
Epigenetic-Like Stimulation of Receptor Expression in SSTR2 Transfected HEK293 Cells as a New Therapeutic Strategy

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Chemical Modulation by 5-aza-dC and VPA
2.3. Chemotoxicity of 5-aza-dC and VPA
2.4. Immunohistochemistry
2.5. Irradiation, Radiosynthesis and Dosimetry
2.6. Treatment Design for Immunostaining
2.7. Measurement of Intracellular Lu-177-DOTATATE Radionuclide Uptake after Stimulation
2.8. Radiosensitivity and Radiotoxicity
2.9. Statistical Analysis
3. Results
3.1. Chemotoxicity of 5-aza-dC and VPA
3.2. Immunohistochemical Staining of Stimulated HEKsst2 and HEK Cells
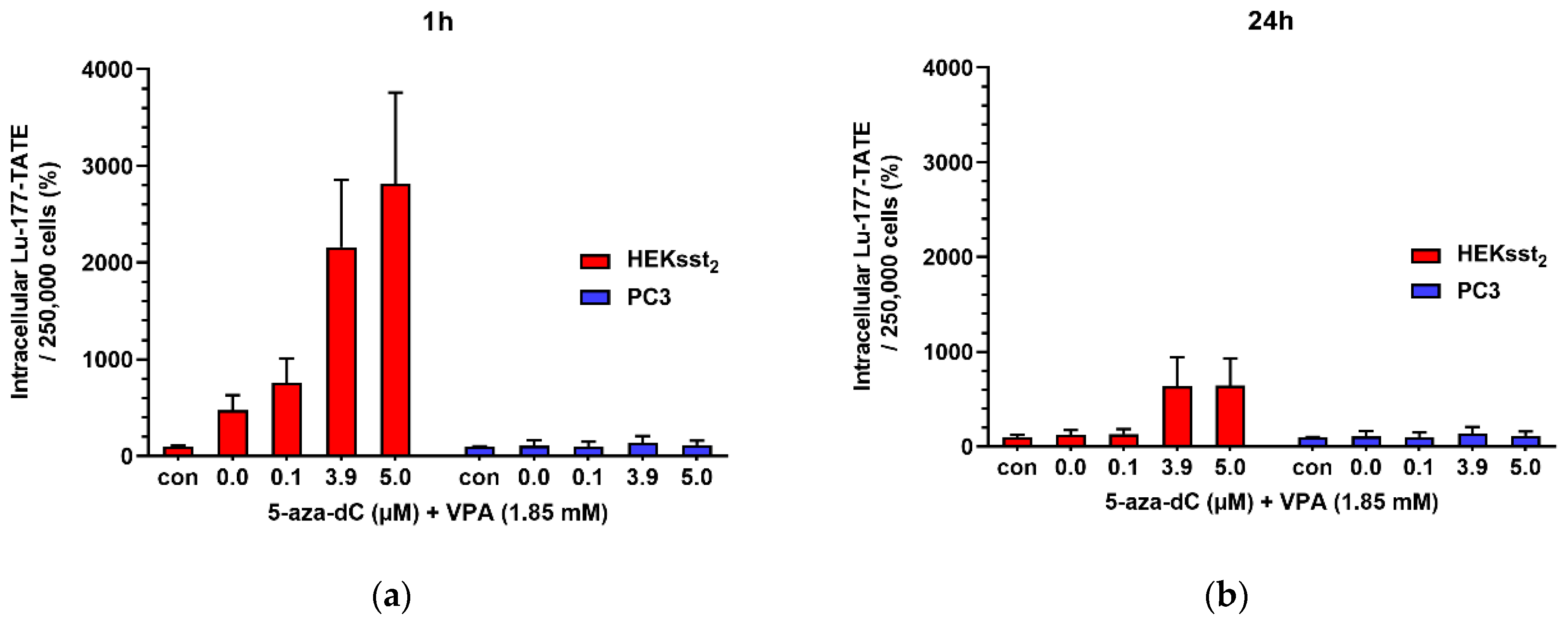
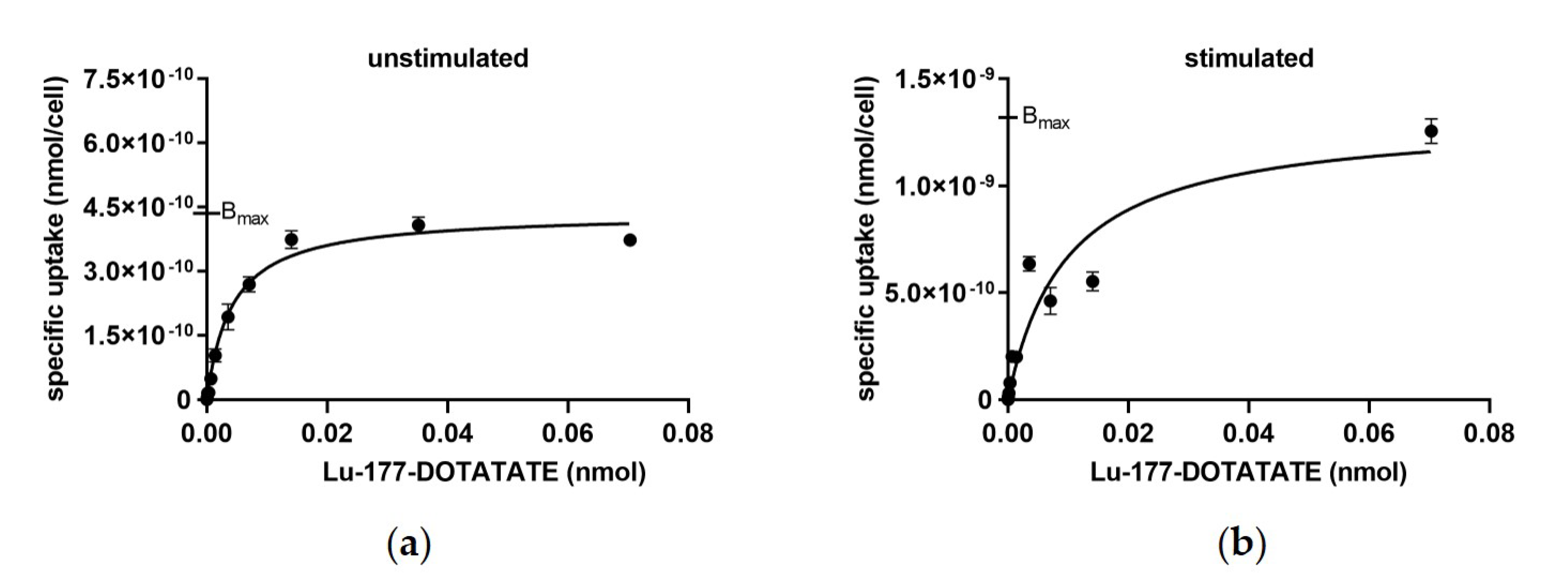
3.3. Uptake and Saturation Binding Assay in Dependency on 5-aza-dC and VPA Stimulation
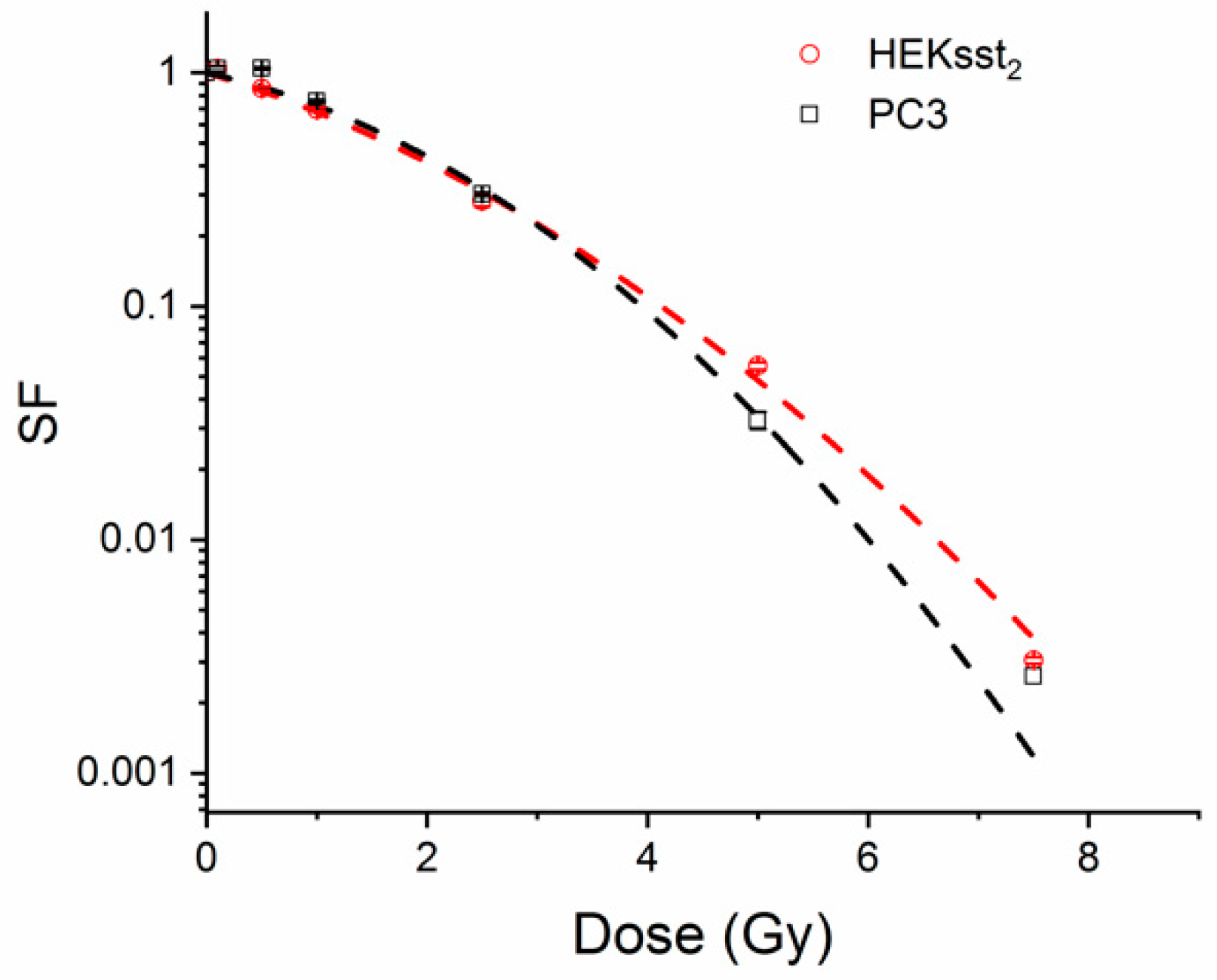
3.4. Radiotoxic Effect of an X-ray on Unstimulated and Stimulated Cells
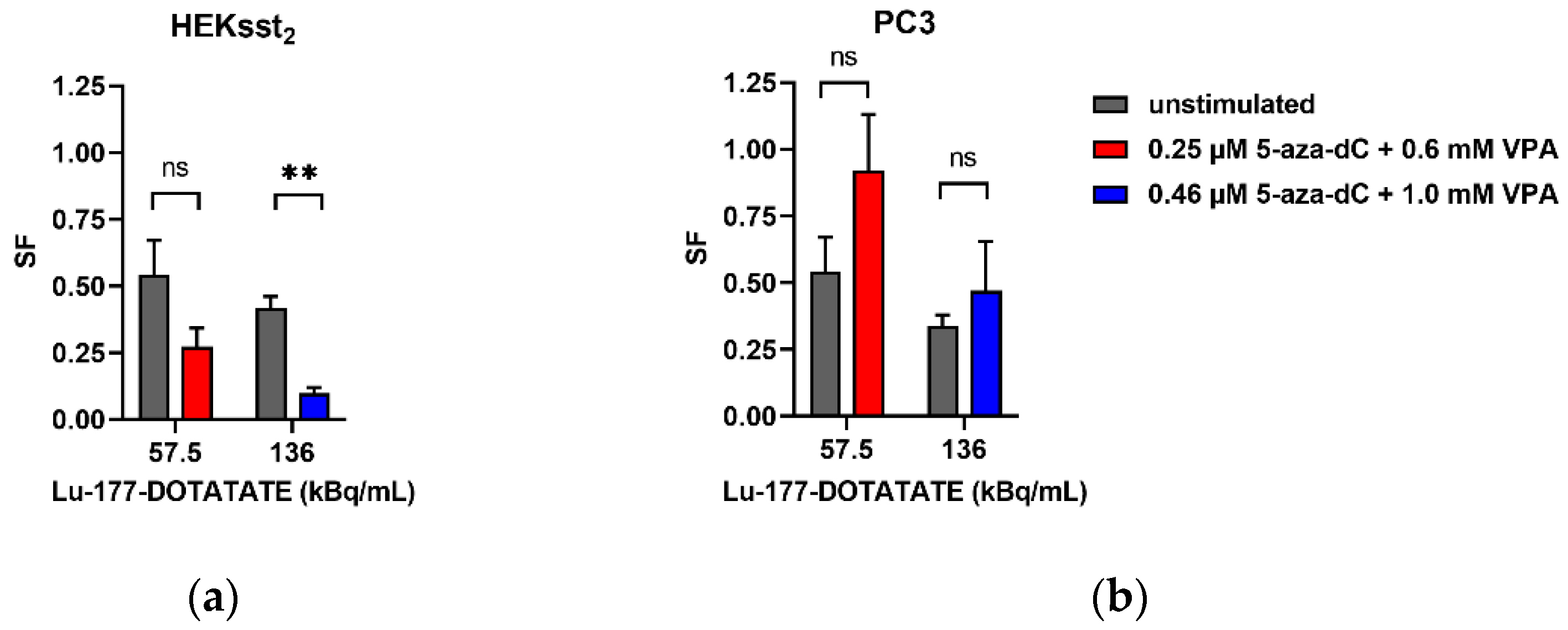
3.5. Radiotoxic Effects of Lu-177-DOTATATE on Unstimulated and Stimulated HEKsst2 and PC3 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reichlin, S. Somatostatin (second of two parts). N. Engl. J. Med. 1983, 309, 1556–1563. [Google Scholar] [CrossRef] [PubMed]
- Ginj, M.; Schmitt, J.S.; Chen, J.; Waser, B.; Reubi, J.C.; de Jong, M.; Schulz, S.; Maecke, H.R. Design, synthesis, and biological evaluation of somatostatin-based radiopeptides. Chem. Biol. 2006, 13, 1081–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogsitter, C.; Hartmann, H.; Wunderlich, G.; Schottelius, M.; Wester, H.J.; Kotzerke, J. Twins in spirit part IV-[(177)Lu] high affinity DOTATATE. A promising new tracer for peptide receptor radiotherapy? Nuklearmedizin 2017, 56, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Taelman, V.F.; Radojewski, P.; Marincek, N.; Ben-Shlomo, A.; Grotzky, A.; Olariu, C.I.; Perren, A.; Stettler, C.; Krause, T.; Meier, L.P.; et al. Upregulation of Key Molecules for Targeted Imaging and Therapy. J. Nucl. Med. 2016, 57, 1805–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veenstra, M.J.; van Koetsveld, P.M.; Dogan, F.; Farrell, W.E.; Feelders, R.A.; Lamberts, S.W.J.; de Herder, W.W.; Vitale, G.; Hofland, L.J. Epidrug-induced upregulation of functional somatostatin type 2 receptors in human pancreatic neuroendocrine tumor cells. Oncotarget 2018, 9, 14791–14802. [Google Scholar] [CrossRef] [Green Version]
- Venturelli, S.; Berger, A.; Weiland, T.; Essmann, F.; Waibel, M.; Nuebling, T.; Hacker, S.; Schenk, M.; Schulze-Osthoff, K.; Salih, H.R.; et al. Differential induction of apoptosis and senescence by the DNA methyltransferase inhibitors 5-azacytidine and 5-aza-2′-deoxycytidine in solid tumor cells. Mol. Cancer Ther. 2013, 12, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Stresemann, C.; Lyko, F. Modes of action of the DNA methyltransferase inhibitors azacytidine and decitabine. Int. J. Cancer 2008, 123, 8–13. [Google Scholar] [CrossRef]
- Griffiths, E.A.; Gore, S.D. Epigenetic therapies in MDS and AML. Adv. Exp. Med. Biol. 2013, 754, 253–283. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Velena, A.; Chen, B.; Petukhov, P.A.; Kozikowski, A.P.; Dritschilo, A. Novel HDAC inhibitors with radiosensitizing properties. Radiat. Res. 2005, 163, 488–493. [Google Scholar] [CrossRef]
- Hosein, A.N.; Lim, Y.C.; Day, B.; Stringer, B.; Rose, S.; Head, R.; Cosgrove, L.; Sminia, P.; Fay, M.; Martin, J.H. The effect of valproic acid in combination with irradiation and temozolomide on primary human glioblastoma cells. J. Neurooncol. 2015, 122, 263–271. [Google Scholar] [CrossRef]
- Klomp, M.J.; Dalm, S.U.; de Jong, M.; Feelders, R.A.; Hofland, J.; Hofland, L.J. Epigenetic regulation of somatostatin and somatostatin receptors in neuroendocrine tumors and other types of cancer. Rev. Endocr. Metab. Disord. 2021, 22, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Mileva, M.; Wimana, Z.; Flamen, P.; Karfis, I. Everolimus-induced somatostatin receptor overexpression in a rectal neuroendocrine tumor patient may promote somatostatin receptor-guided radionuclide therapy (peptide receptor radiotherapy) as an additional treatment option. World J. Nucl. Med. 2021, 20, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.G.; Merlin, M.A.; Adant, S.; Zine-Eddine, F.; Beauregard, J.-M.; Shah, G.M. Chemotherapy-Induced Upregulation of Somatostatin Receptor-2 Increases the Uptake and Efficacy of 177Lu-DOTA-Octreotate in Neuroendocrine Tumor Cells. Cancers 2021, 13, 232. Available online: https://0-www-mdpi-com.brum.beds.ac.uk/2072-6694/13/2/232 (accessed on 6 January 2021). [CrossRef] [PubMed]
- Adant, S.; Shah, G.M.; Beauregard, J.M. Combination treatments to enhance peptide receptor radionuclide therapy of neuroendocrine tumours. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 907–921. [Google Scholar] [CrossRef]
- Cherk, M.H.; Kong, G.; Hicks, R.J.; Hofman, M.S. Changes in biodistribution on (68)Ga-DOTA-Octreotate PET/CT after long acting somatostatin analogue therapy in neuroendocrine tumour patients may result in pseudoprogression. Cancer Imaging 2018, 18, 3. [Google Scholar] [CrossRef] [Green Version]
- Shimura, H.; Haraguchi, K.; Miyazaki, A.; Endo, T.; Onaya, T. Iodide Uptake and Experimental 131I Therapy in Transplanted Undifferentiated Thyroid Cancer Cells Expressing the Na+/I− Symporter Gene. Endocrinology 1997, 138, 4493–4496. [Google Scholar] [CrossRef]
- Haberkorn, U. Gene therapy with sodium/iodide symporter in hepatocarcinoma. Exp. Clin. Endocrinol. Diabetes 2001, 109, 60–62. [Google Scholar] [CrossRef]
- Haberkorn, U.; Kinscherf, R.; Kissel, M.; Kubler, W.; Mahmut, M.; Sieger, S.; Eisenhut, M.; Peschke, P.; Altmann, A. Enhanced iodide transport after transfer of the human sodium iodide symporter gene is associated with lack of retention and low absorbed dose. Gene Ther. 2003, 10, 774–780. [Google Scholar] [CrossRef]
- Baril, P.; Martin-Duque, P.; Vassaux, G. Visualization of gene expression in the live subject using the Na/I symporter as a reporter gene: Applications in biotherapy. Br. J. Pharmacol. 2010, 159, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Bruno, J.F.; Xu, Y.; Song, J.; Berelowitz, M. Molecular cloning and functional expression of a brain-specific somatostatin receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 11151–11155. [Google Scholar] [CrossRef] [Green Version]
- O’Carroll, A.M.; Lolait, S.J.; König, M.; Mahan, L.C. Molecular cloning and expression of a pituitary somatostatin receptor with preferential affinity for somatostatin-28. Mol. Pharmacol. 1992, 42, 939–946. [Google Scholar] [PubMed]
- Rohrer, L.; Raulf, F.; Bruns, C.; Buettner, R.; Hofstaedter, F.; Schüle, R. Cloning and characterization of a fourth human somatostatin receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 4196–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidari, P.; Kunawudhi, A.; Martinez-Quintanilla, J.; Szretter, A.; Shah, K.; Mahmood, U. Somatostatin receptor type 2 as a radiotheranostic PET reporter gene for oncologic interventions. Theranostics 2018, 8, 3380–3391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Moroz, M.A.; Serganova, I.; Ku, T.; Huang, R.; Vider, J.; Maecke, H.R.; Larson, S.M.; Blasberg, R.; Smith-Jones, P.M. Imaging expression of the human somatostatin receptor subtype-2 reporter gene with 68Ga-DOTATOC. J. Nucl. Med. 2011, 52, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kliewer, A.; Mann, A.; Petrich, A.; Poll, F.; Schulz, S. A transplantable phosphorylation probe for direct assessment of G protein-coupled receptor activation. PLoS ONE 2012, 7, e39458. [Google Scholar] [CrossRef] [PubMed]
- Petrich, A.; Mann, A.; Kliewer, A.; Nagel, F.; Strigli, A.; Martens, J.C.; Poll, F.; Schulz, S. Phosphorylation of threonine 333 regulates trafficking of the human sst5 somatostatin receptor. Mol. Endocrinol. 2013, 27, 671–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poll, F.; Lehmann, D.; Illing, S.; Ginj, M.; Jacobs, S.; Lupp, A.; Stumm, R.; Schulz, S. Pasireotide and octreotide stimulate distinct patterns of sst2A somatostatin receptor phosphorylation. Mol. Endocrinol. 2010, 24, 436–446. [Google Scholar] [CrossRef] [Green Version]
- Pulukuri, S.M.; Gondi, C.S.; Lakka, S.S.; Jutla, A.; Estes, N.; Gujrati, M.; Rao, J.S. RNA interference-directed knockdown of urokinase plasminogen activator and urokinase plasminogen activator receptor inhibits prostate cancer cell invasion, survival, and tumorigenicity in vivo. J. Biol. Chem. 2005, 280, 36529–36540. [Google Scholar] [CrossRef] [Green Version]
- Freudenberg, R.; Runge, R.; Maucksch, U.; Berger, V.; Kotzerke, J. On the dose calculation at the cellular level and its implications for the RBE of (99m)Tc and (1)(2)(3)I. Med. Phys. 2014, 41, 062503. [Google Scholar] [CrossRef]
- Maucksch, U.; Runge, R.; Wunderlich, G.; Freudenberg, R.; Naumann, A.; Kotzerke, J. Comparison of the radiotoxicity of the (99m)Tc-labeled compounds (99m)Tc-pertechnetate, (99m)Tc-HMPAO and (99m)Tc-MIBI. Int. J. Radiat. Biol. 2016, 92, 698–706. [Google Scholar] [CrossRef]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Klomp, M.J.; Dalm, S.U.; van Koetsveld, P.M.; Dogan, F.; de Jong, M.; Hofland, L.J. Comparing the Effect of Multiple Histone Deacetylase Inhibitors on SSTR2 Expression and [(111)In]In-DOTATATE Uptake in NET Cells. Cancers 2021, 13, 4905. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, P.; Gűnther, T.; Schulz, S. Pharmacological Characterization of Veldoreotide as a Somatostatin Receptor 4 Agonist. Life 2021, 11, 1075. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Marquez, M.; Nilsson, S.; Holmberg, A.R. Incubation with somatostatin, 5-aza decitabine and trichostatin up-regulates somatostatin receptor expression in prostate cancer cells. Oncol. Rep. 2008, 20, 151–154. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/18575731 (accessed on 1 July 2008). [CrossRef] [PubMed] [Green Version]
- Gailhouste, L.; Liew, L.C.; Hatada, I.; Nakagama, H.; Ochiya, T. Epigenetic reprogramming using 5-azacytidine promotes an anti-cancer response in pancreatic adenocarcinoma cells. Cell Death Dis. 2018, 9, 468. [Google Scholar] [CrossRef] [Green Version]
- Gailhouste, L.; Liew, L.C.; Yasukawa, K.; Hatada, I.; Tanaka, Y.; Nakagama, H.; Ochiya, T. Differentiation Therapy by Epigenetic Reconditioning Exerts Antitumor Effects on Liver Cancer Cells. Mol. Ther. 2018, 26, 1840–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrisani, J.; Hanoun, N.; Laurell, H.; Lopez, F.; Maoret, J.J.; Souque, A.; Susini, C.; Cordelier, P.; Buscail, L. Identification of an upstream promoter of the human somatostatin receptor, hSSTR2, which is controlled by epigenetic modifications. Endocrinology 2008, 149, 3137–3147. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Liu, D.; Lee, D.; Kapoor, S.; Gibson-Corley, K.N.; Quinn, T.P.; Sagastume, E.A.; Mott, S.L.; Walsh, S.A.; Acevedo, M.R.; et al. Enhancing the Efficacy of Melanocortin 1 Receptor-Targeted Radiotherapy by Pharmacologically Upregulating the Receptor in Metastatic Melanoma. Mol. Pharm. 2019, 16, 3904–3915. [Google Scholar] [CrossRef]
- Noss, K.R.; Wolfe, S.A.; Grimes, S.R. Upregulation of prostate specific membrane antigen/folate hydrolase transcription by an enhancer. Gene 2002, 285, 247–256. [Google Scholar] [CrossRef]
- Chie, E.K.; Shin, J.H.; Kim, J.H.; Kim, H.J.; Kim, I.A.; Kim, I.H. In Vitro and In Vivo Radiosensitizing Effect of Valproic Acid on Fractionated Irradiation. Cancer Res. Treat. 2015, 47, 527–533. [Google Scholar] [CrossRef]
- Jin, X.F.; Auernhammer, C.J.; Ilhan, H.; Lindner, S.; Nolting, S.; Maurer, J.; Spottl, G.; Orth, M. Combination of 5-Fluorouracil with Epigenetic Modifiers Induces Radiosensitization, Somatostatin Receptor 2 Expression, and Radioligand Binding in Neuroendocrine Tumor Cells In Vitro. J. Nucl. Med. 2019, 60, 1240–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hontecillas-Prieto, L.; Flores-Campos, R.; Silver, A.; de Álava, E.; Hajji, N.; García-Domínguez, D.J. Synergistic Enhancement of Cancer Therapy Using HDAC Inhibitors: Opportunity for Clinical Trials. Front. Genet. 2020, 11, 1113. [Google Scholar] [CrossRef] [PubMed]
- DuBois, S.G.; Granger, M.M.; Groshen, S.; Tsao-Wei, D.; Ji, L.; Shamirian, A.; Czarnecki, S.; Goodarzian, F.; Berkovich, R.; Shimada, H.; et al. Randomized Phase II Trial of MIBG Versus MIBG, Vincristine, and Irinotecan Versus MIBG and Vorinostat for Patients With Relapsed or Refractory Neuroblastoma: A Report From NANT Consortium. J. Clin. Oncol. 2021, 39, 3506–3514. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ma, W.; Wang, J.; Deng, J.; Yuan, M.; Zhao, M.; Jia, L.; Yang, W.; Zhang, R.; Qin, W.; et al. Imaging and therapy of hSSTR2-transfected tumors using radiolabeled somatostatin analogs. Tumour Biol. 2013, 34, 2451–2457. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, H. Somatostatin receptor based imaging and radionuclide therapy. BioMed Res. Int. 2015, 2015, 917968. [Google Scholar] [CrossRef]
- Zhao, R.; Yang, W.; Wang, Z.; Li, G.; Qin, W.; Wang, J. Treatment of transplanted tumor of lung adenocarcinoma A549 transfected by human somatostatin receptor subtype 2 (hsstr2) gene with 188Re-RC-160. Nucl. Med. Biol. 2010, 37, 977–987. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Conditions | IC20/D80/A80 | IC40/D60/A60 |
|---|---|---|
| 5-aza-dC (µM) | 0.25 | 0.46 |
| VPA (mM) | 0.6 | 1.0 |
| X-ray (Gy) | 0.6 | 1.2 |
| Lu-177-DOTATATE (kBq/mL) | 57.5 | 136 |
| Concentration 5-aza-dC (µM) | |||||||||
| Survival Fraction (%) ± SD | 0 | 0.01 | 0.05 | 0.1 | 0.5 | 1.0 | 5.0 | 10 | 25 |
| HEKsst2 | 100 ± 6.2 | 97.9 ± 1.8 | 118.0 ± 31 | 83.2 ± 4.8 | 96.6 ± 13.2 | 93.7 ± 1.3 | 42.3 ± 4.8 | 18.6 ± 0.8 | 1.7 ± 0.4 |
| PC3 | 100 ± 3.6 | 67.1 ± 3.3 | 52.1 ± 19.0 | 69.4 ± 2.1 | 41.6 ± 1.8 | 24.9 ± 2.0 | 27.5 ± 3.2 | 9.4 ± 0.3 | 1.9 ± 0.02 |
| Concentration VPA (mM) | |||||||||
| Survival fraction (%) ± SD | 0 | 0.1 | 0.5 | 1.0 | 2.5 | 5.0 | 10 | 25 | 50 |
| HEKsst2 | 100 ± 10.7 | 183.1 ± 7.2 | 78.7 ± 10.8 | 67.5 ± 15.9 | 18.5 ± 2.2 | 21.0 ± 5.1 | 53.5 ± 5.8 | 8.0 ± 0.8 | n.a. 1 |
| PC3 | 100 ± 8.1 | 95.6 ± 4.5 | 95.6 ± 2.3 | 72.6 ± 11.6 | 69.4 ± 6.0 | 35.1 ± 4.6 | 24.0 ± 1.2 | 1.2 ± 1.1 | 1.7 ± 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotzerke, J.; Buesser, D.; Naumann, A.; Runge, R.; Huebinger, L.; Kliewer, A.; Freudenberg, R.; Brogsitter, C. Epigenetic-Like Stimulation of Receptor Expression in SSTR2 Transfected HEK293 Cells as a New Therapeutic Strategy. Cancers 2022, 14, 2513. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102513
Kotzerke J, Buesser D, Naumann A, Runge R, Huebinger L, Kliewer A, Freudenberg R, Brogsitter C. Epigenetic-Like Stimulation of Receptor Expression in SSTR2 Transfected HEK293 Cells as a New Therapeutic Strategy. Cancers. 2022; 14(10):2513. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102513
Chicago/Turabian StyleKotzerke, Joerg, Dorothee Buesser, Anne Naumann, Roswitha Runge, Lisa Huebinger, Andrea Kliewer, Robert Freudenberg, and Claudia Brogsitter. 2022. "Epigenetic-Like Stimulation of Receptor Expression in SSTR2 Transfected HEK293 Cells as a New Therapeutic Strategy" Cancers 14, no. 10: 2513. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102513