.; Luo, Y.; Xiao, Y.; Liu, L. MiR-34c-5p Promotes Eradication of Acute Myeloid Leukemia Stem Cells by Inducing Senescence through Selective RAB27B Targeting to Inhibit Exosome Shedding. Leukemia 2018, 32, 1180–1188. [Google Scholar] [CrossRef]
- Wong, K.Y.; Yim, R.L.H.; So, C.C.; Jin, D.-Y.; Liang, R.; Chim, C.S. Epigenetic Inactivation of the MIR34B/C in Multiple Myeloma. Blood 2011, 118, 5901–5904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent Deletions and Down-Regulation of Micro-RNA Genes MiR15 and MiR16 at 13q14 in Chronic Lymphocytic Leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roderburg, C.; Luedde, T. Circulating MicroRNAs as Markers of Liver Inflammation, Fibrosis and Cancer. J. Hepatol. 2014, 61, 1434–1437. [Google Scholar] [CrossRef] [Green Version]
- Sohn, W.; Kim, J.; Kang, S.H.; Yang, S.R.; Cho, J.-Y.; Cho, H.C.; Shim, S.G.; Paik, Y.-H. Serum Exosomal MicroRNAs as Novel Biomarkers for Hepatocellular Carcinoma. Exp. Mol. Med. 2015, 47, e184. [Google Scholar] [CrossRef]
- De Silva, S.; Tennekoon, K.H.; Karunanayake, E.H. Interaction of Gut Microbiome and Host MicroRNAs with the Occurrence of Colorectal and Breast Cancer and Their Impact on Patient Immunity. Oncotarget 2021, 14, 5115–5129. [Google Scholar] [CrossRef]
- Iftikhar, H.; Carney, G.E. Evidence and Potential in Vivo Functions for Biofluid MiRNAs: From Expression Profiling to Functional Testing. BioEssays 2016, 38, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of MicroRNAs in Living Cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Hannafon, B.N.; Carpenter, K.J.; Berry, W.L.; Janknecht, R.; Dooley, W.C.; Ding, W.-Q. Exosome-Mediated MicroRNA Signaling from Breast Cancer Cells Is Altered by the Anti-Angiogenesis Agent Docosahexaenoic Acid (DHA). Mol. Cancer 2015, 14, 133. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Chen, J.; Su, F.; Yu, B.; Su, F.; Lin, L.; Liu, Y.; Huang, J.-D.; Song, E. Microvesicles Secreted by Macrophages Shuttle Invasion-Potentiating MicroRNAs into Breast Cancer Cells. Mol. Cancer 2011, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Sakha, S.; Muramatsu, T.; Ueda, K.; Inazawa, J. Exosomal MicroRNA MiR-1246 Induces Cell Motility and Invasion through the Regulation of DENND2D in Oral Squamous Cell Carcinoma. Sci. Rep. 2016, 6, 38750. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Rao, S.-S.; Wang, Z.-X.; Cao, J.; Tan, Y.-J.; Luo, J.; Li, H.-M.; Zhang, W.-S.; Chen, C.-Y.; Xie, H. Exosomes from Human Umbilical Cord Blood Accelerate Cutaneous Wound Healing through MiR-21-3p-Mediated Promotion of Angiogenesis and Fibroblast Function. Theranostics 2018, 8, 169–184. [Google Scholar] [CrossRef]
- Zhou, W.; Fong, M.Y.; Min, Y.; Somlo, G.; Liu, L.; Palomares, M.R.; Yu, Y.; Chow, A.; O’Connor, S.T.F.; Chin, A.R.; et al. Cancer-Secreted MiR-105 Destroys Vascular Endothelial Barriers to Promote Metastasis. Cancer Cell 2014, 25, 501–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.; Chen, J. MicroRNAs in Cancer Biology and Therapy: Current Status and Perspectives. Genes Dis. 2014, 1, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Wang, C.; Chen, X.; Chen, S.; Zhang, Y.; Zhi, F.; Wang, J.; Li, L.; Zhou, X.; Li, N.; et al. Identification of Seven Serum MicroRNAs from a Genome-Wide Serum MicroRNA Expression Profile as Potential Noninvasive Biomarkers for Malignant Astrocytomas. Int. J. Cancer 2013, 132, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Arab, A.; Karimipoor, M.; Irani, S.; Kiani, A.; Zeinali, S.; Tafsiri, E.; Sheikhy, K. Potential Circulating MiRNA Signature for Early Detection of NSCLC. Cancer Genet. 2017, 216–217, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating MicroRNAs as Potential Cancer Biomarkers: The Advantage and Disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Mao, F.; Shen, T.; Luo, Q.; Ding, Z.; Qian, L.; Huang, J. Plasma MiR-145, MiR-20a, MiR-21 and MiR-223 as Novel Biomarkers for Screening Early-Stage Non-Small Cell Lung Cancer. Oncol. Lett. 2017, 13, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, S.M.; Krüger, C.; Park, B.; Derkow, K.; Rosenberger, K.; Baumgart, J.; Trimbuch, T.; Eom, G.; Hinz, M.; Kaul, D.; et al. An Unconventional Role for MiRNA: Let-7 Activates Toll-like Receptor 7 and Causes Neurodegeneration. Nat. Neurosci. 2012, 15, 827–835. [Google Scholar] [CrossRef]
- Fabbri, M. MicroRNAs and MiRceptors: A New Mechanism of Action for Intercellular Communication. Philos. Trans. R Soc. Lond. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef]
- Khan, K.; Javed, Z.; Sadia, H.; Sharifi-Rad, J.; Cho, W.C.; Luparello, C. Quercetin and MicroRNA Interplay in Apoptosis Regulation in Ovarian Cancer. Curr. Pharm. Des. 2021, 27, 2328–2336. [Google Scholar] [CrossRef]
- Wang, F.; Zheng, Z.; Guo, J.; Ding, X. Correlation and Quantitation of MicroRNA Aberrant Expression in Tissues and Sera from Patients with Breast Tumor. Gynecol. Oncol. 2010, 119, 586–593. [Google Scholar] [CrossRef]
- Bhat, S.A.; Majid, S.; Hassan, T. MicroRNAs and Its Emerging Role as Breast Cancer Diagnostic Marker—A Review. Adv. Biomark. Sci. Technol. 2019, 1, 1–8. [Google Scholar] [CrossRef]
- Asaga, S.; Kuo, C.; Nguyen, T.; Terpenning, M.; Giuliano, A.E.; Hoon, D.S.B. Direct Serum Assay for MicroRNA-21 Concentrations in Early and Advanced Breast Cancer. Clin. Chem. 2011, 57, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelli, V.; Compagnoni, C.; Capelli, R.; Cannita, K.; Sidoni, T.; Ficorella, C.; Capalbo, C.; Zazzeroni, F.; Tessitore, A.; Alesse, E. Circulating MicroRNAs as Prognostic and Therapeutic Biomarkers in Breast Cancer Molecular Subtypes. J. Pers. Med. 2020, 10, E98. [Google Scholar] [CrossRef] [PubMed]
- Khalighfard, S.; Alizadeh, A.M.; Irani, S.; Omranipour, R. Plasma MiR-21, MiR-155, MiR-10b, and Let-7a as the Potential Biomarkers for the Monitoring of Breast Cancer Patients. Sci. Rep. 2018, 8, 17981. [Google Scholar] [CrossRef]
- Jones, C.I.; Zabolotskaya, M.V.; King, A.J.; Stewart, H.J.S.; Horne, G.A.; Chevassut, T.J.; Newbury, S.F. Identification of Circulating MicroRNAs as Diagnostic Biomarkers for Use in Multiple Myeloma. Br. J. Cancer 2012, 107, 1987–1996. [Google Scholar] [CrossRef]
- Xu, P.; Xia, T.; Ling, Y.; Chen, B. MiRNAs with Prognostic Significance in Multiple Myeloma: A Systemic Review and Meta-Analysis. Medicine 2019, 98, e16711. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. MiRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplin, D.D. Overview of the Immune Response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef] [PubMed]
- Grizzi, F.; Borroni, E.M.; Yiu, D.; Farina, F.M.; Cananzi, F.C.M.; Laghi, L. Prognostic Value of Innate and Adaptive Immunity in Cancers. In Cancer Immunology: A Translational Medicine Context; Rezaei, N., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 403–415. ISBN 978-3-030-30845-2. [Google Scholar]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The Microbiome and Innate Immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.B.; Beatty, G.L. The Interplay between Innate and Adaptive Immunity in Cancer Shapes the Productivity of Cancer Immunosurveillance. J. Leukoc. Biol. 2020, 108, 363–376. [Google Scholar] [CrossRef]
- Bonilla, F.A.; Oettgen, H.C. Adaptive Immunity. J. Allergy Clin. Immunol. 2010, 125, S33–S40. [Google Scholar] [CrossRef]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or Adaptive Immunity? The Example of Natural Killer Cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, Mast Cells, Basophils, and Eosinophils. J. Allergy Clin. Immunol. 2010, 125, S73–S80. [Google Scholar] [CrossRef]
- Zhu, J.; Paul, W.E. Heterogeneity and Plasticity of T Helper Cells. Cell Res. 2010, 20, 4–12. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, D.; Sanin, D.E.; Pearce, E.J.; Pearce, E.L. Metabolic Interventions in the Immune Response to Cancer. Nat. Rev. Immunol. 2019, 19, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Zamarron, B.F.; Chen, W. Dual Roles of Immune Cells and Their Factors in Cancer Development and Progression. Int. J. Biol. Sci. 2011, 7, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The Immunobiology of Cancer Immunosurveillance and Immunoediting. Immunity 2004, 21, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.; Verhagen, J.; Blaser, K.; Akdis, M.; Akdis, C.A. Mechanisms of Immune Suppression by Interleukin-10 and Transforming Growth Factor-Beta: The Role of T Regulatory Cells. Immunology 2006, 117, 433–442. [Google Scholar] [CrossRef]
- Gasparoto, T.H.; de Souza Malaspina, T.S.; Benevides, L.; de Melo, E.J.F.; Costa, M.R.S.N.; Damante, J.H.; Ikoma, M.R.V.; Garlet, G.P.; Cavassani, K.A.; da Silva, J.S.; et al. Patients with Oral Squamous Cell Carcinoma Are Characterized by Increased Frequency of Suppressive Regulatory T Cells in the Blood and Tumor Microenvironment. Cancer Immunol. Immunother. 2010, 59, 819–828. [Google Scholar] [CrossRef]
- Marin-Acevedo, J.A.; Dholaria, B.; Soyano, A.E.; Knutson, K.L.; Chumsri, S.; Lou, Y. Next Generation of Immune Checkpoint Therapy in Cancer: New Developments and Challenges. J. Hematol. Oncol. 2018, 11, 39. [Google Scholar] [CrossRef]
- Yi, M.; Xu, L.; Jiao, Y.; Luo, S.; Li, A.; Wu, K. The Role of Cancer-Derived MicroRNAs in Cancer Immune Escape. J. Hematol. Oncol. 2020, 13, 25. [Google Scholar] [CrossRef] [Green Version]
- Restifo, N.P.; Esquivel, F.; Kawakami, Y.; Yewdell, J.W.; Mulé, J.J.; Rosenberg, S.A.; Bennink, J.R. Identification of Human Cancers Deficient in Antigen Processing. J. Exp. Med. 1993, 177, 265–272. [Google Scholar] [CrossRef]
- Mariathasan, S.; Turley, S.J.; Nickles, D.; Castiglioni, A.; Yuen, K.; Wang, Y.; Kadel Iii, E.E.; Koeppen, H.; Astarita, J.L.; Cubas, R.; et al. TGFβ Attenuates Tumour Response to PD-L1 Blockade by Contributing to Exclusion of T Cells. Nature 2018, 554, 544–548. [Google Scholar] [CrossRef]
- Lin, A.; Zhang, H.; Hu, X.; Chen, X.; Wu, G.; Luo, P.; Zhang, J. Age, Sex, and Specific Gene Mutations Affect the Effects of Immune Checkpoint Inhibitors in Colorectal Cancer. Pharmacol. Res. 2020, 159, 105028. [Google Scholar] [CrossRef]
- Hirschberger, S.; Hinske, L.C.; Kreth, S. MiRNAs: Dynamic Regulators of Immune Cell Functions in Inflammation and Cancer. Cancer Lett. 2018, 431, 11–21. [Google Scholar] [CrossRef]
- Bourquin, C.; Pommier, A.; Hotz, C. Harnessing the Immune System to Fight Cancer with Toll-like Receptor and RIG-I-like Receptor Agonists. Pharmacol. Res. 2020, 154, 104192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqubal, A.; Sharma, S.; Najmi, A.K.; Syed, M.A.; Ali, J.; Alam, M.M.; Haque, S.E. Nerolidol Ameliorates Cyclophosphamide-Induced Oxidative Stress, Neuroinflammation and Cognitive Dysfunction: Plausible Role of Nrf2 and NF- ΚB. Life Sci. 2019, 236, 116867. [Google Scholar] [CrossRef] [PubMed]
- Iqubal, A.; Sharma, S.; Ansari, M.A.; Najmi, A.K.; Syed, M.A.; Ali, J.; Alam, M.M.; Ahmad, S.; Haque, S.E. Nerolidol Attenuates Cyclophosphamide-Induced Cardiac Inflammation, Apoptosis and Fibrosis in Swiss Albino Mice. Eur. J. Pharmacol. 2019, 863, 172666. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Fukuda, Y.; Kiga, K.; Sharif, J.; Hino, K.; Enomoto, Y.; Kawamura, A.; Nakamura, K.; Takeuchi, T.; Tanabe, M. An Evolutionarily Conserved Mechanism for MicroRNA-223 Expression Revealed by MicroRNA Gene Profiling. Cell 2007, 129, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Kurkewich, J.L.; Hansen, J.; Klopfenstein, N.; Zhang, H.; Wood, C.; Boucher, A.; Hickman, J.; Muench, D.E.; Grimes, H.L.; Dahl, R. The MiR-23a~27a~24-2 MicroRNA Cluster Buffers Transcription and Signaling Pathways during Hematopoiesis. PLOS Genet. 2017, 13, e1006887. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, B.; Fu, Y.; He, M.; Wang, J.; Shen, P.; Bai, L. MiR-23a Suppresses Proliferation of Osteosarcoma Cells by Targeting SATB1. Tumour Biol. 2015, 36, 4715–4721. [Google Scholar] [CrossRef]
- Cho, S.; Wu, C.-J.; Yasuda, T.; Cruz, L.O.; Khan, A.A.; Lin, L.-L.; Nguyen, D.T.; Miller, M.; Lee, H.-M.; Kuo, M.-L.; et al. MiR-23∼27∼24 Clusters Control Effector T Cell Differentiation and Function. J. Exp. Med. 2016, 213, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Jiang, H.-Y.; Li, J.; Tang, H.-C.; Zhang, X.-M.; Wang, X.-R.; Du, J.-T.; Li, H.-B.; Xu, G. MicroRNA-23b Promotes Tolerogenic Properties of Dendritic Cells in Vitro through Inhibiting Notch1/NF-ΚB Signalling Pathways. Allergy 2012, 67, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, C.; Olive, V.; Lykken, E.; Feng, F.; Sevilla, J.; Wan, Y.; He, L.; Li, Q.-J. Molecular Dissection of the MiR-17-92 Cluster’s Critical Dual Roles in Promoting Th1 Responses and Preventing Inducible Treg Differentiation. Blood 2011, 118, 5487–5497. [Google Scholar] [CrossRef] [PubMed]
- Baumjohann, D.; Kageyama, R.; Clingan, J.M.; Morar, M.M.; Patel, S.; de Kouchkovsky, D.; Bannard, O.; Bluestone, J.A.; Matloubian, M.; Ansel, K.M.; et al. The MicroRNA Cluster MiR-17∼92 Promotes TFH Cell Differentiation and Represses Subset-Inappropriate Gene Expression. Nat. Immunol. 2013, 14, 840–848. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Zhao, J.L.; Rao, D.S. MicroRNA Function in Myeloid Biology. Blood 2011, 118, 2960–2969. [Google Scholar] [CrossRef] [Green Version]
- Möhnle, P.; Schütz, S.V.; van der Heide, V.; Hübner, M.; Luchting, B.; Sedlbauer, J.; Limbeck, E.; Hinske, L.C.; Briegel, J.; Kreth, S. MicroRNA-146a Controls Th1-Cell Differentiation of Human CD4+ T Lymphocytes by Targeting PRKCε. Eur. J. Immunol. 2015, 45, 260–272. [Google Scholar] [CrossRef]
- Lu, L.-F.; Boldin, M.P.; Chaudhry, A.; Lin, L.-L.; Taganov, K.D.; Hanada, T.; Yoshimura, A.; Baltimore, D.; Rudensky, A.Y. Function of MiR-146a in Controlling Treg Cell-Mediated Regulation of Th1 Responses. Cell 2010, 142, 914–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.-F.; Thai, T.-H.; Calado, D.P.; Chaudhry, A.; Kubo, M.; Tanaka, K.; Loeb, G.B.; Lee, H.; Yoshimura, A.; Rajewsky, K.; et al. Foxp3-Dependent MicroRNA155 Confers Competitive Fitness to Regulatory T Cells by Targeting SOCS1 Protein. Immunity 2009, 30, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Vigorito, E.; Clare, S.; Warren, M.V.; Couttet, P.; Soond, D.R.; van Dongen, S.; Grocock, R.J.; Das, P.P.; Miska, E.A.; et al. Requirement of Bic/MicroRNA-155 for Normal Immune Function. Science 2007, 316, 608–611. [Google Scholar] [CrossRef] [Green Version]
- Neudecker, V.; Haneklaus, M.; Jensen, O.; Khailova, L.; Masterson, J.C.; Tye, H.; Biette, K.; Jedlicka, P.; Brodsky, K.S.; Gerich, M.E.; et al. Myeloid-Derived MiR-223 Regulates Intestinal Inflammation via Repression of the NLRP3 Inflammasome. J. Exp. Med. 2017, 214, 1737–1752. [Google Scholar] [CrossRef]
- Ghorbani, S.; Talebi, F.; Chan, W.F.; Masoumi, F.; Vojgani, M.; Power, C.; Noorbakhsh, F. MicroRNA-181 Variants Regulate T Cell Phenotype in the Context of Autoimmune Neuroinflammation. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wang, F.-L.; Wang, H.-B.; Dong, N.; Zhu, X.-M.; Wu, Y.; Wang, Y.-T.; Yao, Y.-M. TNF-α MRNA Is Negatively Regulated by MicroRNA-181a-5p in Maturation of Dendritic Cells Induced by High Mobility Group Box-1 Protein. Sci. Rep. 2017, 7, 12239. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.; Zeng, X.; Zhao, L.; Wei, Q.; Yu, L.; Wang, X.; Yu, Z.; Cao, Y.; Shan, F.; Wei, M. MiR-181a Induces Macrophage Polarized to M2 Phenotype and Promotes M2 Macrophage-Mediated Tumor Cell Metastasis by Targeting KLF6 and C/EBPα. Mol. Ther. Nucleic Acids 2016, 5, e368. [Google Scholar] [CrossRef] [Green Version]
- Cortez, M.A.; Anfossi, S.; Ramapriyan, R.; Menon, H.; Atalar, S.C.; Aliru, M.; Welsh, J.; Calin, G.A. Role of MiRNAs in Immune Responses and Immunotherapy in Cancer. Genes Chromosomes Cancer 2019, 58, 244–253. [Google Scholar] [CrossRef]
- Fontana, L.; Pelosi, E.; Greco, P.; Racanicchi, S.; Testa, U.; Liuzzi, F.; Croce, C.M.; Brunetti, E.; Grignani, F.; Peschle, C. MicroRNAs 17-5p-20a-106a Control Monocytopoiesis through AML1 Targeting and M-CSF Receptor Upregulation. Nat. Cell Biol. 2007, 9, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Schmeier, S.; MacPherson, C.R.; Essack, M.; Kaur, M.; Schaefer, U.; Suzuki, H.; Hayashizaki, Y.; Bajic, V.B. Deciphering the Transcriptional Circuitry of MicroRNA Genes Expressed during Human Monocytic Differentiation. BMC Genom. 2009, 10, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aras, S.; Zaidi, M.R. TAMeless Traitors: Macrophages in Cancer Progression and Metastasis. Br. J. Cancer 2017, 117, 1583–1591. [Google Scholar] [CrossRef] [Green Version]
- Self-Fordham, J.B.; Naqvi, A.R.; Uttamani, J.R.; Kulkarni, V.; Nares, S. MicroRNA: Dynamic Regulators of Macrophage Polarization and Plasticity. Front. Immunol. 2017, 8, 1062. [Google Scholar] [CrossRef] [Green Version]
- Trotta, R.; Chen, L.; Ciarlariello, D.; Josyula, S.; Mao, C.; Costinean, S.; Yu, L.; Butchar, J.P.; Tridandapani, S.; Croce, C.M.; et al. MiR-155 Regulates IFN-γ Production in Natural Killer Cells. Blood 2012, 119, 3478–3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Gu, Y.; Zhang, Q.; Han, Y.; Hou, J.; Lin, L.; Wu, C.; Bao, Y.; Su, X.; Jiang, M.; et al. Identification of Resting and Type I IFN-Activated Human NK Cell MiRNomes Reveals MicroRNA-378 and MicroRNA-30e as Negative Regulators of NK Cell Cytotoxicity. J. Immunol. 2012, 189, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Perez, L.G.; Kempski, J.; McGee, H.M.; Pelzcar, P.; Agalioti, T.; Giannou, A.; Konczalla, L.; Brockmann, L.; Wahib, R.; Xu, H.; et al. TGF-β Signaling in Th17 Cells Promotes IL-22 Production and Colitis-Associated Colon Cancer. Nat. Commun. 2020, 11, 2608. [Google Scholar] [CrossRef]
- Kosaka, A.; Ohkuri, T.; Ikeura, M.; Kohanbash, G.; Okada, H. Transgene-Derived Overexpression of MiR-17-92 in CD8+ T-Cells Confers Enhanced Cytotoxic Activity. Biochem. Biophys. Res. Commun. 2015, 458, 549–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Xue, Z.; Liu, Y.; Liu, H.; Guo, X.; Li, Y.; Yang, H.; Zhang, L.; Da, Y.; Yao, Z.; et al. MicroRNA-181c Promotes Th17 Cell Differentiation and Mediates Experimental Autoimmune Encephalomyelitis. Brain Behav. Immun. 2018, 70, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Gracias, D.T.; Stelekati, E.; Hope, J.L.; Boesteanu, A.C.; Doering, T.A.; Norton, J.; Mueller, Y.M.; Fraietta, J.A.; Wherry, E.J.; Turner, M.; et al. The MicroRNA MiR-155 Controls CD8(+) T Cell Responses by Regulating Interferon Signaling. Nat. Immunol. 2013, 14, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Bodor, J.N.; Boumber, Y.; Borghaei, H. Biomarkers for Immune Checkpoint Inhibition in Non–Small Cell Lung Cancer (NSCLC). Cancer 2020, 126, 260–270. [Google Scholar] [CrossRef]
- Rompré-Brodeur, A.; Shinde-Jadhav, S.; Ayoub, M.; Piccirillo, C.A.; Seuntjens, J.; Brimo, F.; Mansure, J.J.; Kassouf, W. PD-1/PD-L1 Immune Checkpoint Inhibition with Radiation in Bladder Cancer: In Situ and Abscopal Effects. Mol. Cancer Ther. 2020, 19, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zhang, Y.; Sun, B.; McMahon, A.P.; Wang, Y. Hedgehog Signaling: From Basic Biology to Cancer Therapy. Cell Chem. Biol. 2017, 24, 252–280. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 Pathway: Current Researches in Cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar] [PubMed]
- Yang, J.; Zhang, Z.; Chen, C.; Liu, Y.; Si, Q.; Chuang, T.-H.; Li, N.; Gomez-Cabrero, A.; Reisfeld, R.A.; Xiang, R.; et al. MicroRNA-19a-3p Inhibits Breast Cancer Progression and Metastasis by Inducing Macrophage Polarization through Downregulated Expression of Fra-1 Proto-Oncogene. Oncogene 2014, 33, 3014–3023. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Han, J.; Cui, Y.; Zhou, X.; Fan, K. MiRNA-21 Inhibition Enhances RANTES and IP-10 Release in MCF-7 via PIAS3 and STAT3 Signalling and Causes Increased Lymphocyte Migration. Biochem. Biophys. Res. Commun. 2013, 439, 384–389. [Google Scholar] [CrossRef]
- Ma, S.; Liu, M.; Xu, Z.; Li, Y.; Guo, H.; Ge, Y.; Liu, Y.; Zheng, D.; Shi, J. A Double Feedback Loop Mediated by MicroRNA-23a/27a/24-2 Regulates M1 versus M2 Macrophage Polarization and Thus Regulates Cancer Progression. Oncotarget 2016, 7, 13502–13519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, P.; Sun, T.; Li, D.; Xu, X.; Rui, Y.; Li, C.; Chong, M.; Ibrahim, T.; Mercatali, L.; et al. MiR-126 and MiR-126* Repress Recruitment of Mesenchymal Stem Cells and Inflammatory Monocytes to Inhibit Breast Cancer Metastasis. Nat. Cell Biol. 2013, 15, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Zhang, H.-W.; Lu, M.-H.; He, X.-H.; Li, Y.; Gu, H.; Liu, M.-F.; Wang, E.-D. MicroRNA-155 Functions as an OncomiR in Breast Cancer by Targeting the Suppressor of Cytokine Signaling 1 Gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lai, L.; Chen, Q.; Song, Y.; Xu, S.; Ma, F.; Wang, X.; Wang, J.; Yu, H.; Cao, X.; et al. MicroRNA-494 Is Required for the Accumulation and Functions of Tumor-Expanded Myeloid-Derived Suppressor Cells via Targeting of PTEN. J. Immunol. 2012, 188, 5500–5510. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Liu, M.; Li, Y.; Nie, Y.; Mi, Q.; Zhao, S. Ovarian Tumor-Associated MicroRNA-20a Decreases Natural Killer Cell Cytotoxicity by Downregulating MICA/B Expression. Cell. Mol. Immunol. 2014, 11, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Tao, Z.; Hai, B.; Liang, H.; Shi, Y.; Wang, T.; Song, W.; Chen, Y.; OuYang, J.; Chen, J.; et al. MiR-424(322) Reverses Chemoresistance via T-Cell Immune Response Activation by Blocking the PD-L1 Immune Checkpoint. Nat. Commun. 2016, 7, 11406. [Google Scholar] [CrossRef]
- Chen, R.; Alvero, A.B.; Silasi, D.A.; Kelly, M.G.; Fest, S.; Visintin, I.; Leiser, A.; Schwartz, P.E.; Rutherford, T.; Mor, G. Regulation of IKKbeta by MiR-199a Affects NF-KappaB Activity in Ovarian Cancer Cells. Oncogene 2008, 27, 4712–4723. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Li, Q.-J.; Feng, Y.; Zhang, Y.; Markowitz, G.J.; Ning, S.; Deng, Y.; Zhao, J.; Jiang, S.; Yuan, Y.; et al. TGF-β-MiR-34a-CCL22 Signaling-Induced Treg Cell Recruitment Promotes Venous Metastases of HBV-Positive Hepatocellular Carcinoma. Cancer Cell 2012, 22, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Gupta, S.; Du, W.W.; Yang, B.B. MicroRNA-17 Inhibits Tumor Growth by Stimulating T-Cell Mediated Host Immune Response. Oncoscience 2014, 1, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, F.; Jia, X.; Iwanowycz, S.; Wang, Y.; Huang, S.; Ai, W.; Fan, D. MicroRNA-155 Deficiency Enhances the Recruitment and Functions of Myeloid-Derived Suppressor Cells in Tumor Microenvironment and Promotes Solid Tumor Growth. Int. J. Cancer 2015, 136, E602–E613. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, X.; Li, H.; Yu, J.; Ren, X. The Role of MiRNA-29 Family in Cancer. Eur. J. Cell Biol. 2013, 92, 123–128. [Google Scholar] [CrossRef]
- Yin, Y.; Cai, X.; Chen, X.; Liang, H.; Zhang, Y.; Li, J.; Wang, Z.; Chen, X.; Zhang, W.; Yokoyama, S.; et al. Tumor-Secreted MiR-214 Induces Regulatory T Cells: A Major Link between Immune Evasion and Tumor Growth. Cell Res. 2014, 24, 1164–1180. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gibbons, D.L.; Goswami, S.; Cortez, M.A.; Ahn, Y.-H.; Byers, L.A.; Zhang, X.; Yi, X.; Dwyer, D.; Lin, W.; et al. Metastasis Is Regulated via MicroRNA-200/ZEB1 Axis Control of Tumour Cell PD-L1 Expression and Intratumoral Immunosuppression. Nat. Commun. 2014, 5, 5241. [Google Scholar] [CrossRef]
- Yang, N.; Zhu, S.; Lv, X.; Qiao, Y.; Liu, Y.-J.; Chen, J. MicroRNAs: Pleiotropic Regulators in the Tumor Microenvironment. Front. Immunol. 2018, 9, 2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Chen, L.; Chen, G.; Hu, C.; Jiang, S.; Sevilla, J.; Wan, Y.; Sampson, J.H.; Zhu, B.; Li, Q.-J. Targeting MiR-23a in CD8+ Cytotoxic T Lymphocytes Prevents Tumor-Dependent Immunosuppression. J. Clin. Invest. 2014, 124, 5352–5367. [Google Scholar] [CrossRef]
- Pyfferoen, L.; Mestdagh, P.; Vergote, K.; De Cabooter, N.; Vandesompele, J.; Lambrecht, B.N.; Vermaelen, K.Y. Lung Tumours Reprogram Pulmonary Dendritic Cell Immunogenicity at the MicroRNA Level. Int. J. Cancer 2014, 135, 2868–2877. [Google Scholar] [CrossRef]
- Wei, J.; Wang, F.; Kong, L.-Y.; Xu, S.; Doucette, T.; Ferguson, S.D.; Yang, Y.; McEnery, K.; Jethwa, K.; Gjyshi, O.; et al. MiR-124 Inhibits STAT3 Signaling to Enhance T Cell-Mediated Immune Clearance of Glioma. Cancer Res. 2013, 73, 3913–3926. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, R.; Deng, Y.; Qian, J.; Lu, Z.; Wang, Y.; Zhang, D.; Luo, F.; Chu, Y. MiR-15a/16 Deficiency Enhances Anti-Tumor Immunity of Glioma-Infiltrating CD8+ T Cells through Targeting MTOR. Int. J. Cancer 2017, 141, 2082–2092. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Johnston, N.; Zheng, X.; Wang, H.; Zhang, X.; Gao, D.; Min, W. MiR-28 Modulates Exhaustive Differentiation of T Cells through Silencing Programmed Cell Death-1 and Regulating Cytokine Secretion. Oncotarget 2016, 7, 53735–53750. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Nduom, E.K.; Kong, L.-Y.; Hashimoto, Y.; Xu, S.; Gabrusiewicz, K.; Ling, X.; Huang, N.; Qiao, W.; Zhou, S.; et al. MiR-138 Exerts Anti-Glioma Efficacy by Targeting Immune Checkpoints. Neuro Oncol. 2016, 18, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, M.M.; Fawzy, I.O.; Bassiouni, A.A.; Gomaa, A.I.; Esmat, G.; Waked, I.; Abdelaziz, A.I. Enhancing NK Cell Cytotoxicity by MiR-182 in Hepatocellular Carcinoma. Hum. Immunol. 2016, 77, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Xia, Z.; Deng, Y.-N.; Yang, Y.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. Emerging MicroRNA Biomarkers for Colorectal Cancer Diagnosis and Prognosis. Open Biol. 2019, 9, 180212. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Khan, H.; Ullah, H.; Hassan, S.T.S.; Šmejkal, K.; Efferth, T.; Mahomoodally, M.F.; Xu, S.; Habtemariam, S.; Filosa, R.; et al. MicroRNA Targeting by Quercetin in Cancer Treatment and Chemoprotection. Pharmacol. Res. 2019, 147, 104346. [Google Scholar] [CrossRef]
- Gomarasca, M.; Maroni, P.; Banfi, G.; Lombardi, G. MicroRNAs in the Antitumor Immune Response and in Bone Metastasis of Breast Cancer: From Biological Mechanisms to Therapeutics. Int. J. Mol. Sci. 2020, 21, 2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchie, A. First MicroRNA Mimic Enters Clinic. Nat. Biotechnol. 2013, 31, 577. [Google Scholar] [CrossRef]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. MicroRNA Therapeutics in Cancer—An Emerging Concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantafio, M.E.G.; Nielsen, B.S.; Mignogna, C.; Arbitrio, M.; Botta, C.; Frandsen, N.M.; Rolfo, C.; Tagliaferri, P.; Tassone, P.; Martino, M.T.D. Pharmacokinetics and Pharmacodynamics of a 13-Mer LNA-Inhibitor-MiR-221 in Mice and Non-Human Primates. Mol. Ther. Nucleic Acids 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA Sponges: Competitive Inhibitors of Small RNAs in Mammalian Cells. Nat. Methods 2007, 4, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Van Roosbroeck, K.; Fanini, F.; Setoyama, T.; Ivan, C.; Rodriguez-Aguayo, C.; Fuentes-Mattei, E.; Xiao, L.; Vannini, I.; Redis, R.S.; D’Abundo, L.; et al. Combining Anti-Mir-155 with Chemotherapy for the Treatment of Lung Cancers. Clin. Cancer Res. 2017, 23, 2891–2904. [Google Scholar] [CrossRef] [Green Version]
- Paranjape, T.; Slack, F.J.; Weidhaas, J.B. MicroRNAs: Tools for Cancer Diagnostics. Gut 2009, 58, 1546–1554. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.A.; Ivan, C.; Valdecanas, D.; Wang, X.; Peltier, H.J.; Ye, Y.; Araujo, L.; Carbone, D.P.; Shilo, K.; Giri, D.K.; et al. PDL1 Regulation by P53 via MiR-34. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef] [Green Version]
- Wiggins, J.F.; Ruffino, L.; Kelnar, K.; Omotola, M.; Patrawala, L.; Brown, D.; Bader, A.G. Development of a Lung Cancer Therapeutic Based on the Tumor Suppressor MicroRNA-34. Cancer Res. 2010, 70, 5923–5930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs Bind to Toll-like Receptors to Induce Prometastatic Inflammatory Response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Deng, Z.; Zhao, W.; Cao, Z. Searching Synergistic Dose Combinations for Anticancer Drugs. Front. Pharmacol. 2018, 9, 535. [Google Scholar] [CrossRef] [PubMed]
- Lakhal, S.; Wood, M.J.A. Exosome Nanotechnology: An Emerging Paradigm Shift in Drug Delivery: Exploitation of Exosome Nanovesicles for Systemic in Vivo Delivery of RNAi Heralds New Horizons for Drug Delivery across Biological Barriers. Bioessays 2011, 33, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.C.; Chan, W.C.W. Nanotoxicity: The Growing Need for in Vivo Study. Curr. Opin. Biotechnol. 2007, 18, 565–571. [Google Scholar] [CrossRef] [PubMed]



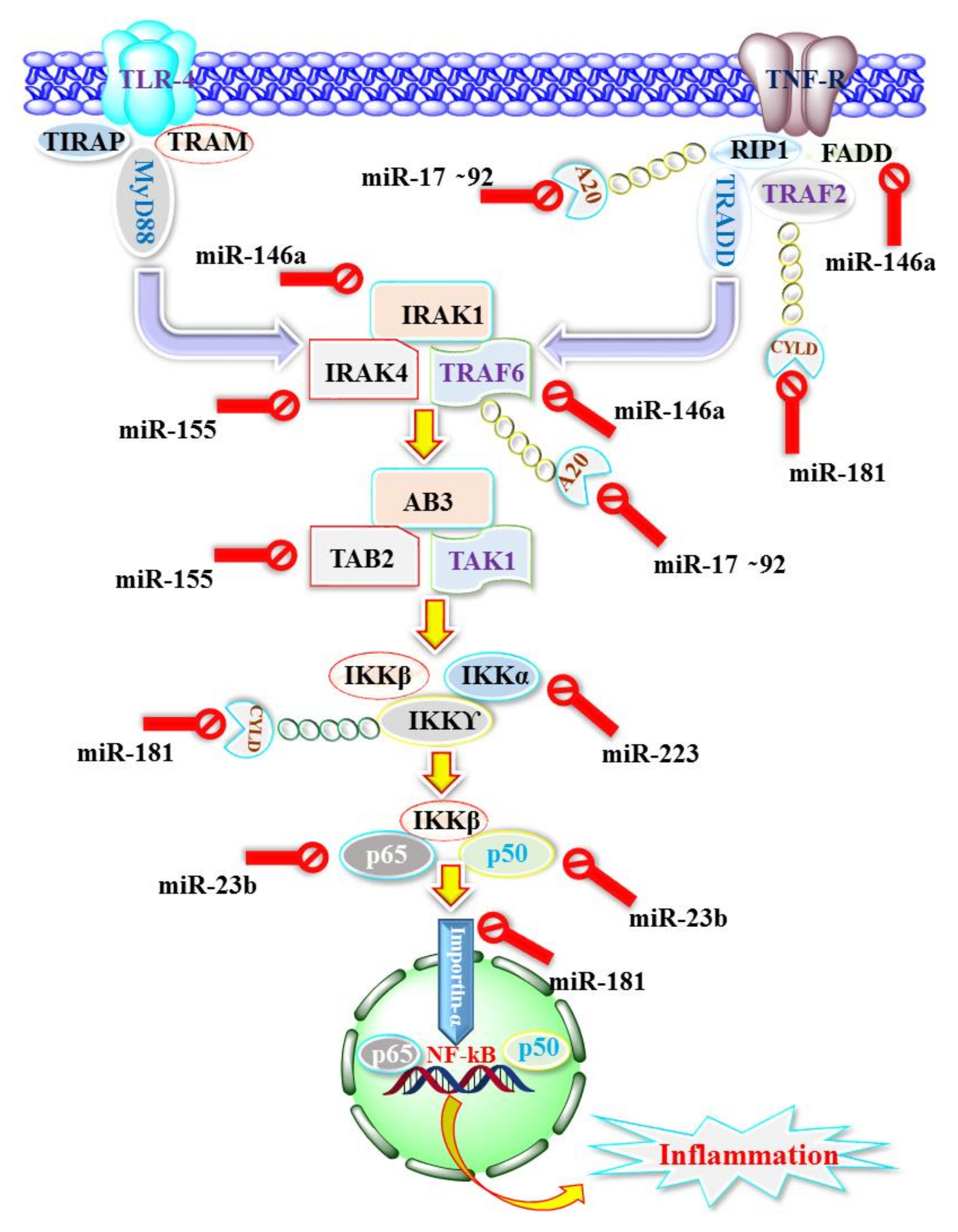{kind=link}
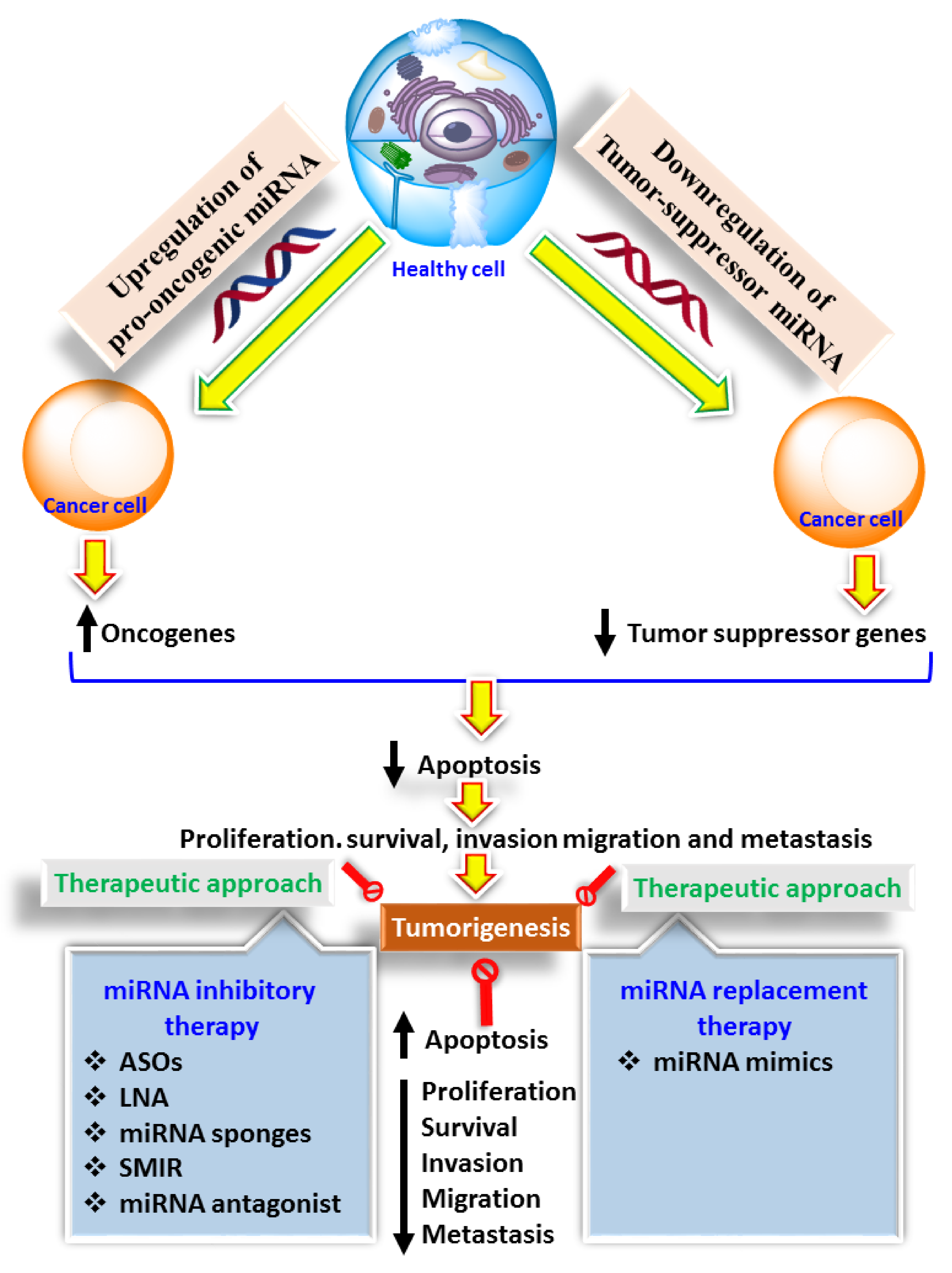{kind=link}
| S. No. | Type of Cancer | miRNAs Involved | References | |
|---|---|---|---|---|
| 1 | Acute Myeloid Leukemia | miR-196b; miR-126; miR-9; miR-17-92 | Oncogenic | [59] |
| 2 | Gastric | miR-421; miR-196a | Oncogenic | [56,57] |
| 3 | Breast | miR-181a/b, miR-21 miR-34, miR-34 b/c | Oncogenic Tumor Suppressor | [60,61] |
| 4 | Non-Small Cell Lung Cancer | miR-27a | Oncogenic | [62] |
| 5 | Renal Cell Carcinomas | miR-30a/c | Oncogenic | [63,64] |
| 6 | Colorectal | miR-34a, miR-34b, miR34b/c | Tumor suppressor | [65,66,67] |
| 7 | Prostate | miR-34a, miR-34b, miR34b/c | Tumor suppressor | [68,69] |
| 8 | Lung | miR-34a, miR-34b, miR34b/c | Tumor Suppressor | [70,71,72] |
| 9 | Liver | miR-34b | Tumor Suppressor | [73] |
| 10 | Ostosarcoma | miR-34a, miR34b, miR-192 | Tumor Suppressor | [74,75,76] |
| 11 | haematological neoplasms | miR-34a, miR-34b-5p, miR-34c, miR-34b/c | Tumor Suppressor | [77,78,79] |
| 12 | Lymphophytic leukemia | miR-15/16 | Tumor Suppressor | [80] |
| Sl. No. | Type of Cancer | miRNAs Biomarkers | ||
|---|---|---|---|---|
| Predictive | Prognostic | Diagnostic | ||
| 1 | Prostate cancer | miR-21 | miR-20a; miR -21; miR-141 | miR-141, -375; let-7c, -7e miR-141 |
| 2 | Lung cancer | miR-128b | miR-221; let-7a, -137; miR-372; miR -182; miR-15b, -16 | miR-16, miR-17, -19b, miR-200 family; miR-29c, -30c |
| 3 | Breast cancer | miR-125b | miR-210; miR-10b | miR-21, -30a; miR-141, -145; miR- 801 |
| 4 | Ovarian cancer | miR-181a,b, -213; miR-23a, -27a; let-7g, 3p | miR-200 family; miR-410, -645 | miR-126, -127; miR-200 & let-7 family; miR-21, -29a |
| 5 | Liver cancer | miR-21, -200b | miR-200 family; miR-21, -22, -26 | miR-200c, -203, -224; miR-222, -223 |
| miRNA | Effect/Mechanism | Cancer Types |
|---|---|---|
| miR-24-3p | Increased T reg cells activity and reduced Th17 proliferation via targeting fibroblast growth factor 11 | Nasopharyngeal cancer |
| miR-183 | Reduced the activity of NK cells | Lung cancer |
| miR-23a | Reduced activity of CD8+ cells via acting on B lymphocyte-induced maturation protein-1 | |
| miR-155 | Reduced activity of MDSCs via acing on hypoxia inducing factor-α | pancreatic cancer |
| miR-212-3p | Increased immune response of DC via acting on Regulatory Factor X Associated Protein | |
| miR-92a-3p | Increased tumor-associated macrophages and IL-6 level | Liposarcoma |
| miR-25-3p | Increased tumor-associated macrophages and IL-6 level | |
| miR-155 | Reduced activity of MDSCs via acing on hypoxia inducing factor-α | Skin cancer |
| miR-210 | Reduced activity of MDSCs via acing on C-X-C motif chemokine 12 and IL-16 | |
| miR-34a | Reduced recruitment of T reg cells via acting on C-X-C motif chemokine 12 | Hepatic cancer |
| miR-20a and miR-17-5p | Reduced activity of MDSCs via acing on Reduced activity of MDSCs via acing on C-X-C motif chemokine 12 and IL-16 | Colon cancer |
| miR-494 | Reduced activity of MDSCs via acing on Phosphatase and tensin homolog | Breast cancer |
| miR-222-3p | Increased polarization of M2 macrophage via acting on suppressor of cytokine signaling 3 | Ovarian cancer |
| miRNA | Immune Cells/Cancer Cells | Effect | Cancer Types | References |
|---|---|---|---|---|
| miR-19a-3p | Macrophages (M2) | Polarization of macrophage and amelioration of cancer progression and metastasis | Breast cancer | [162] |
| miR-21 | T reg cells | Reduce the proliferation of T reg cells leading to reduced survival and proliferation | [163] | |
| miR-23a/27a/24-2 | Macrophage | Polarization of M2 macrophage and promote tumour growth | [164] | |
| miR-126 | Cancerous cells | Reduced monocyte recruitment and stimulated metastasis | [165] | |
| miR-146a | Macrophages | Activation of NF-kB and promote tumour cell invasion | [140] | |
| miR-155 | Cancerous cells | Activation of JAK-STAT pathway and proliferation of tumor cells. | [166] | |
| miR-223 | Macrophages (M2) | Differentiation of macrophage and stimulate tumour cells invasion | [88] | |
| miR-494 | MDSCs | MDSCs accumulation stimulate tumor cells invasion and metastasis | [167] | |
| miR-20a | Cancerous cells | Suppress NK-mediated antitumor effect and promote tumour invasion as well as proliferation | Ovarian cancer | [168] |
| miR-424 | Cancerous cells | Activate T cells and stimulate sensitivity of tucells towards chemotherapy | [169] | |
| miR-199a | Cancerous cells | Production of cytokines and progression of tumorigenesis | [170] | |
| miR-34a/c | Cancerous cells | Suppress NK-mediated antitumor effect and promote tumour invasion as well as proliferation | Skin cancer | [171] |
| miR-17 | T cells | Alters the function of T cells and promote tumour growth | [172] | |
| miR-155 | MDSCs | Increased HIF-α and promote tumor growth | [173] | |
| miR-29 | Cancer cells | Mitigate the NK cell’s function, T cells and promote the growth of tumors | Solid tumors | [174] |
| miR-214 | CD4+ and CD25+ T cells | Stimulate T reg cells and promote tumour growth | [175] | |
| miR-146a | Cancer cells | Reduced IL-8, TRAF-6 and exhibit the antitumor effect | Gastric cancer | [176] |
| miR-5, 18 and 22 | Cancer cells | Reduced PD-1 expression 6 and exhibit antitumor effect | [177] | |
| miR-23a | T cells | Inhibit CD8+ function and promote tumour growth and TGF-β induced tumour invasion | Lung cancer | [178] |
| miR-155 | Dendritic cell | Stimulate dendritic cell maturation and activation of T cells | [179] | |
| miR-124 | T cells | Inhibit STAT-3 and promote T cell-induced killing of tumour cells | Glioblastoma | [180] |
| miR-15a and 16a | T cells | Reduced PD-1 and increased CD8+ mediated antitumor effect | [181] | |
| miR-28 | T cells | Enervation of T cells and reduced PD-1 | [182] | |
| miR-138 | T cells | Diminished expression of PD-1 and exhibit the anti-tumor effect | [183] | |
| miR-182 | NK cells | Stimulate the tumour-killing potential of NK cells and increase the release of perforin-1 | Hepatic cancer | [184] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pottoo, F.H.; Iqubal, A.; Iqubal, M.K.; Salahuddin, M.; Rahman, J.U.; AlHajri, N.; Shehadeh, M. miRNAs in the Regulation of Cancer Immune Response: Effect of miRNAs on Cancer Immunotherapy. Cancers 2021, 13, 6145. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13236145
Pottoo FH, Iqubal A, Iqubal MK, Salahuddin M, Rahman JU, AlHajri N, Shehadeh M. miRNAs in the Regulation of Cancer Immune Response: Effect of miRNAs on Cancer Immunotherapy. Cancers. 2021; 13(23):6145. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13236145
Chicago/Turabian StylePottoo, Faheem Hyder, Ashif Iqubal, Mohammad Kashif Iqubal, Mohammed Salahuddin, Jawad Ur Rahman, Noora AlHajri, and Mustafa Shehadeh. 2021. "miRNAs in the Regulation of Cancer Immune Response: Effect of miRNAs on Cancer Immunotherapy" Cancers 13, no. 23: 6145. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13236145