
Influence of Malignant Pleural Fluid from Lung Adenocarcinoma Patients on Neutrophil Response

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Pleural Fluid Samples
2.2. Phenotyping Neutrophils from Pleural Effusions by Flow Cytometry
2.3. Isolation of Neutrophils from Healthy Donors
2.4. Induction and Quantification of NETosis in the Presence of Cell-Free Pleural Fluids
2.5. Measurement of Oxidative Burst in the Presence of Cell-Free Pleural Fluids
2.6. Induction of Neutrophil Apoptosis in the Presence of Cell-Free Pleural Fluids
2.7. Determination of Biochemical Parameters and Soluble Mediators in Pleural Fluids
2.8. Statistics Analysis
3. Results
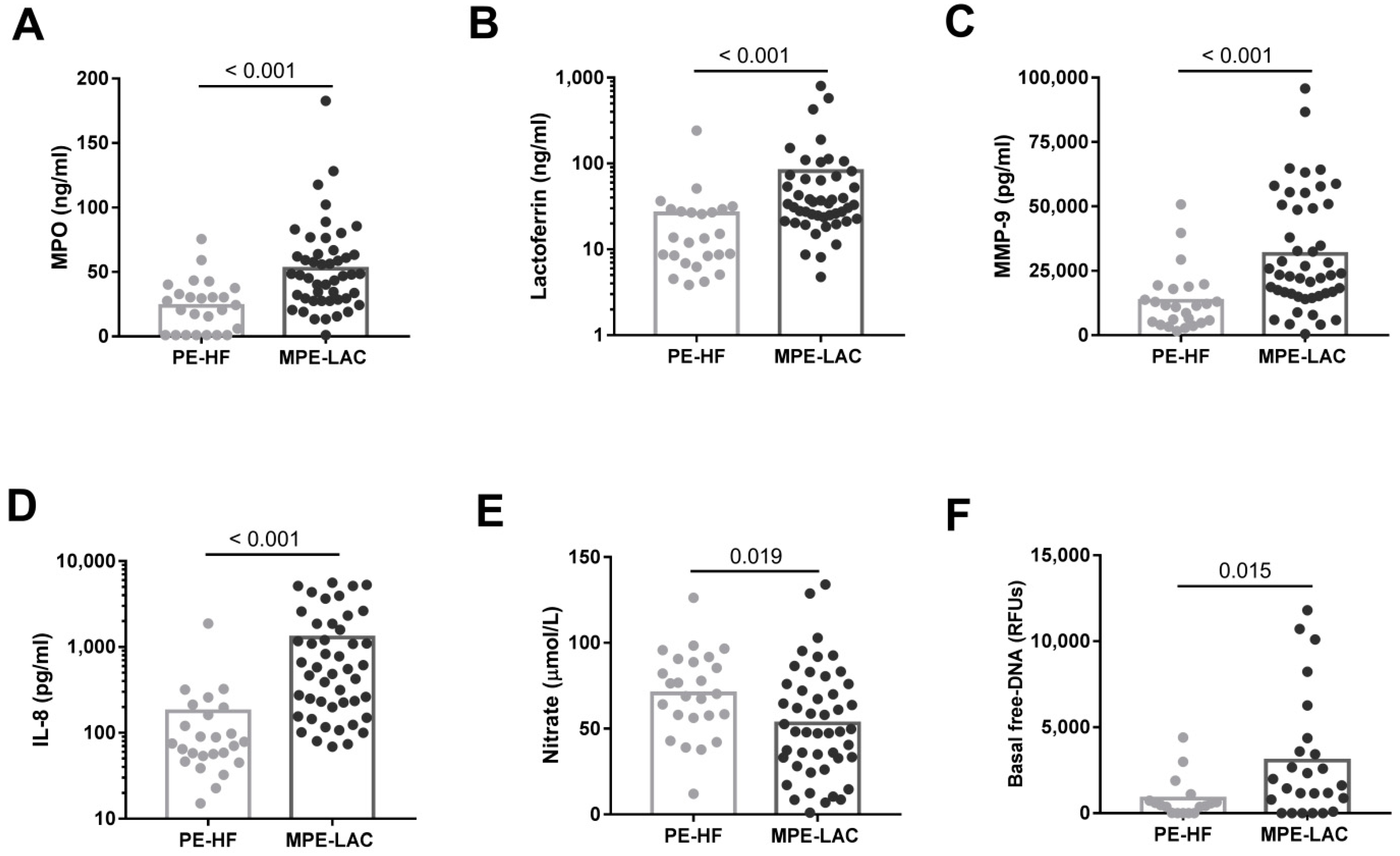
3.1. Phenotype of Pleural Neutrophils and Concentration of Neutrophil-Associated Molecules in Pleural Fluids
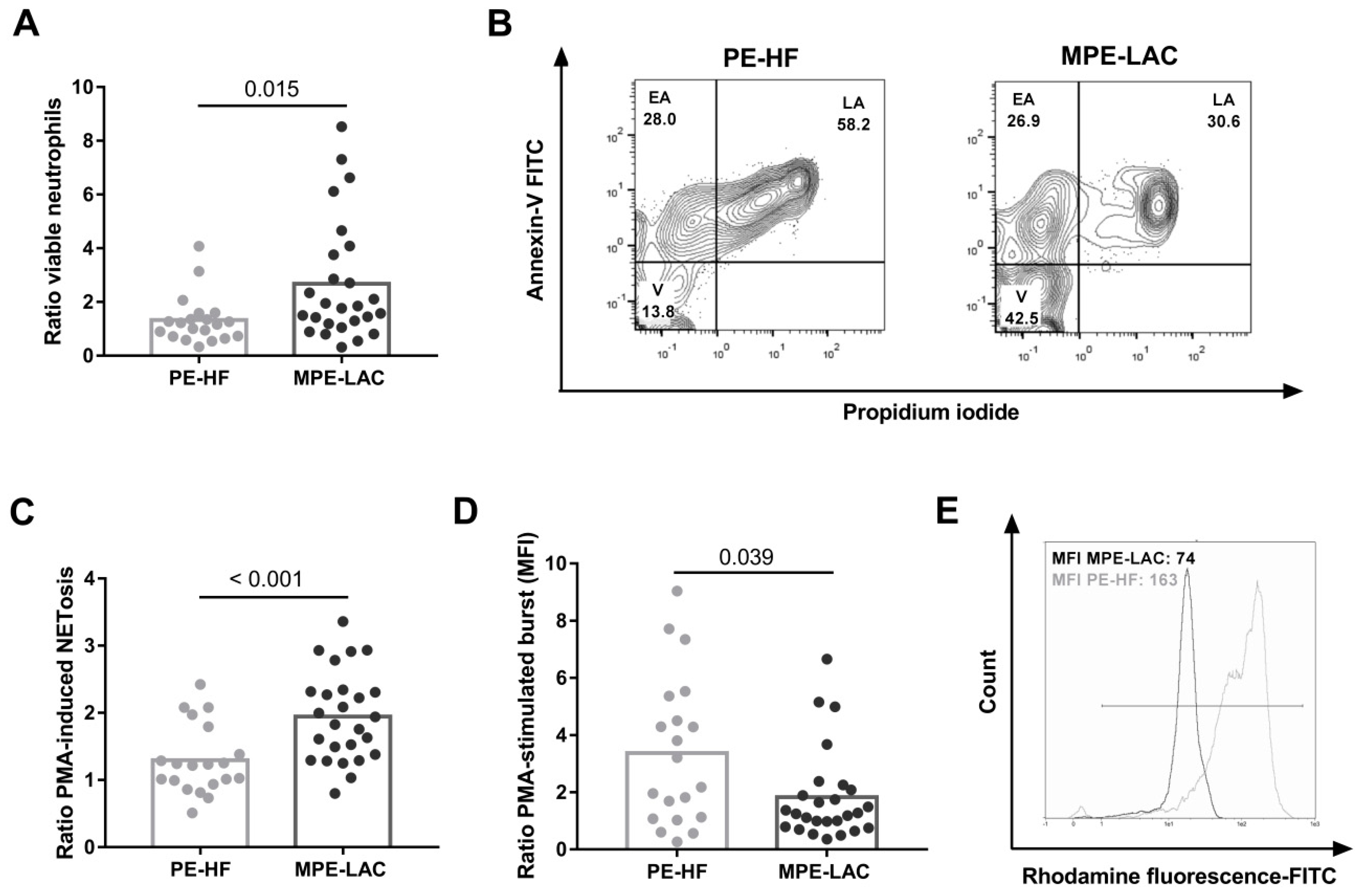
3.2. Effect of Cell-Free Pleural Fluid on Neutrophil Viability and Function
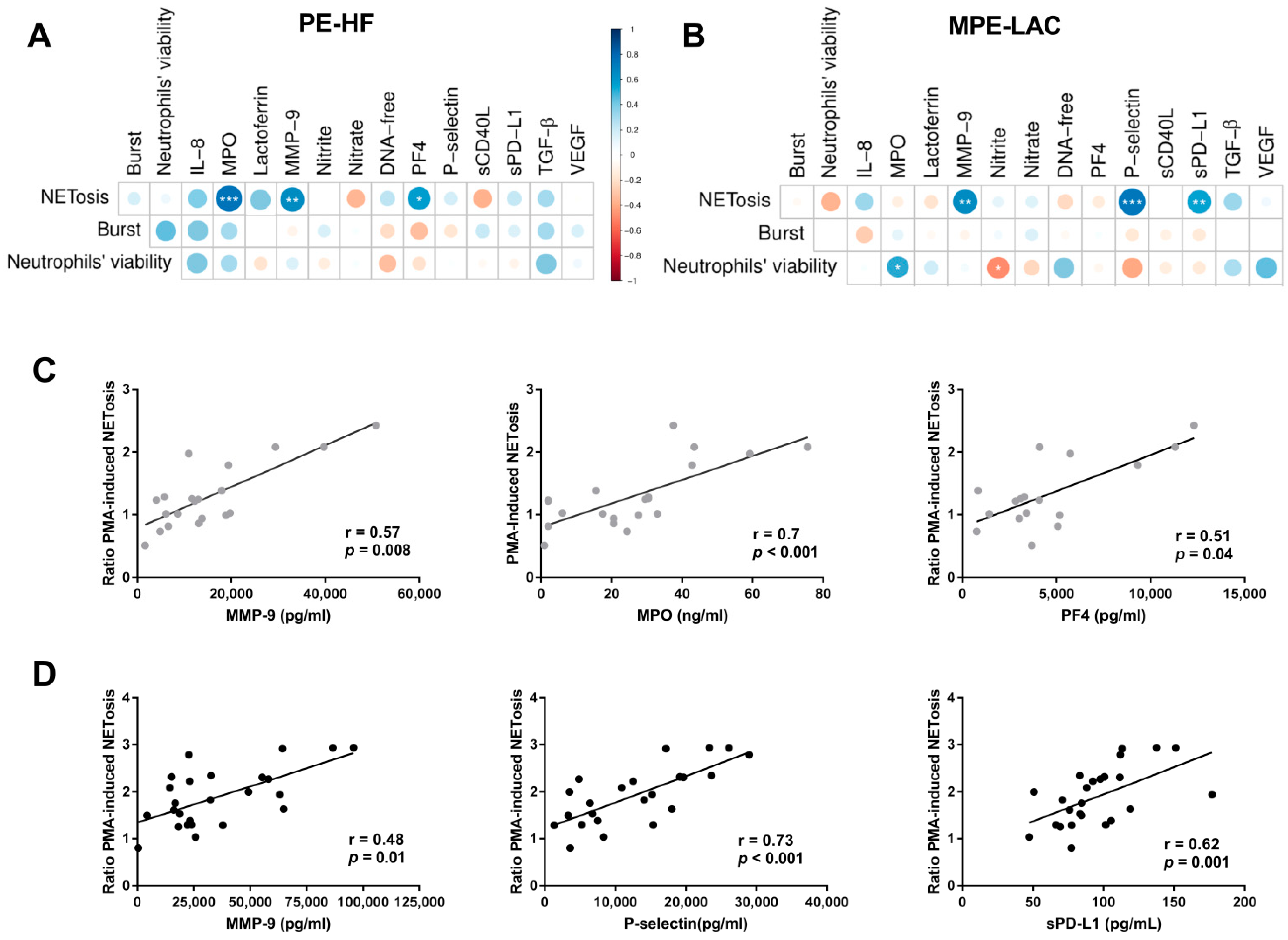
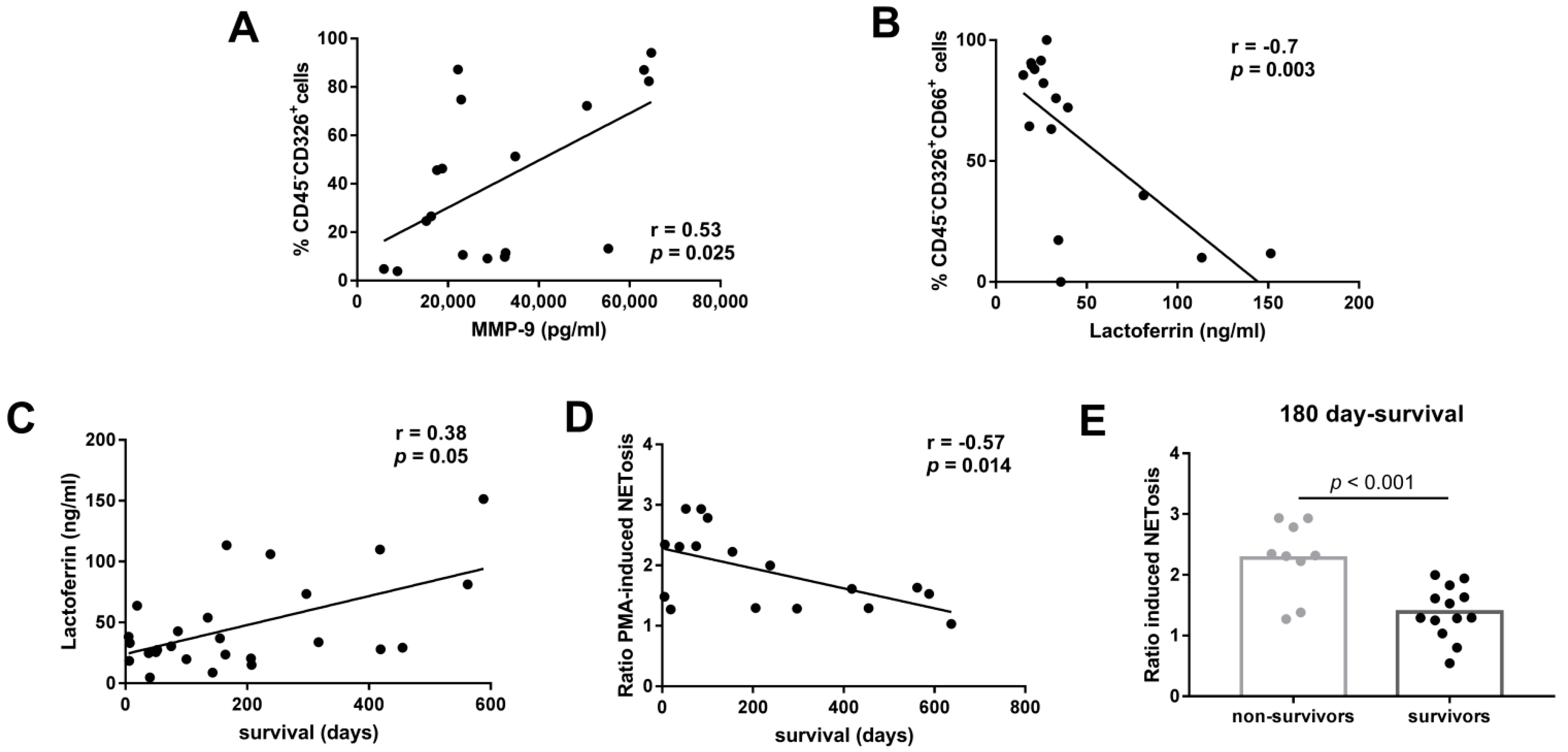
3.3. Association between Immunomodulatory Factors in Pleural Fluid and Neutrophil Function
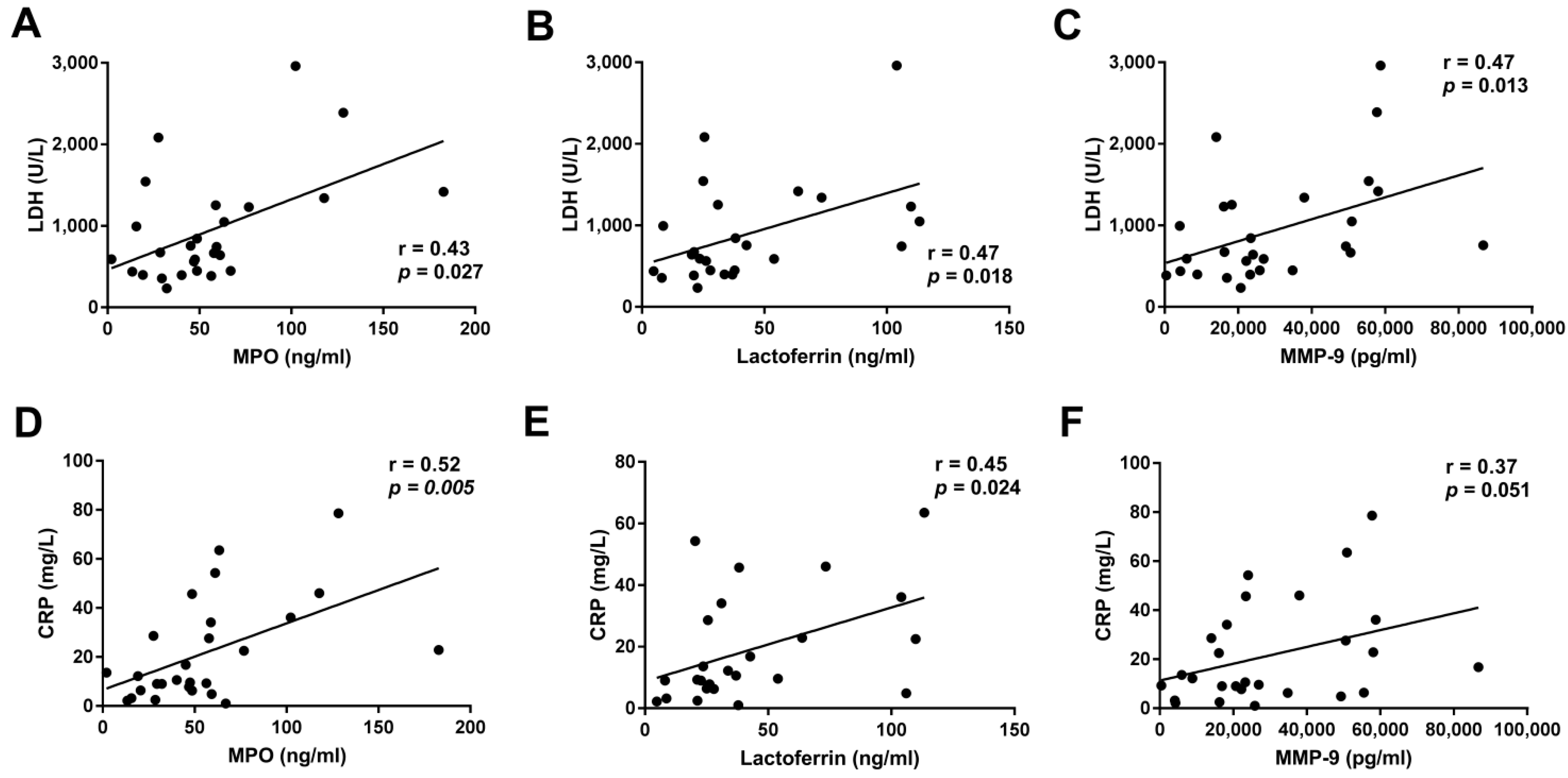
3.4. Association between Neutrophil Functions and the Clinical Outcome of LAC Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Psallidas, I.; Kalomenidis, I.; Porcel, J.M.; Robinson, B.W.; Stathopoulos, G.T. Malignant Pleural Effusion: From Bench to Bedside. Eur. Respir. Rev. 2016, 25, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Porcel, J.M.; Light, R.W. Pleural Effusions. Disease-a-Month 2013, 59, 29–57. [Google Scholar] [CrossRef] [PubMed]
- Hooper, C.; Lee, Y.C.G.; Maskell, N. Investigation of a Unilateral Pleural Effusion in Adults: British Thoracic Society Pleural Disease Guideline 2010. Thorax 2010, 65, ii4–ii17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Nam, H.S.; Lim, J.H.; Kim, J.S.; Moon, Y.; Cho, J.H.; Ryu, J.S.; Kwak, S.M.; Lee, H.L. Prognostic Impact of a New Score Using Neutrophil-to-Lymphocyte Ratios in the Serum and Malignant Pleural Effusion in Lung Cancer Patients. BMC Cancer 2017, 17, 557. [Google Scholar] [CrossRef]
- Nieto, J.C.; Zamora, C.; Porcel, J.M.; Mulet, M.; Pajares, V.; Muñoz-Fernandez, A.M.; Calvo, N.; Espinosa, I.; Pascual-García, M.; Bielsa, S.; et al. Migrated T Lymphocytes into Malignant Pleural Effusions: An Indicator of Good Prognosis in Lung Adenocarcinoma Patients. Sci. Rep. 2019, 9, 2996. [Google Scholar] [CrossRef]
- Broaddus, V.C.; Hébert, C.A.; Vitangcol, R.V.; Hoeffel, J.M.; Bernstein, M.S.; Boylan, A.M. Interleukin-8 is a major neutrophil chemotactic factor in pleural liquid of patients with empyema. Am. Rev. Respir. Dis. 1992, 146, 825. [Google Scholar] [CrossRef]
- Baǧci Ceyhan, B.; Özgün, S.; Çelikel, T.; Yalçin, M.; Koç, M. IL-8 in Pleural Effusion. Respir. Med. 1996, 90, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Pace, E.; Profita, M.; Melis, M.; Bonanno, A.; Paternò, A.; Mody, C.H.; Spatafora, M.; Ferraro, M.; Siena, L.; Vignola, A.M.; et al. LTB4 Is Present in Exudative Pleural Effusions and Contributes Actively to Neutrophil Recruitment in the Inflamed Pleural Space. Clin. Exp. Immunol. 2004, 135, 519–527. [Google Scholar] [CrossRef]
- Liu, G.N.; Shi, H.Z.; Xie, Z.H.; Shen, H.H.; Huang, H.Q.; Deng, J.M.; Liang, Q.L.; Wu, Y.B. Epithelial Neutrophil-Activating Peptide-78 Recruits Neutrophils into Pleural Effusion. Eur. Respir. J. 2009, 34, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Furumaya, C.; Martinez-Sanz, P.; Bouti, P.; Kuijpers, T.W.; Matlung, H.L. Plasticity in Pro- and Anti-Tumor Activity of Neutrophils: Shifting the Balance. Front. Immunol. 2020, 11, 2100. [Google Scholar] [CrossRef]
- Singel, K.L.; Emmons, T.R.; Khan, A.N.H.; Mayor, P.C.; Shen, S.; Wong, J.T.; Morrell, K.; Eng, K.H.; Mark, J.; Bankert, R.B.; et al. Mature Neutrophils Suppress T Cell Immunity in Ovarian Cancer Microenvironment. JCI Insight 2019, 4, e122311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masucci, M.T.; Minopoli, M.; Carriero, M.V. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front. Oncol. 2019, 9, 1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houghton, A.M.G.; Rzymkiewicz, D.M.; Ji, H.; Gregory, A.D.; Egea, E.E.; Metz, H.E.; Stolz, D.B.; Land, S.R.; Marconcini, L.A.; Kliment, C.R.; et al. Neutrophil Elastase-Mediated Degradation of IRS-1 Accelerates Lung Tumor Growth. Nat. Med. 2010, 16, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Merchant, N.; Nagaraju, G.P.; Rajitha, B.; Lammata, S.; Jella, K.K.; Buchwald, Z.S.; Lakka, S.S.; Ali, A.N. Matrix Metalloproteinases: Their Functional Role in Lung Cancer. Carcinogenesis 2017, 38, 766–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, A.K.; Cedervall, J. NETosis in Cancer—Platelet-Neutrophil Crosstalk Promotes Tumor-Associated Pathology. Front. Immunol. 2016, 7, 373. [Google Scholar] [CrossRef] [Green Version]
- Keshari, R.S.; Jyoti, A.; Dubey, M.; Kothari, N.; Kohli, M.; Bogra, J.; Barthwal, M.K.; Dikshit, M. Cytokines Induced Neutrophil Extracellular Traps Formation: Implication for the Inflammatory Disease Condition. PLoS ONE 2012, 7, e48111. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, O.E.; Borregaard, N. Neutrophil Extracellular Traps—The Dark Side of Neutrophils. J. Clin. Investig. 2016, 126, 1612–1620. [Google Scholar] [CrossRef] [Green Version]
- Ireland, A.S.; Oliver, T.G. Neutrophils Create an ImpeNETrable Shield between Tumor and Cytotoxic Immune Cells. Immunity 2020, 52, 729–731. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil Extracellular Traps Produced during Inflammation Awaken Dormant Cancer Cells in Mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef] [Green Version]
- Knaapen, A.M.; Güngör, N.; Schins, R.P.F.; Borm, P.J.A.; Van Schooten, F.J. Neutrophils and Respiratory Tract DNA Damage and Mutagenesis: A Review. Mutagenesis 2006, 21, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-Associated Macrophages as Treatment Targets in Oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Nieto, J.C.; Perea, L.; Soriano, G.; Zamora, C.; Cantó, E.; Medina, A.; Poca, M.; Sanchez, E.; Roman, E.; Julià, G.; et al. Ascitic Fluid Regulates the Local Innate Immune Response of Patients with Cirrhosis. J. Leukoc. Biol. 2018, 104, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, e24. [Google Scholar]
- Warner, A.; Dahele, M.; Hu, B.; Palma, D.A.; Senan, S.; Oberije, C.; Tsujino, K.; Moreno-Jimenez, M.; Kim, T.H.; Marks, L.B.; et al. Factors Associated with Early Mortality in Patients Treated with Concurrent Chemoradiation Therapy for Locally Advanced Non-Small Cell Lung Cancer. Int. J. Radiat. Oncol. Biol. Phys. 2016, 94, 612–620. [Google Scholar] [CrossRef]
- Mulet, M.; Zamora, C.; Porcel, J.M.; Nieto, J.C.; Pajares, V.; Muñoz-Fernandez, A.M.; Calvo, N.; Esquerda, A.; Vidal, S. Platelet Factor 4 Regulates T Cell Effector Functions in Malignant Pleural Effusions. Cancer Lett. 2020, 491, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Segura, R.M.; Alegre, J.; Varela, E.; Marti, R.; Suriñach, J.M.; Jufresa, J.; Armadans, L.; Pascual, C.; De Sevilla, T.F. Interleukin-8 and Markers of Neutrophil Degranulation in Pleural Effusions. Am. J. Respir. Crit. Care Med. 1998, 157, 1565–1572. [Google Scholar] [CrossRef]
- Park, K.J.; Hwang, S.C.; Sheen, S.S.; Oh, Y.J.; Han, J.H.; Lee, K.B. Expression of Matrix Metalloproteinase-9 in Pleural Effusions of Tuberculosis and Lung Cancer. Respiration 2005, 72, 166–175. [Google Scholar] [CrossRef]
- Hegnhoj, J.; Galloee, A. Diagnostic Value of Lactoferrin Analysis in Pleural Effusions. Clin. Chem. Lab. Med. 1994, 32, 521–524. [Google Scholar] [CrossRef]
- Pace, E.; Gjomarkaj, M.; Melis, M.; Profita, M.; Spatafora, M.; Vignola, A.M.; Bonsignore, G.; Mody, C.H. Interleukin-8 Induces Lymphocyte Chemotaxis into the Pleural Space Role of Pleural Macrophages. Am. J. Respir. Crit. Care Med. 1999, 159, 1592–1599. [Google Scholar] [CrossRef] [Green Version]
- Brostjan, C.; Oehler, R. The Role of Neutrophil Death in Chronic Inflammation and Cancer. Cell Death Discov. 2020, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- El Kebir, D.; József, L.; Pan, W.; Filep, J.G. Myeloperoxidase Delays Neutrophil Apoptosis through CD11b/CD18 Integrins and Prolongs Inflammation. Circ. Res. 2008, 103, 352–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andonegui, G.; Trevani, A.S.; López, D.H.; Raiden, S.; Giordano, M.; Geffner, J.R. Inhibition of Human Neutrophil Apoptosis by Platelets. J. Immunol. 1997, 158, 3372–3377. [Google Scholar] [PubMed]
- Colotta, F.; Re, F.; Polentarutti, N.; Sozzani, S.; Mantovani, A. Modulation of Granulocyte Survival and Programmed Cell Death by Cytokines and Bacterial Products. Blood 1992, 80, 2012–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, A.G.; Bastos, V.A.F.; Fujimura, P.T.; Ferreira, I.C.C.; Leal, L.F.; da Silva, L.S.; Laus, A.C.; Reis, R.M.; Martins, M.M.; Santos, P.S.; et al. Cell-Free DNA Promotes Malignant Transformation in Non-Tumor Cells. Sci. Rep. 2020, 10, 21674. [Google Scholar] [CrossRef] [PubMed]
- Twaddell, S.H.; Gibson, P.G.; Grainge, C.; Baines, K.J. Parapneumonic Effusions Are Characterized by Elevated Levels of Neutrophil Extracellular Traps. Chest 2021, 160, 1645–1655. [Google Scholar] [CrossRef]
- Carmona-Rivera, C.; Zhao, W.; Yalavarthi, S.; Kaplan, M.J. Neutrophil Extracellular Traps Induce Endothelial Dysfunction in Systemic Lupus Erythematosus through the Activation of Matrix Metalloproteinase-2. Ann. Rheum. Dis. 2015, 74, 1417–1424. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Han, L.; Qiu, X.; Wang, G.; Zheng, J. Neutrophil Extracellular Traps in Digestive Cancers: Warrior or Accomplice. Front. Oncol. 2021, 11, 4914. [Google Scholar] [CrossRef]
- Etulain, J.; Martinod, K.; Wong, S.L.; Cifuni, S.M.; Schattner, M.; Wagner, D.D. P-Selectin Promotes Neutrophil Extracellular Trap Formation in Mice. Blood 2015, 126, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Kaltenmeier, C.; Yazdani, H.O.; Morder, K.; Geller, D.A.; Simmons, R.L.; Tohme, S. Neutrophil Extracellular Traps Promote T Cell Exhaustion in the Tumor Microenvironment. Front. Immunol. 2021, 12, 785222. [Google Scholar] [CrossRef]
- Ardi, V.C.; Kupriyanova, T.A.; Deryugina, E.I.; Quigley, J.P. Human Neutrophils Uniquely Release TIMP-Free MMP-9 to Provide a Potent Catalytic Stimulator of Angiogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 20262–20267. [Google Scholar] [CrossRef] [Green Version]
- Barillari, G. The Impact of Matrix Metalloproteinase-9 on the Sequential Steps of the Metastatic Process. Int. J. Mol. Sci. 2020, 21, 4526. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Kamiya, M.; Urano, Y.; Nishi, H.; Herter, J.M.; Mayadas, T.; Hirohama, D.; Suzuki, K.; Kawakami, H.; Tanaka, M.; et al. Lactoferrin Suppresses Neutrophil Extracellular Traps Release in Inflammation. EBioMedicine 2016, 10, 204–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofoluwe, A.; Bacchetta, M.; Badaoui, M.; Kwak, B.R.; Chanson, M. ATP Amplifies NADPH-Dependent and -Independent Neutrophil Extracellular Trap Formation. Sci. Rep. 2019, 9, 16556. [Google Scholar] [CrossRef] [PubMed]
- Elbim, C.; Lizard, G. Flow Cytometric Investigation of Neutrophil Oxidative Burst and Apoptosis in Physiological and Pathological Situations. Cytom. Part A 2009, 75, 475–481. [Google Scholar] [CrossRef]
- Jiao, Y.; Li, W.; Wang, W.; Tong, X.; Xia, R.; Fan, J.; Du, J.; Zhang, C.; Shi, X. Platelet-Derived Exosomes Promote Neutrophil Extracellular Trap Formation during Septic Shock. Crit. Care 2020, 24, 380. [Google Scholar] [CrossRef]
- Leal, A.C.; Mizurini, D.M.; Gomes, T.; Rochael, N.C.; Saraiva, E.M.; Dias, M.S.; Werneck, C.C.; Sielski, M.S.; Vicente, C.P.; Monteiro, R.Q. Tumor-Derived Exosomes Induce the Formation of Neutrophil Extracellular Traps: Implications For The Establishment of Cancer-Associated Thrombosis. Sci. Rep. 2017, 7, 6438. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients Characteristics | HF | LAC | p Value |
|---|---|---|---|
| Number of patients | 25 | 47 | |
| Male/female | 17/8 | 36/11 | n.s. |
| Median age (range), years | 85 (81–88) | 69 (62–75) | <0.001 |
| Median (IQR) survival time from diagnosis, days | NA | 203.5 (69.3–527.8) | |
| EGFR mutations | NA | 11/47 | |
| ALK mutations | NA | 4/47 | |
| ECOG PS | |||
| 0 | NA | 12/47 | |
| 1 | NA | 8/47 | |
| 2 | NA | 5/47 | |
| 3 | NA | 10/47 | |
| 4 | NA | 5/47 | |
| NA | NA | 7/47 | |
| Biochemical parameters | |||
| Glucose (mg/mL) | 125.5 (102.8–154) | 99 (81.56–123.8) | 0.026 |
| Lactate dehydrogenase (U/L) | 163.5 (138.8–211.3) | 755 (449–1253) | <0.001 |
| C-reactive protein (mg/L) | 3.8 (2.3–10.9) | 11.2 (6.4–34.6) | 0.004 |
| Adenosine deaminase (U/L) | 4.3 (3.2–7.8) | 7.4 (5.1–10.5) | n.s. |
| Total protein (g/dL) | 2.6 (1.8–3) | 4.8 (4–5.7) | <0.001 |
| pH | 7.5 ± 0.02 | 7.4 ± 0.02 | 0.001 |
| Treatment | |||
| Radiotherapy | NA | 5/47 | |
| Chemotherapy | NA | 17/47 | |
| Immunotherapy | NA | 10/47 | |
| ALK inhibitors | NA | 2/47 | |
| EGFR inhibitors | NA | 5/47 | |
| Her2 inhibitors | NA | 2/47 | |
| Untreated or palliative care | NA | 6/47 | |
| Soluble platelet- related molecules in pleural fluids | |||
| PF4 (pg/mL) | 3281 (2125–4593) | 8282 (4107–14,605) | <0.001 |
| VEGF (ng/mL) | 119.3 (1–455.3) | 2377 (313.3–3598) | 0.001 |
| P-selectin (pg/mL) | 4440 ± 323.7 | 11,484 ± 1260 | 0.001 |
| TGF-β (pg/mL) | 11,572 (9320–15,950) | 16,838 (8662–28,772) | 0.04 |
| CD40L (pg/mL) | 2.1 (1–56.2) | 26.2 (2.1–157.1) | 0.07 |
| PD-L1 (pg/mL) | 40.3 (32.5–73.6) | 96.4 (76.4–132.6) | <0.001 |
| PE-HF | MPE-LAC | p Value | |
|---|---|---|---|
| Neutrophils (cell/mL) | 24,000 (4967–106,120) | 21,170 (1491–140,580) | n.s.1 |
| Neutrophil phenotype | |||
| CD15 expression (MFI) | 48.6 ± 6.5 | 55.9 ± 10.9 | n.s. 2 |
| CD16 expression (MFI) | 21.8 ± 3.8 | 26.2 ± 3.6 | n.s. 2 |
| CD66b expression (MFI) | 19.2 (14.2–25.3) | 22.9 (14.7–26.8) | n.s. 1 |
| S100A8/9+ neutrophils (%) | 48.5 ± 7.4 | 31.4 ± 3.8 | 0.09 2 |
| CD200R+ neutrophils (%) | 5.1 ± 1.9 | 4.3 ± 0.6 | n.s. 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulet, M.; Osuna-Gómez, R.; Zamora, C.; Porcel, J.M.; Nieto, J.C.; Perea, L.; Pajares, V.; Muñoz-Fernandez, A.M.; Calvo, N.; Sorolla, M.A.; et al. Influence of Malignant Pleural Fluid from Lung Adenocarcinoma Patients on Neutrophil Response. Cancers 2022, 14, 2529. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102529
Mulet M, Osuna-Gómez R, Zamora C, Porcel JM, Nieto JC, Perea L, Pajares V, Muñoz-Fernandez AM, Calvo N, Sorolla MA, et al. Influence of Malignant Pleural Fluid from Lung Adenocarcinoma Patients on Neutrophil Response. Cancers. 2022; 14(10):2529. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102529
Chicago/Turabian StyleMulet, Maria, Rubén Osuna-Gómez, Carlos Zamora, José M. Porcel, Juan C. Nieto, Lídia Perea, Virginia Pajares, Ana M. Muñoz-Fernandez, Nuria Calvo, Maria Alba Sorolla, and et al. 2022. "Influence of Malignant Pleural Fluid from Lung Adenocarcinoma Patients on Neutrophil Response" Cancers 14, no. 10: 2529. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102529