
Development of Novel Follicular Thyroid Cancer Models Which Progress to Poorly Differentiated and Anaplastic Thyroid Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
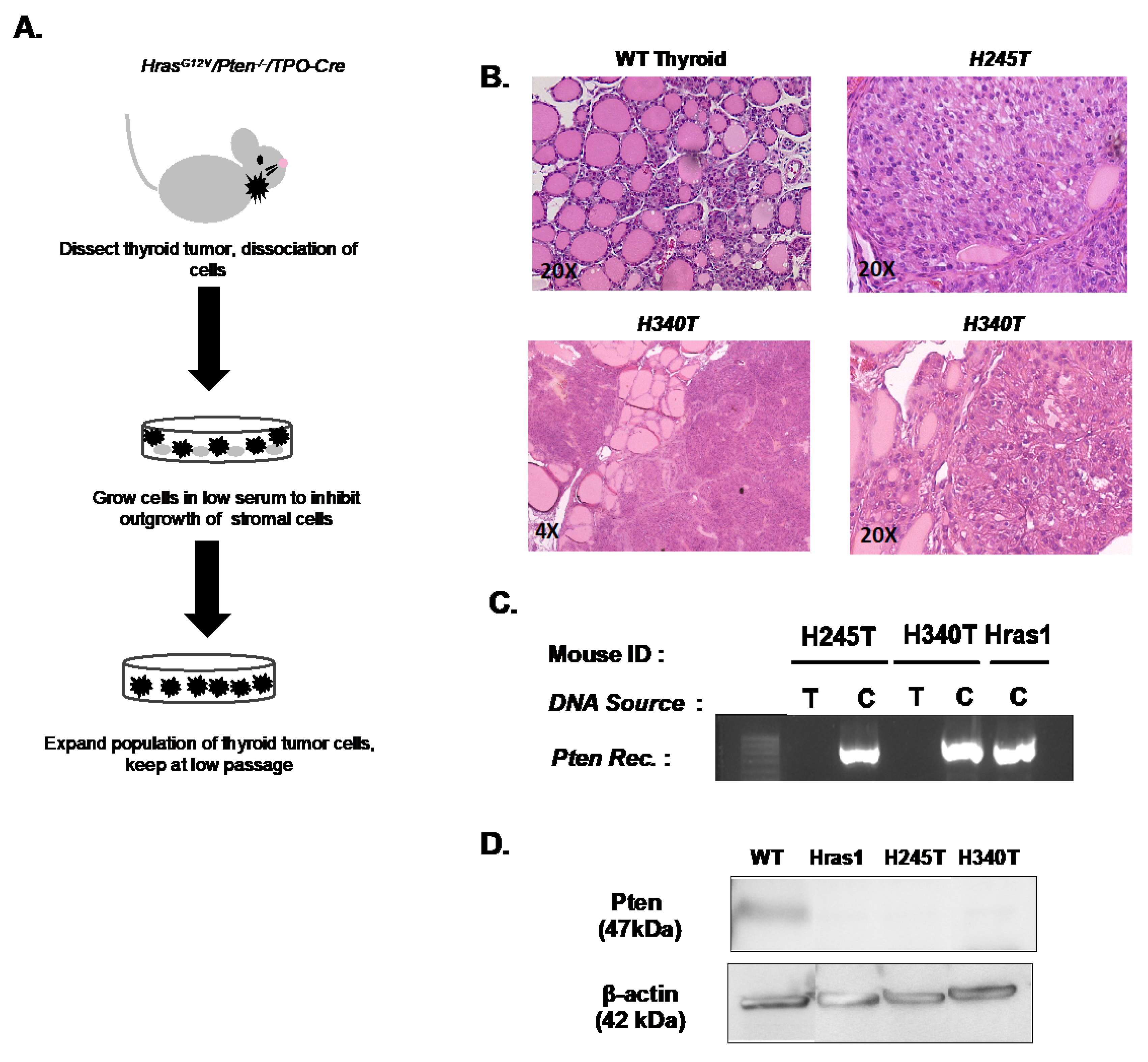
2.1. Derivation of Murine Thyroid Tumor Cell Lines and Wild Type Thyrocyte Cultures
2.2. Immunofluorescence
2.3. RT-PCR Analysis
2.4. Western Blot Analysis
2.5. Proliferation Assays
2.6. Cytotoxicity Assay
2.7. Cell Cycle Analysis
2.8. Three Dimensional Matrigel Growth Assay
2.9. Hanging Drop Assay
2.10. In Vivo Tumorigenecity Assay
2.11. FACS Analysis
2.12. Conditioned Media Collection
2.13. Proteomics Analysis
2.14. Statistical Analysis
3. Results
3.1. Generation of Stable Tumor Cell Lines from HrasG12V/Pten−/−/TPO Cre Thyroid Tumors
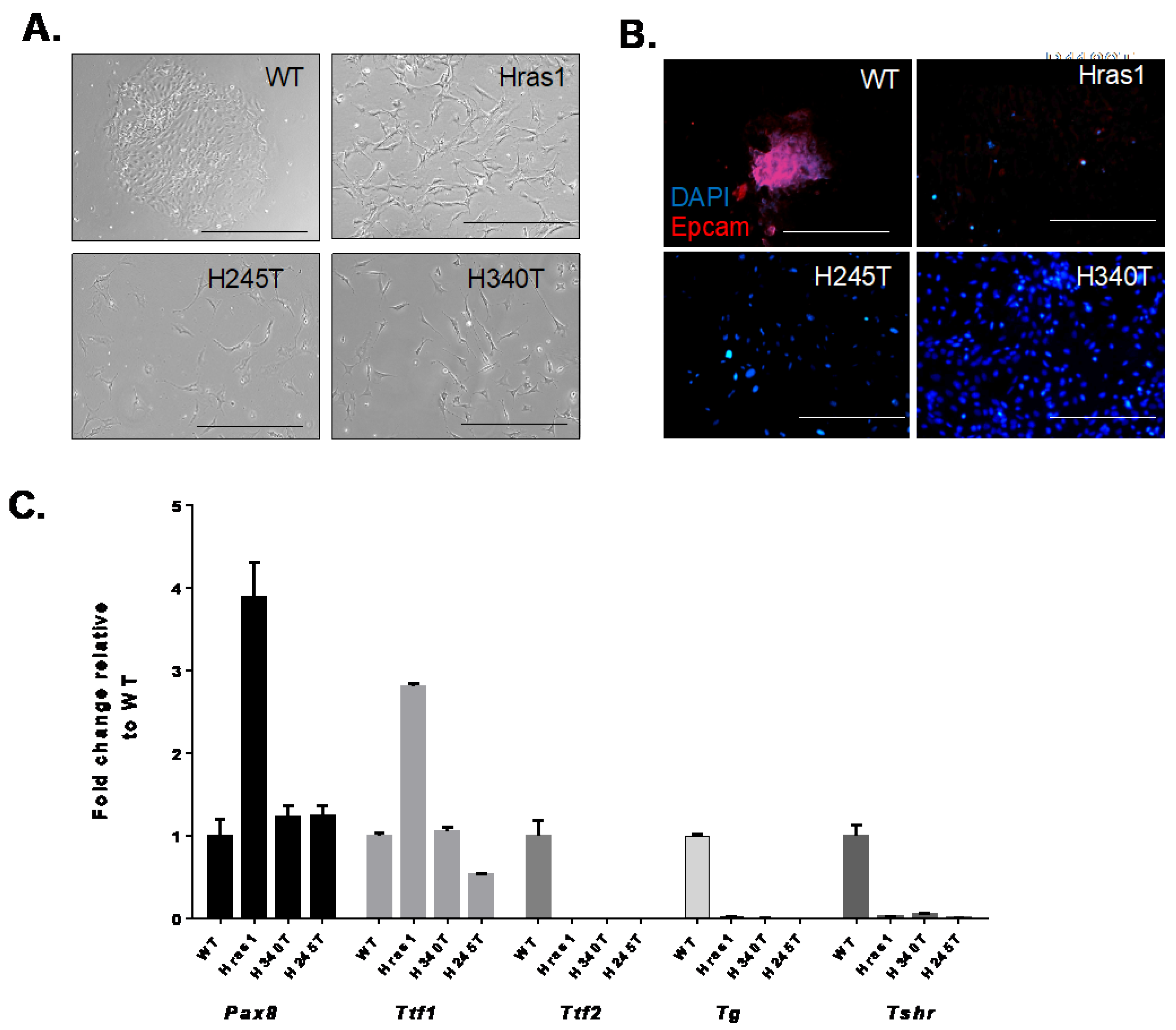
3.2. Morphological Features and Expression of Thyroid-Specific Genes in HrasG12V/Pten−/−/TPO Cre Thyroid Tumor Cell Lines
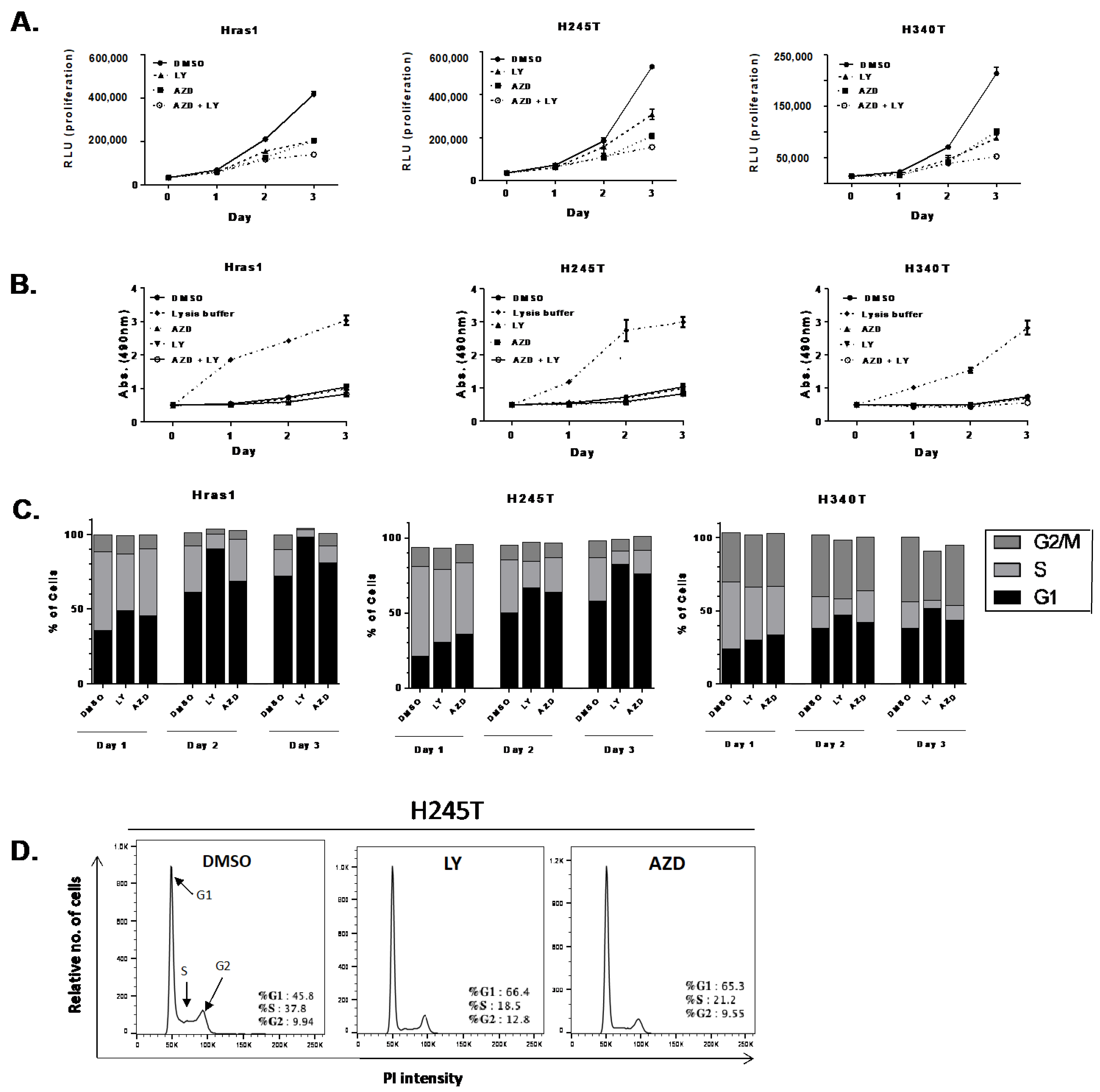
3.3. Proliferation Rates of HrasG12V/Pten−/−/TPO-Cre Cell Lines In Vitro
3.4. Growth Suppressive Effect of Pathway-Specific Inhibitors in Hras-Driven Thyroid Tumor Cell Lines
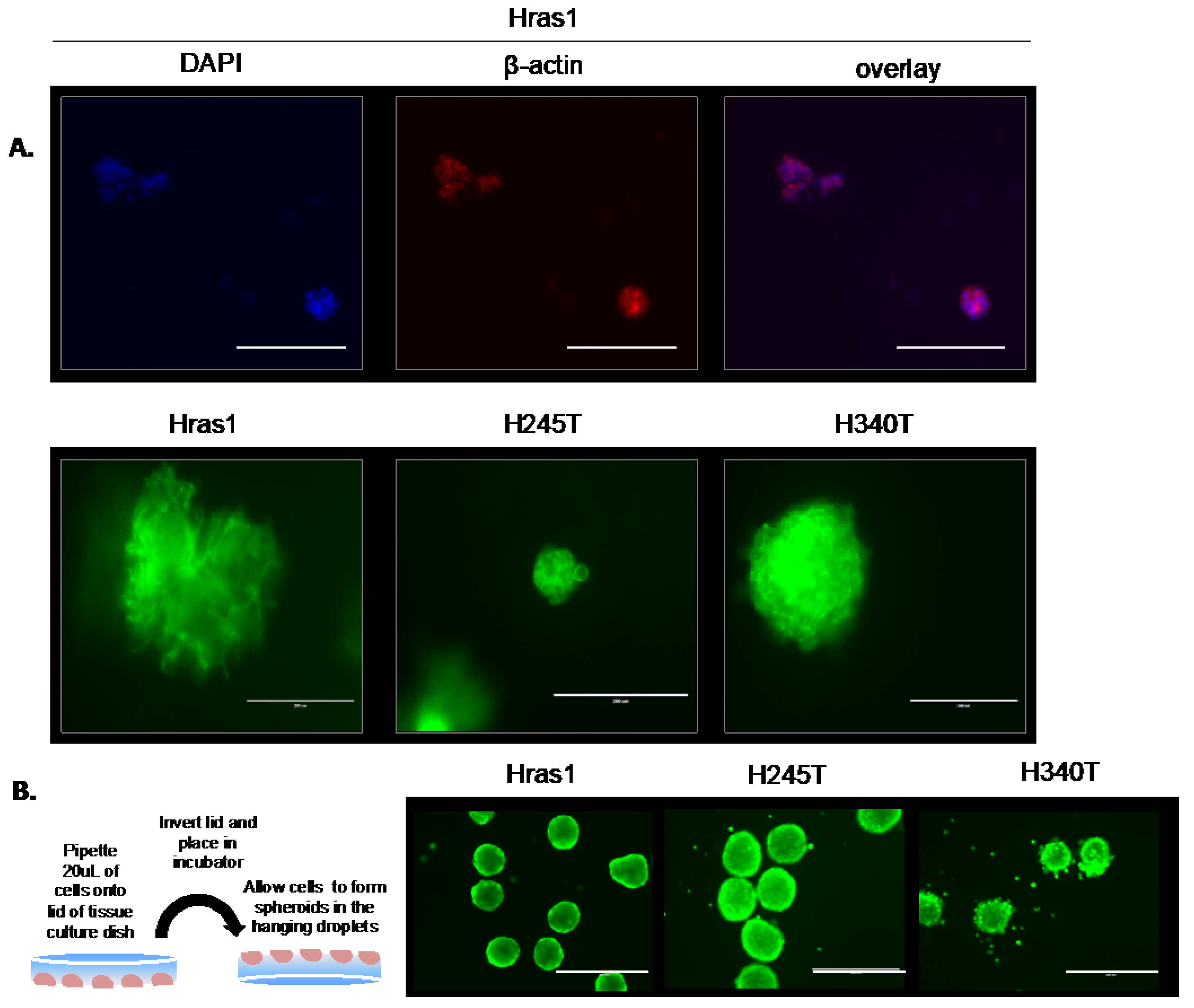
3.5. Development of Physiologically Relevant Three-Dimensional In Vitro Tumor Models
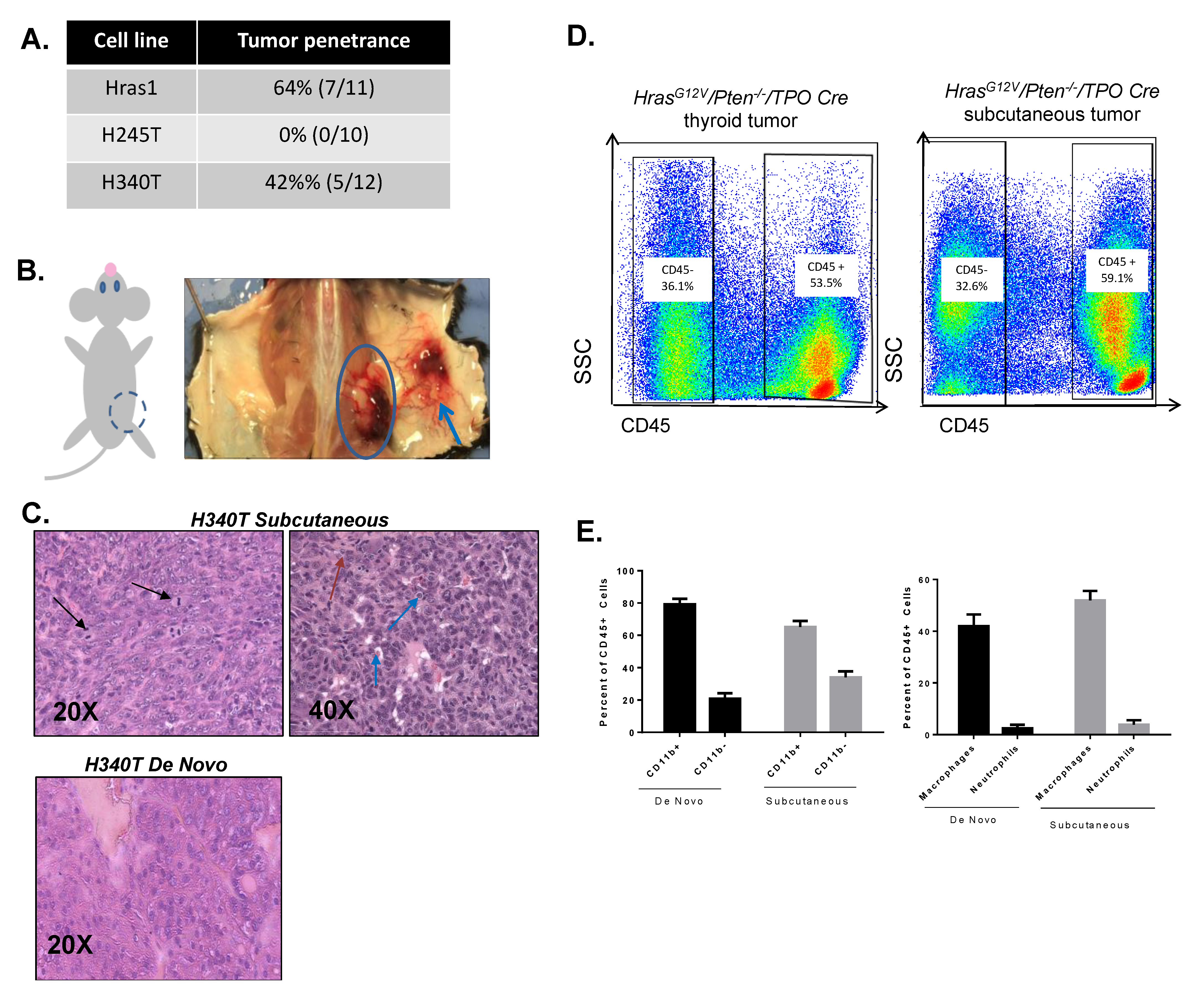
3.6. Development of Syngeneic HrasG12V/Pten−/−/TPO-Cre Tumor Model
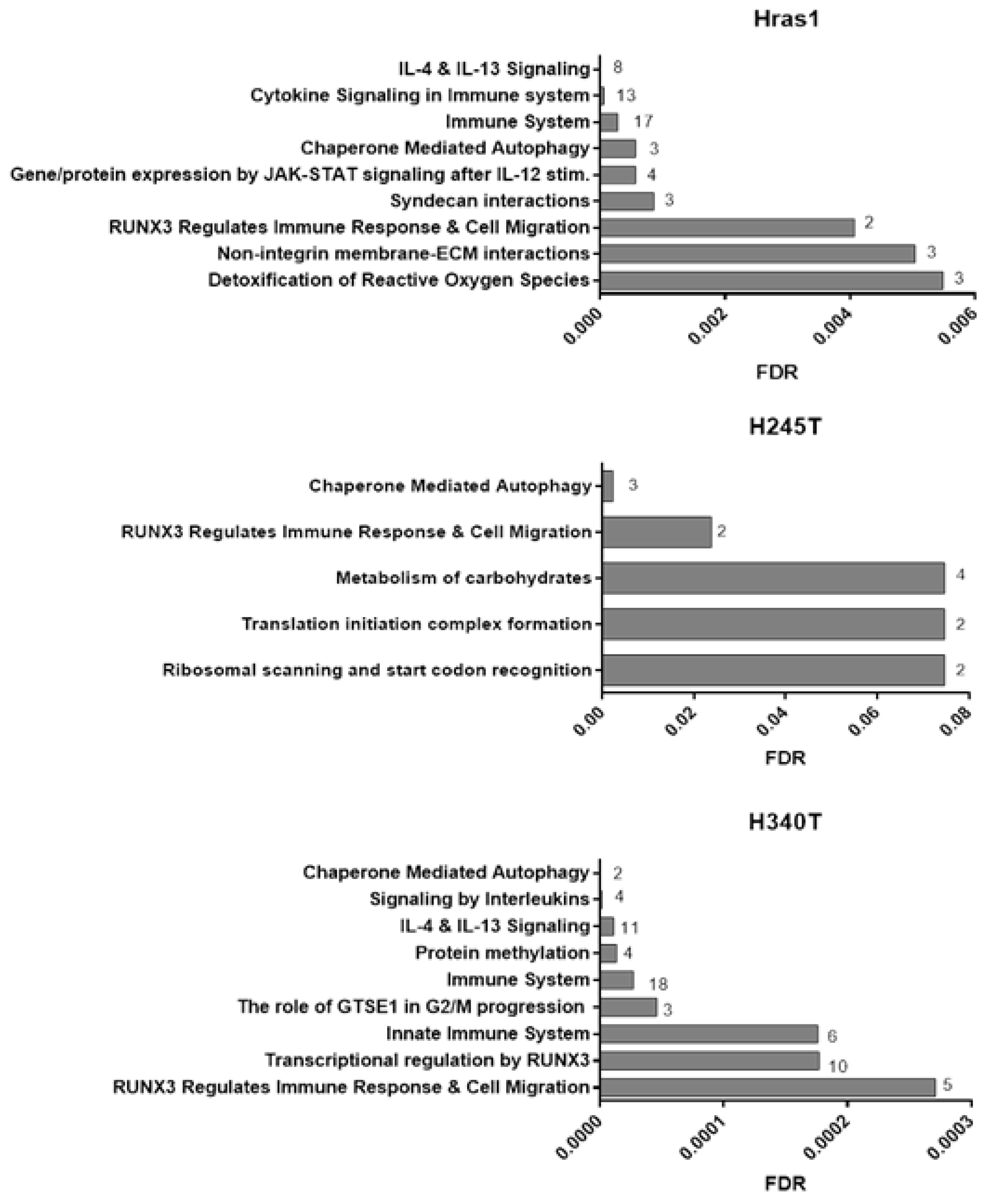
3.7. Proteomic Analysis of Conditioned Media from HrasG12V/Pten−/−/TPO-Cre Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Kitahara, C.M.; Sosa, J.A. Understanding the ever-changing incidence of thyroid cancer. Nat. Rev. Endocrinol. 2020, 16, 1–2. [Google Scholar] [CrossRef]
- Nikiforov, Y.E.; Seethala, R.R.; Tallini, G.; Baloch, Z.W.; Basolo, F.; Thompson, L.D.R.; Barletta, J.A.; Wenig, B.M.; Ghuzlan, A.A.; Kakudo, K.; et al. Nomenclature revision for encapsulated follicular variant of papillary thyroid carcinoma: A paradigm shift to reduce overtreatment of indolent tumors. JAMA Oncol. 2016, 2, 1023–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; et al. SEER Cancer Statistics Review, 1975–2017; National Cancer Institute: Bethesda, MD, USA, November 2019. Available online: https://seer.cancer.gov/csr/1975_2017/ (accessed on 15 December 2020).
- Saini, S.; Tulla, K.; Maker, A.V.; Burman, K.D.; Prabhakar, B.S. Therapeutic advances in anaplastic thyroid cancer: A current perspective. Mol. Cancer 2018, 17, 154. [Google Scholar] [CrossRef] [Green Version]
- Ricarte-Filho, J.C.; Ryder, M.; Chitale, D.A.; Rivera, M.; Heguy, A.; Ladanyi, M.; Janakiraman, M.; Solit, D.; Knauf, J.A.; Tuttle, R.M.; et al. Mutational Profile of Advanced Primary and Metastatic Radioactive Iodine-Refractory Thyroid Cancers Reveals Distinct Pathogenetic Roles for BRAF, PIK3CA, and AKT1. Cancer Res. 2009, 69, 4885–4893. [Google Scholar] [CrossRef] [Green Version]
- García-Rostán, G.; Costa, A.M.; Pereira-Castro, I.; Salvatore, G.; Hernandez, R.; Hermsem, M.J.A.; Herrero, A.; Fusco, A.; Cameselle-Teijeiro, J.; Santoro, M. Mutation of the PIK3CA Gene in Anaplastic Thyroid Cancer. Cancer Res. 2005, 65, 10199–10207. [Google Scholar] [CrossRef] [Green Version]
- Krajewska, J.; Handkiewicz-Junak, D.; Jarzab, B. Sorafenib for the treatment of thyroid cancer: An updated review. Expert Opin. Pharm. 2014, 16, 573–583. [Google Scholar] [CrossRef]
- Subbiah, V.; Kreitman, R.J.; Wainberg, Z.A.; Cho, J.Y.; Schellens, J.H.; Soria, J.C.; Wen, P.Y.; Zielinski, C.; Cabanillas, M.E.; Urbanowitz, G.; et al. Dabrafenib and Trametinib Treatment in Patients with Locally Advanced or Metastatic BRAF V600–Mutant Anaplastic Thyroid Cancer. J. Clin. Oncol. 2018, 36, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Ancker, O.V.; Krüger, M.; Wehland, M.; Infanger, M.; Grimm, D. Multikinase Inhibitor Treatment in Thyroid Cancer. Int. J. Mol. Sci. 2019, 21, 10. [Google Scholar] [CrossRef] [Green Version]
- Cabanillas, M.E.; Ryder, M.; Jimenez, C. Targeted Therapy for Advanced Thyroid Cancer: Kinase Inhibitors and Beyond. Endocr. Rev. 2019, 40, 1573–1604. [Google Scholar] [CrossRef] [Green Version]
- Schweppe, R.E.; Klopper, J.P.; Korch, C.; Pugazhenthi, U.; Benezra, M.; Knauf, J.A.; Fagin, J.A.; Marlow, L.A.; Copland, J.A.; Smallridge, R.C.; et al. Deoxyribonucleic Acid Profiling Analysis of 40 Human Thyroid Cancer Cell Lines Reveals Cross-Contamination Resulting in Cell Line Redundancy and Misidentification. J. Clin. Endocrinol. Metab. 2008, 93, 4331–4341. [Google Scholar] [CrossRef] [Green Version]
- Dadon, T.; Ball, D.; Nelkin, B.D. Misidentification of Putative Medullary Thyroid Cancer Cell Lines RO-H85-1 and RO-D81-1. J. Clin. Endocrinol. Metab. 2013, 98, 954–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, Y.C.; Ahn, S.-H.; Ryu, J.; Chen, Y.; Williams, M.D.; El-Naggar, A.K.; Gagea, M.; Schweppe, R.E.; Haugen, B.R.; Lai, S.Y.; et al. Development and characterization of six new human papillary thyroid carcinoma cell lines. J. Clin. Endocrinol. Metab. 2015, 100, E243–E252. [Google Scholar] [CrossRef] [Green Version]
- Schweppe, R.E.; Pozdeyev, N.; Pike, L.A.; Korch, C.; Zhou, Q.; Sams, S.B.; Sharma, V.; Pugazhenthi, U.; Raeburn, C.; Albuja-Cruz, M.B.; et al. Establishment and Characterization of Four Novel Thyroid Cancer Cell Lines and PDX Models Expressing the RET/PTC1 Rearrangement, BRAFV600E, or RASQ61R as Drivers. Mol. Cancer Res. 2019, 17, 1036–1048. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Hou, P.; Ji, M.; Guan, H.; Studeman, K.; Jensen, K.; Vasko, V.; El-Naggar, A.K.; Xing, M. Highly Prevalent Genetic Alterations in Receptor Tyrosine Kinases and Phosphatidylinositol 3-Kinase/Akt and Mitogen-Activated Protein Kinase Pathways in Anaplastic and Follicular Thyroid Cancers. J. Clin. Endocrinol. Metab. 2008, 93, 3106–3116. [Google Scholar] [CrossRef] [Green Version]
- Volante, M.; Rapa, I.; Gandhi, M.; Bussolati, G.; Giachino, D.; Papotti, M.; Nikiforov, Y.E. RAS Mutations Are the Predominant Molecular Alteration in Poorly Differentiated Thyroid Carcinomas and Bear Prognostic Impact. J. Clin. Endocrinol. Metab. 2009, 94, 4735–4741. [Google Scholar] [CrossRef] [Green Version]
- Dettmer, M.S.; Schmitt, A.; Komminoth, P.; Perren, A. Poorly differentiated thyroid carcinoma: An underdiagnosed entity. Gering differenzierte Schilddrüsenkarzinome: Eine unterdiagnostizierte Entität. Der. Pathol. 2020, 41 (Suppl. S1), 1–8. [Google Scholar] [CrossRef]
- Nikiforova, M.N.; Lynch, R.A.; Biddinger, P.W.; Alexander, E.K.; Dorn, G.W., 2nd; Tallini, G.; Nikiforov, Y.E. RAS point mutations and PAX8-PPAR gamma rearrangement in thyroid tumors: Evidence for distinct molecular pathways in thyroid follicular carcinoma. J. Clin. Endocrinol. Metab. 2003, 88, 2318–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, L.A.; Massoll, N.; Franco, A.T. Immune Suppression Mediated by Myeloid and Lymphoid Derived Immune Cells in the Tumor Microenvironment Facilitates Progression of Thyroid Cancers Driven by HrasG12V and Pten Loss. J. Clin. Cell. Immunol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Jolly, L.A.; Novitskiy, S.; Owens, P.; Massoll, N.; Cheng, N.; Fang, W.; Moses, H.L.; Franco, A.T. Fibroblast-Mediated Collagen Remodeling Within the Tumor Microenvironment Facilitates Progression of Thyroid Cancers Driven by BrafV600E and Pten Loss. Cancer Res. 2016, 76, 1804–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, P.Y.; Janovjak, H.; Miserez, A.R.; Dobbie, Z. Processing of gene expression data generated by quantitative real-time RT-PCR. Biotechniques 2002, 32, 1372–1379. [Google Scholar]
- Watson, J.V.; Chambers, S.H.; Smith, P.J. A pragmatic approach to the analysis of DNA histograms with a definable G1 peak. Cytometry 1987, 8, 1–8. [Google Scholar] [CrossRef]
- Kimura, S. Thyroid-Specific Transcription Factors and Their Roles in Thyroid Cancer. J. Thyroid. Res. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, L.P.; López-Márquez, A.; Santisteban, P. Thyroid transcription factors in development, differentiation and disease. Nat. Rev. Endocrinol. 2015, 11, 29–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, D.; Tang, Y.; Chiriboga, L.; Rivera, M.; A Ghossein, R. Diagnostic utility of thyroid transcription factors Pax8 and TTF-2 (FoxE1) in thyroid epithelial neoplasms. Mod. Pathol. 2007, 21, 192–200. [Google Scholar] [CrossRef]
- Kanjanahattakij, N.; Chayangsu, P.; Kanoksil, W.; Chontong, S.; Sriphrapradang, C. Pitfall in immunohistochemical staining for thyroglobulin in case of thyroid metastasis from lung carcinoma. CytoJournal 2015, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Men, Q.; Su, X.; Chen, W.; Zou, L.; Li, Q.; Song, M.; Ouyang, D.; Chen, Y.; Li, Z.; et al. Downregulated expression of TSHR is associated with distant metastasis in thyroid cancer. Oncol. Lett. 2017, 14, 7506–7512. [Google Scholar] [CrossRef]
- Fagin, J.A.; Wells, S.A. Biologic and Clinical Perspectives on Thyroid Cancer. N. Engl. J. Med. 2016, 375, 1054–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.A.; Yeager, N.; Baker, K.; Liao, X.-H.; Refetoff, S.; Di Cristofano, A. Oncogenic Kras Requires Simultaneous PI3K Signaling to Induce ERK Activation and Transform Thyroid Epithelial Cells In vivo. Cancer Res. 2009, 69, 3689–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caperton, C.O.; Jolly, L.A.; Massoll, N.; Bauer, A.J.; Franco, A.T. Development of Novel Follicular Thyroid Cancer Models Which Progress to Poorly Differentiated and Anaplastic Thyroid Cancer. Cancers 2021, 13, 1094. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13051094
Caperton CO, Jolly LA, Massoll N, Bauer AJ, Franco AT. Development of Novel Follicular Thyroid Cancer Models Which Progress to Poorly Differentiated and Anaplastic Thyroid Cancer. Cancers. 2021; 13(5):1094. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13051094
Chicago/Turabian StyleCaperton, Caitlin O., Lee Ann Jolly, Nicole Massoll, Andrew J. Bauer, and Aime T. Franco. 2021. "Development of Novel Follicular Thyroid Cancer Models Which Progress to Poorly Differentiated and Anaplastic Thyroid Cancer" Cancers 13, no. 5: 1094. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13051094