
Mutational Activation of the NRF2 Pathway Upregulates Kynureninase Resulting in Tumor Immunosuppression and Poor Outcome in Lung Adenocarcinoma

 , , , , , ,
, , , , , ,  add
Show full author list
add
Show full author list
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transfection
2.2. Chemicals
2.3. Western Blot Analysis
2.4. RT-PCR Analysis
2.5. Proteomic Analysis
2.6. Metabolomic Analysis
2.6.1. Exometabolome Experiments
2.6.2. Assessment of KP-Related Metabolites
2.6.3. Data Processing
2.6.4. Data Normalization
2.7. TCGA and Oncomine Datasets
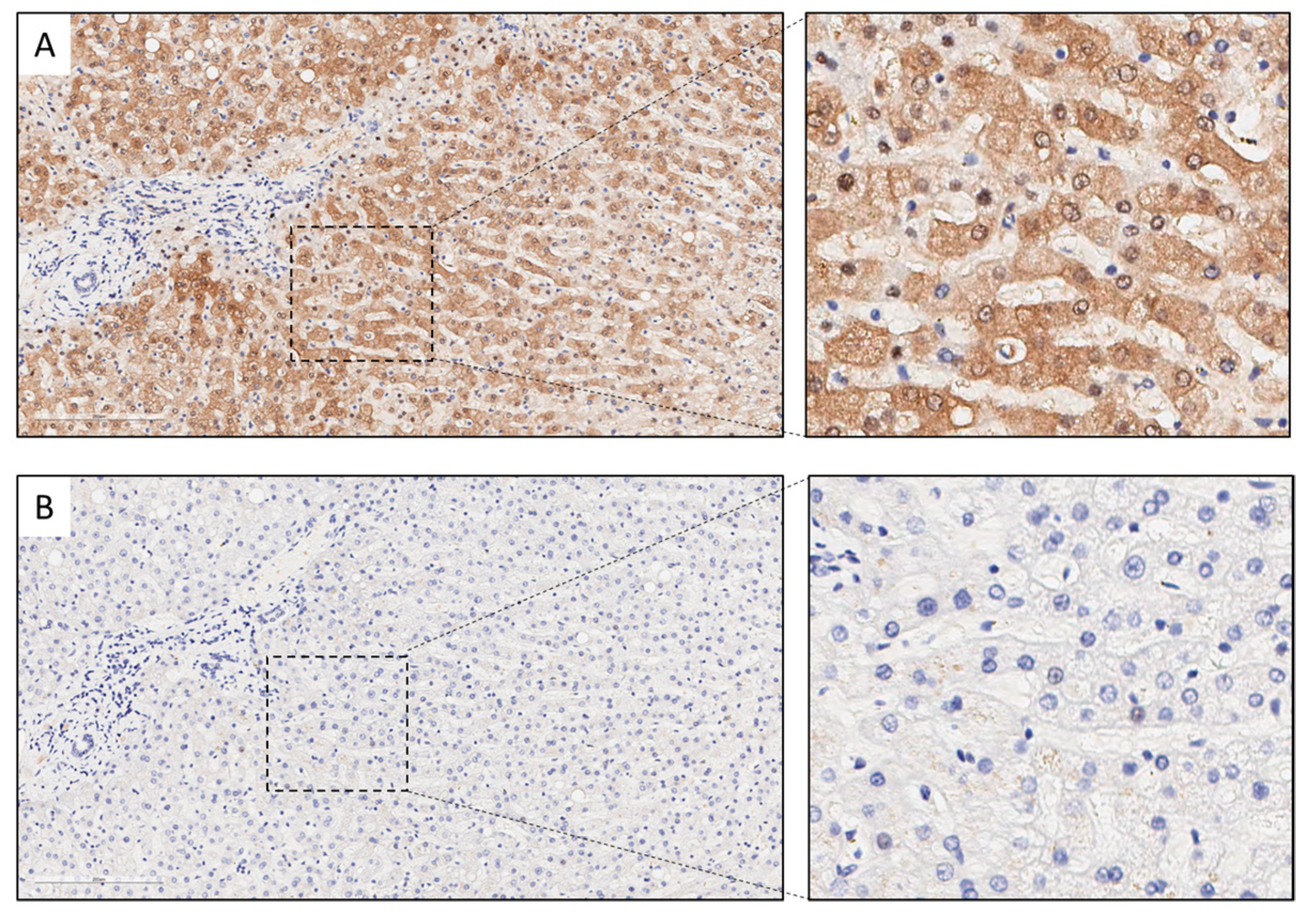
2.8. Immunohistochemistry
2.9. Statistics
3. Results
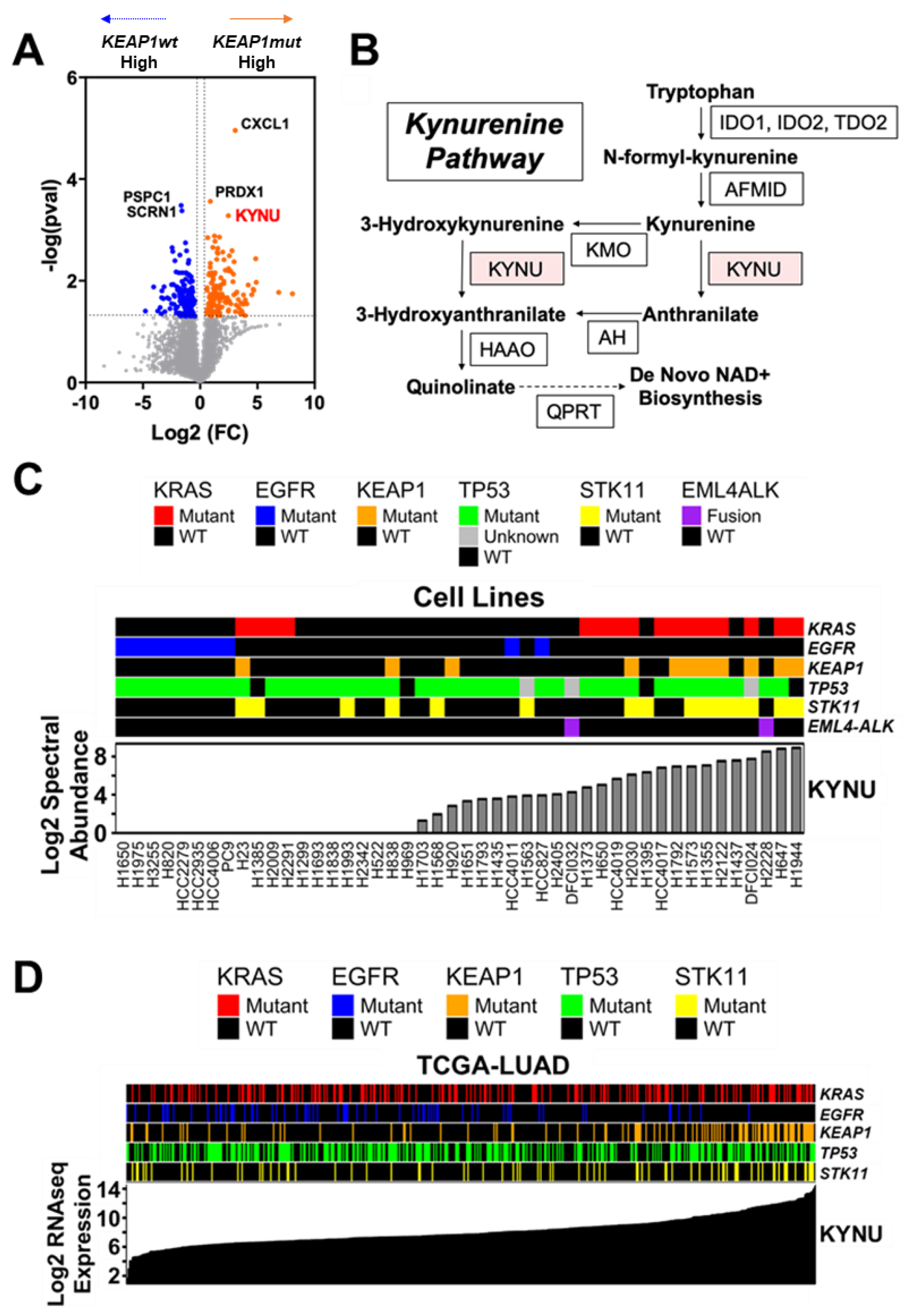
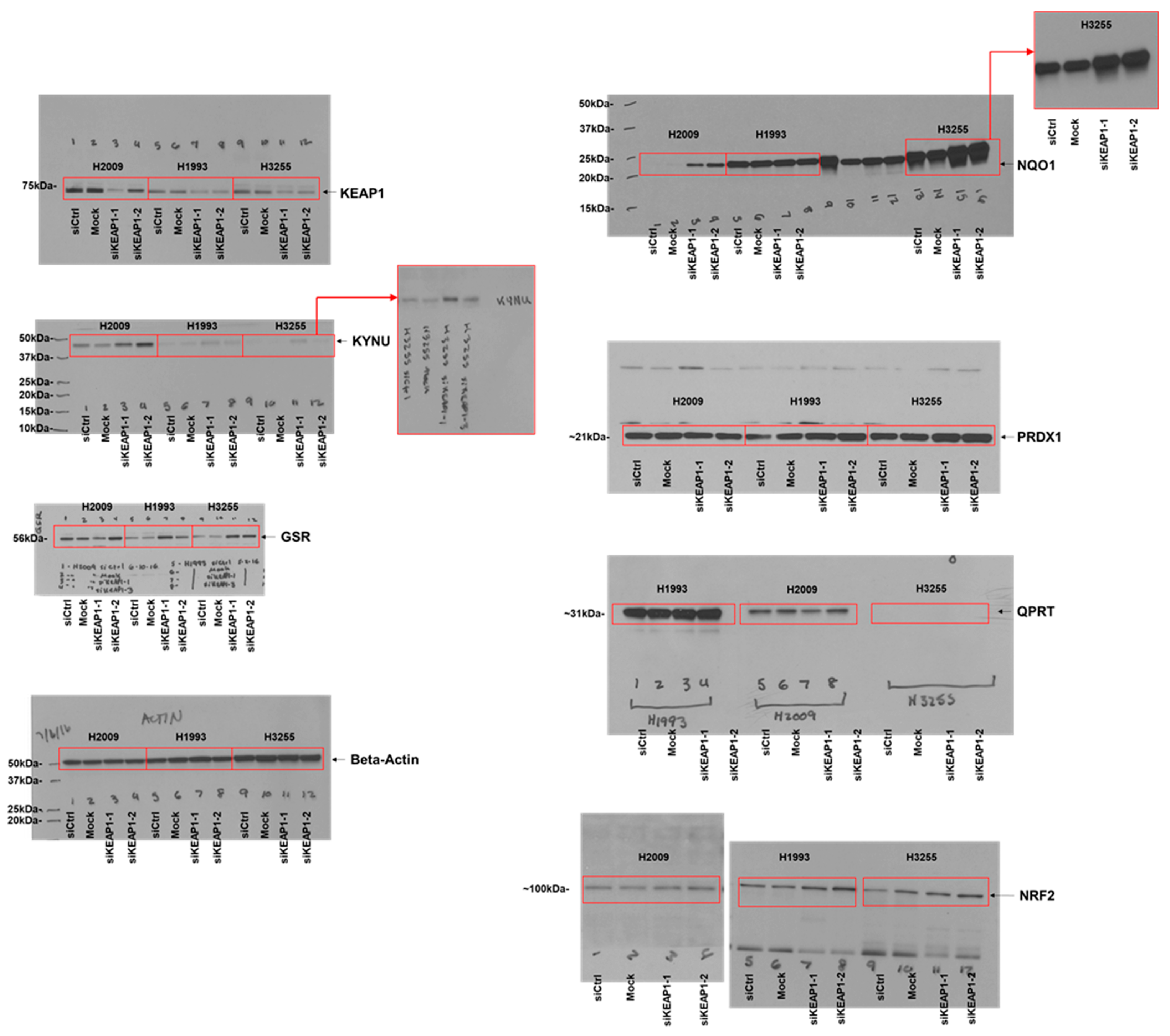
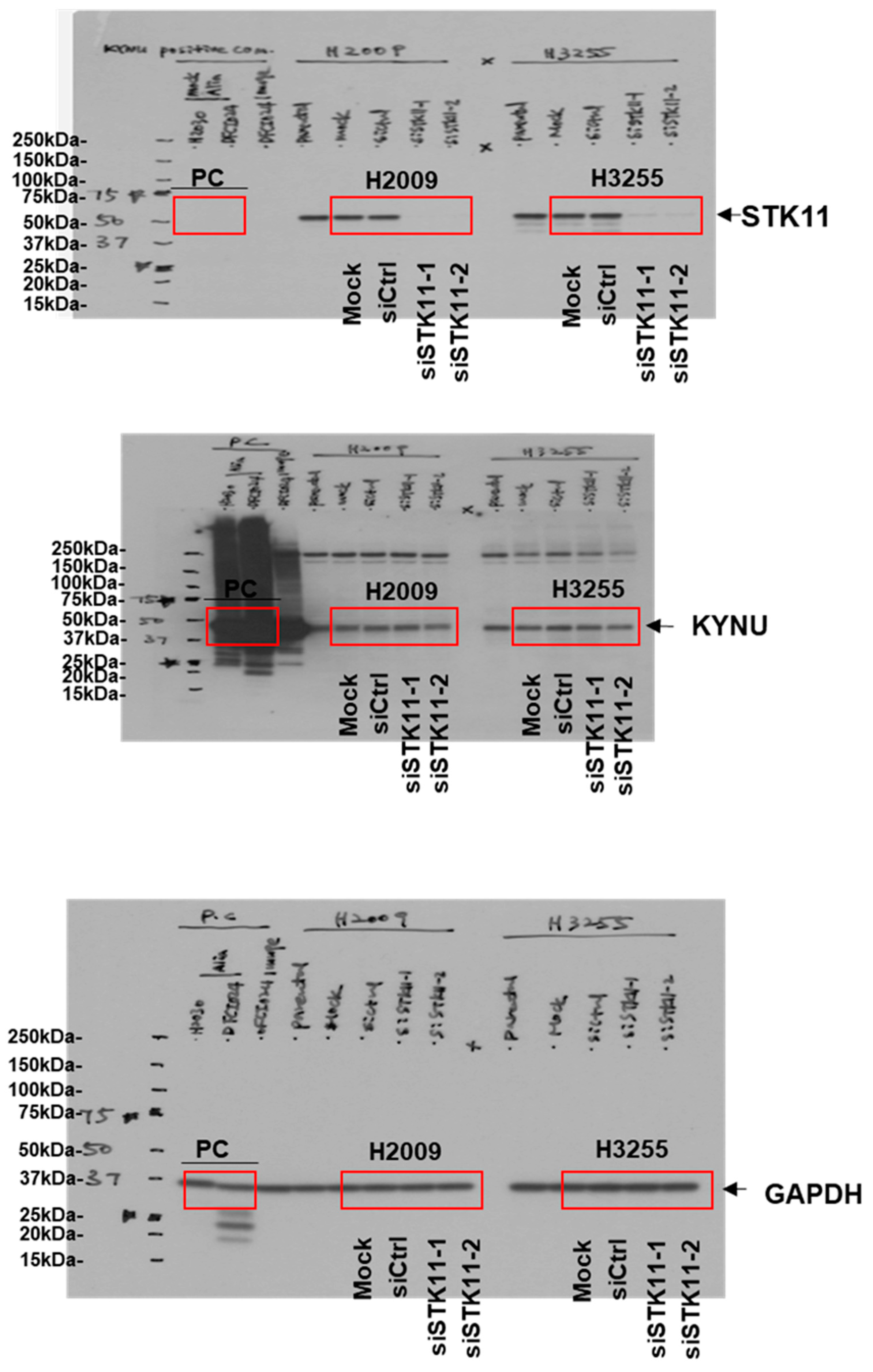
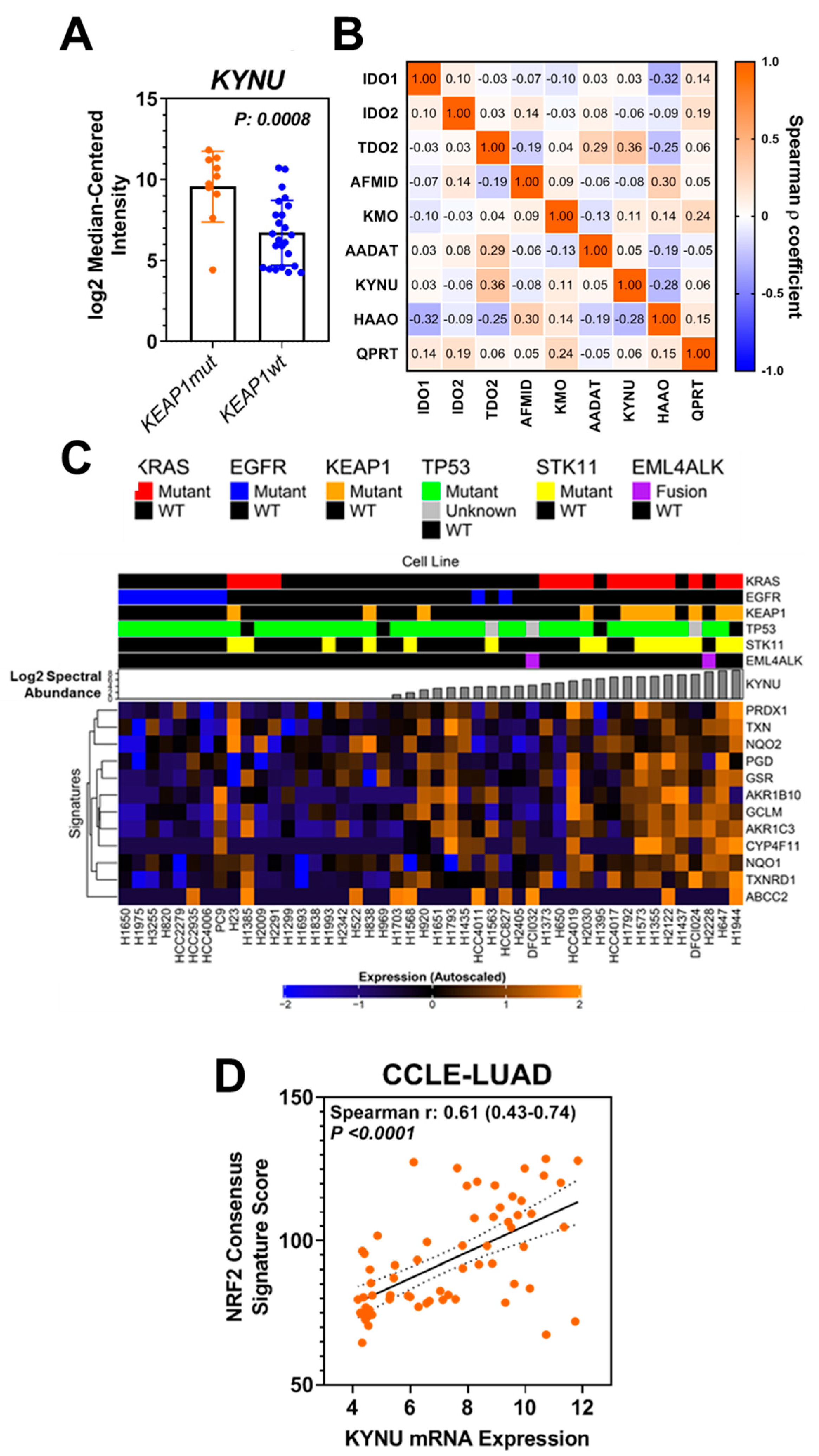
3.1. Analysis of the Protein Signature of KEAP1 Mutation in LUAD Cell Lines
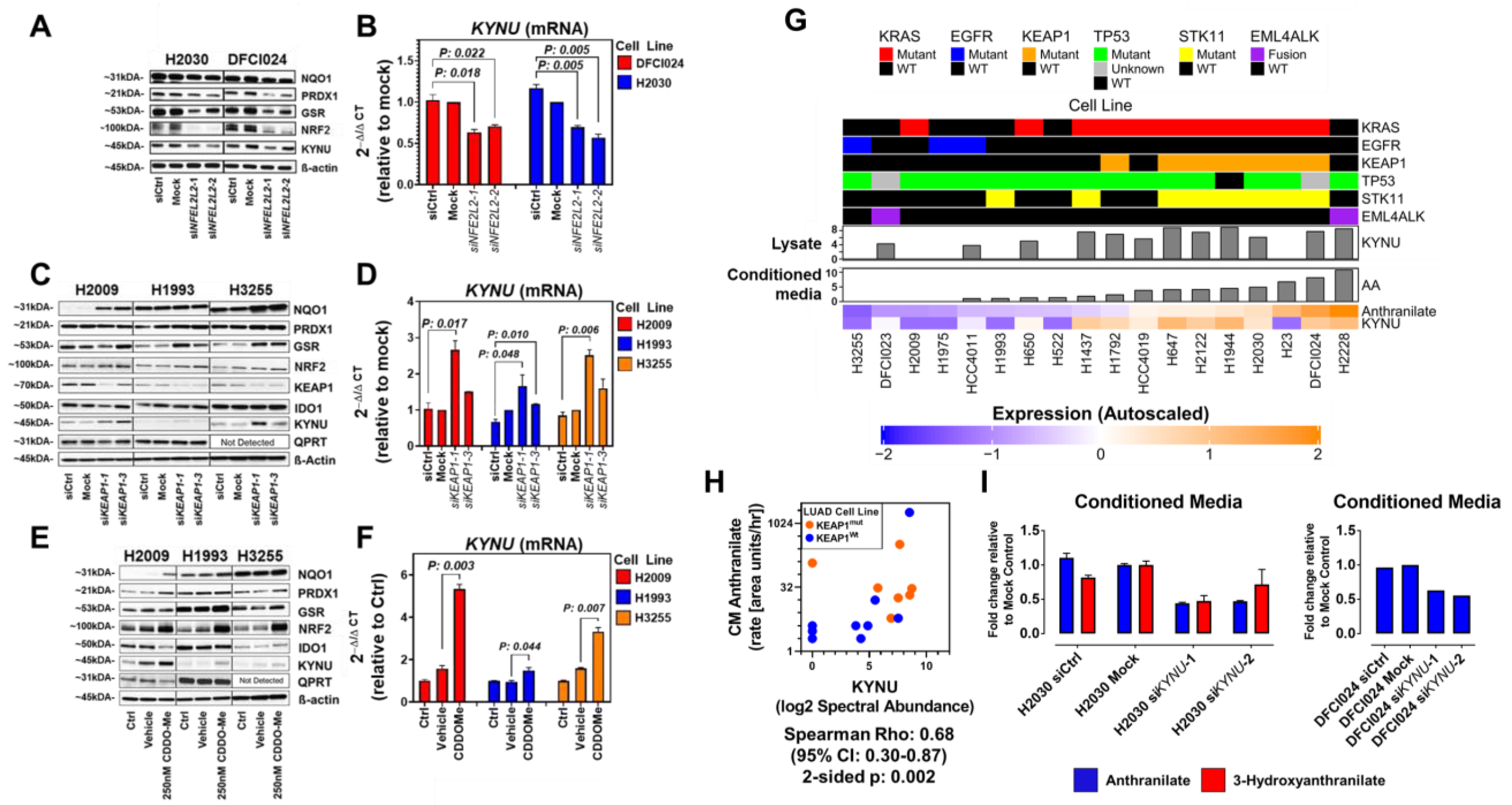
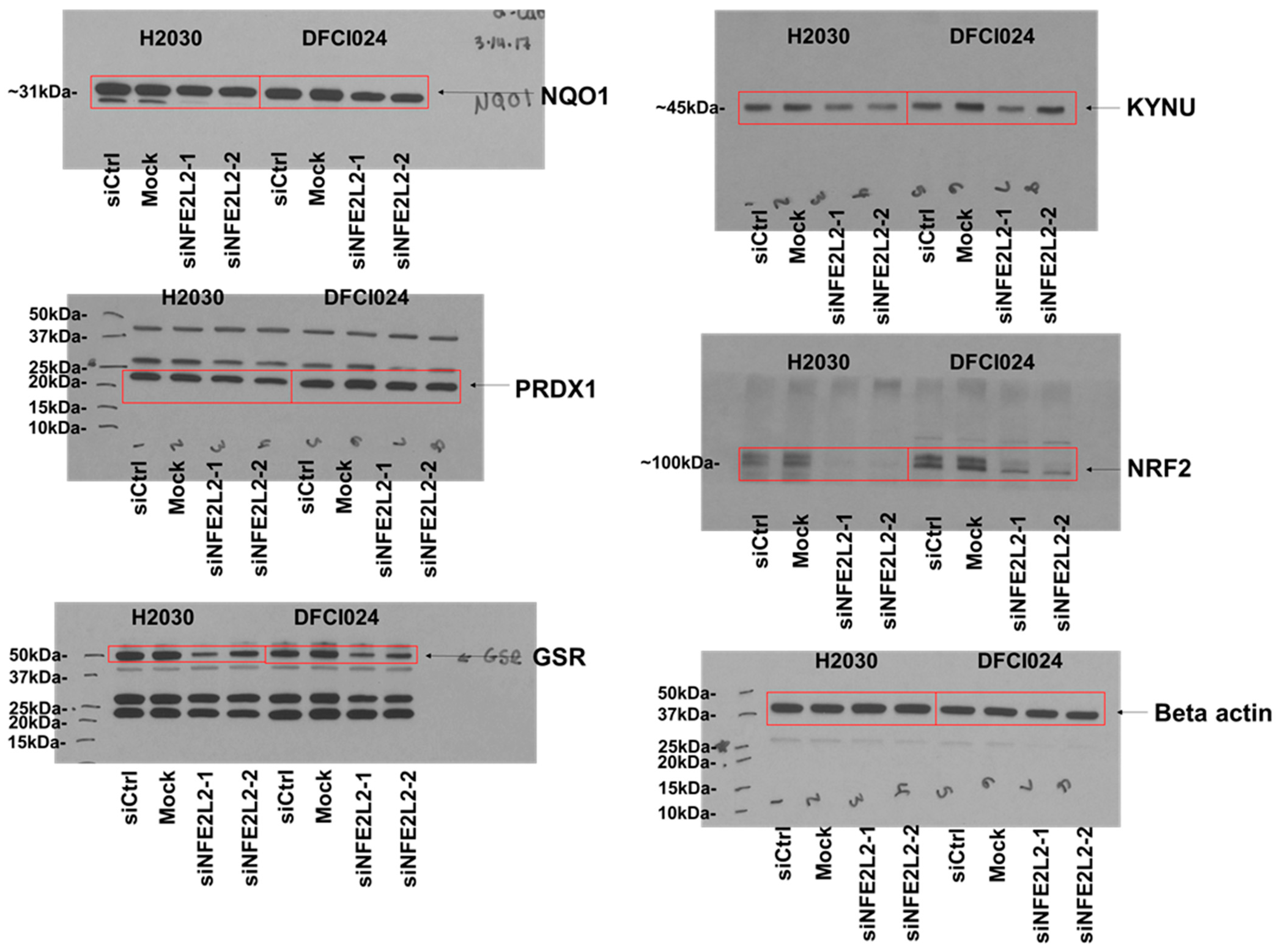
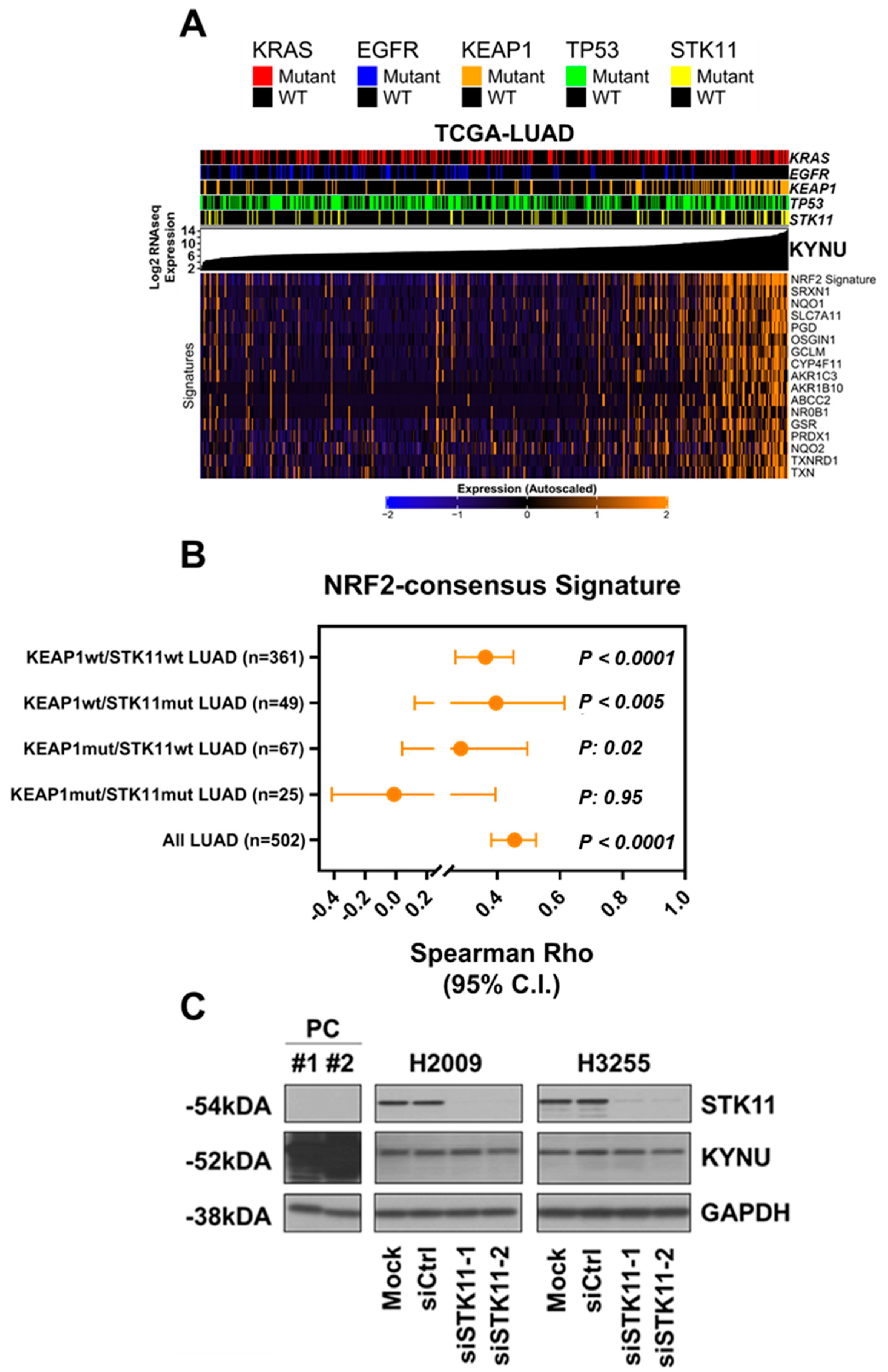
3.2. KYNU Protein Expression Is Regulated by NRF2 Activation in Lung Adenocarcinoma
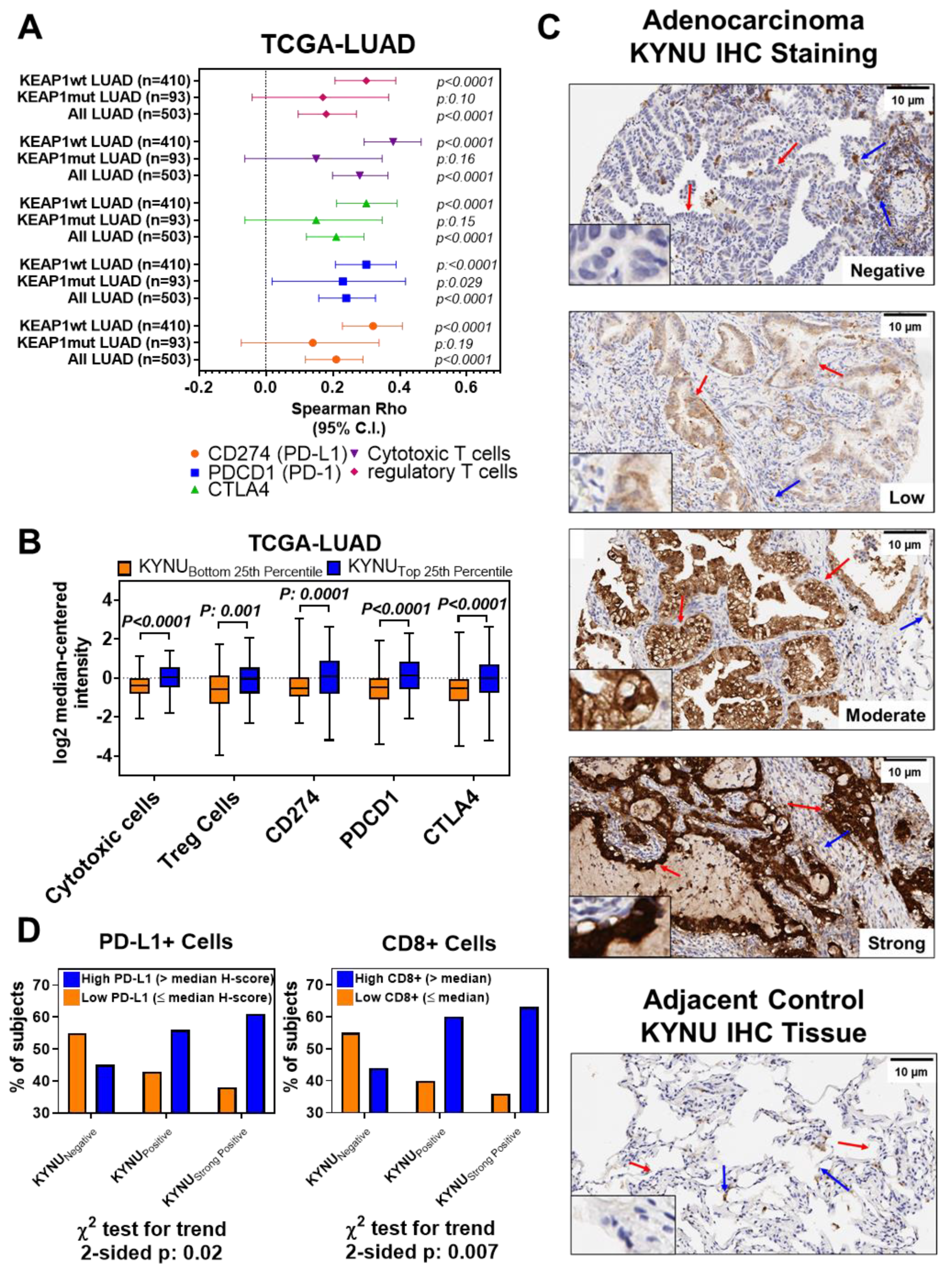
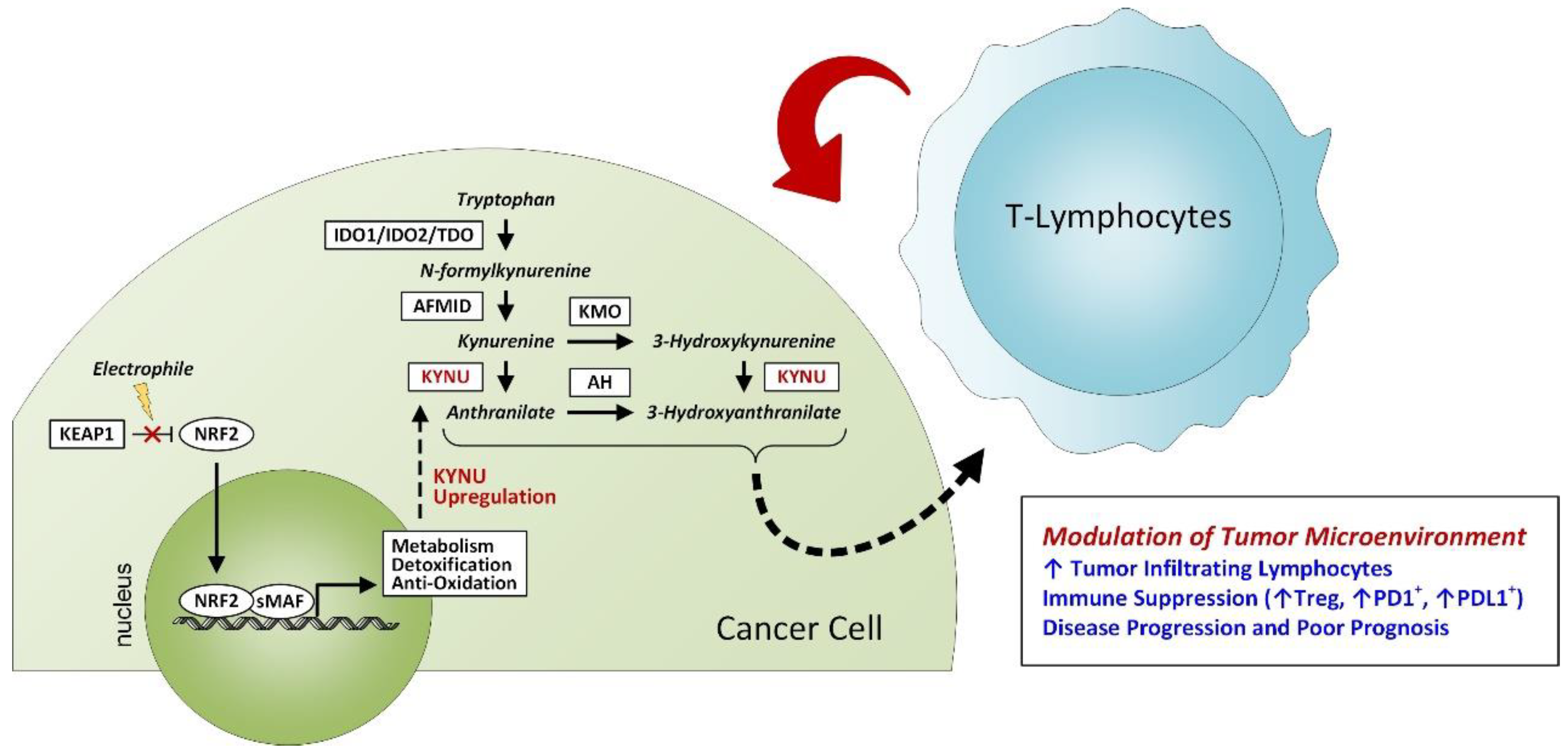
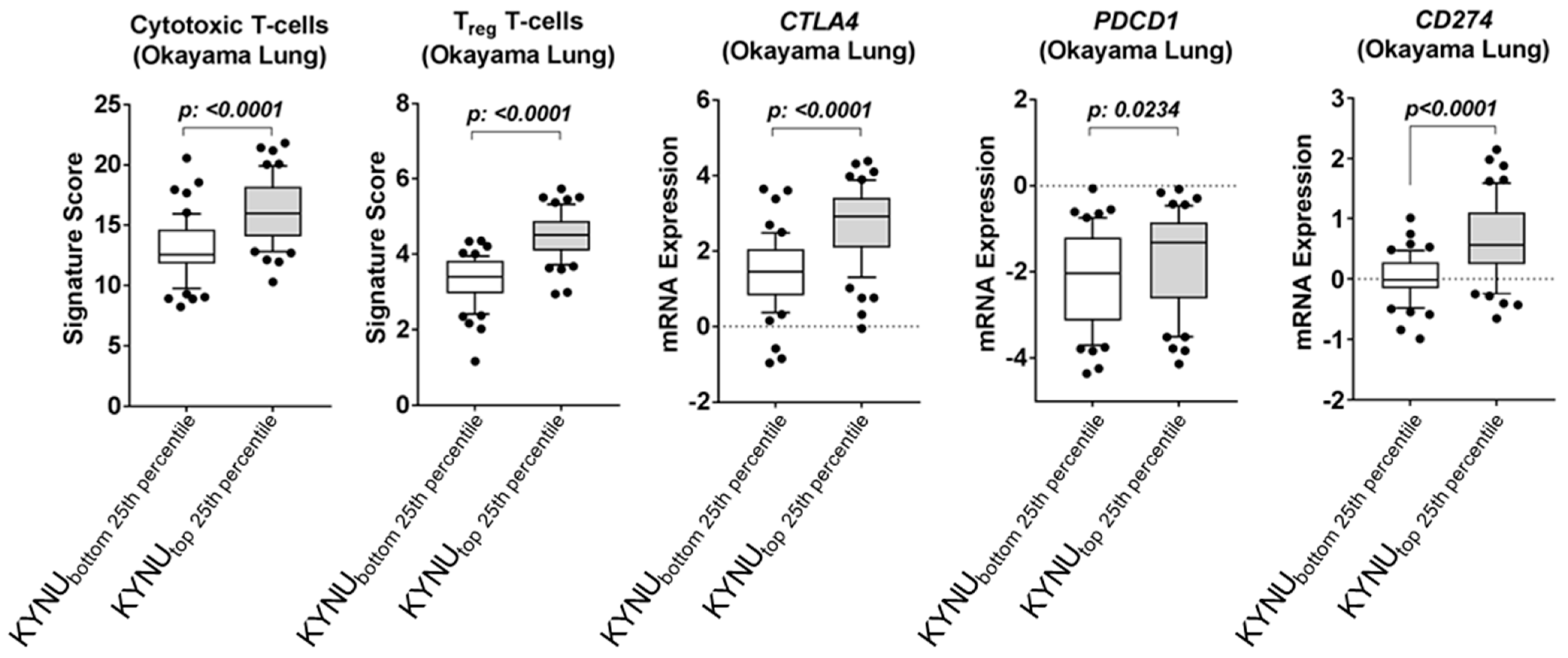
3.3. Association between Tumor KYNU Expression and Tumor Immunophenotype
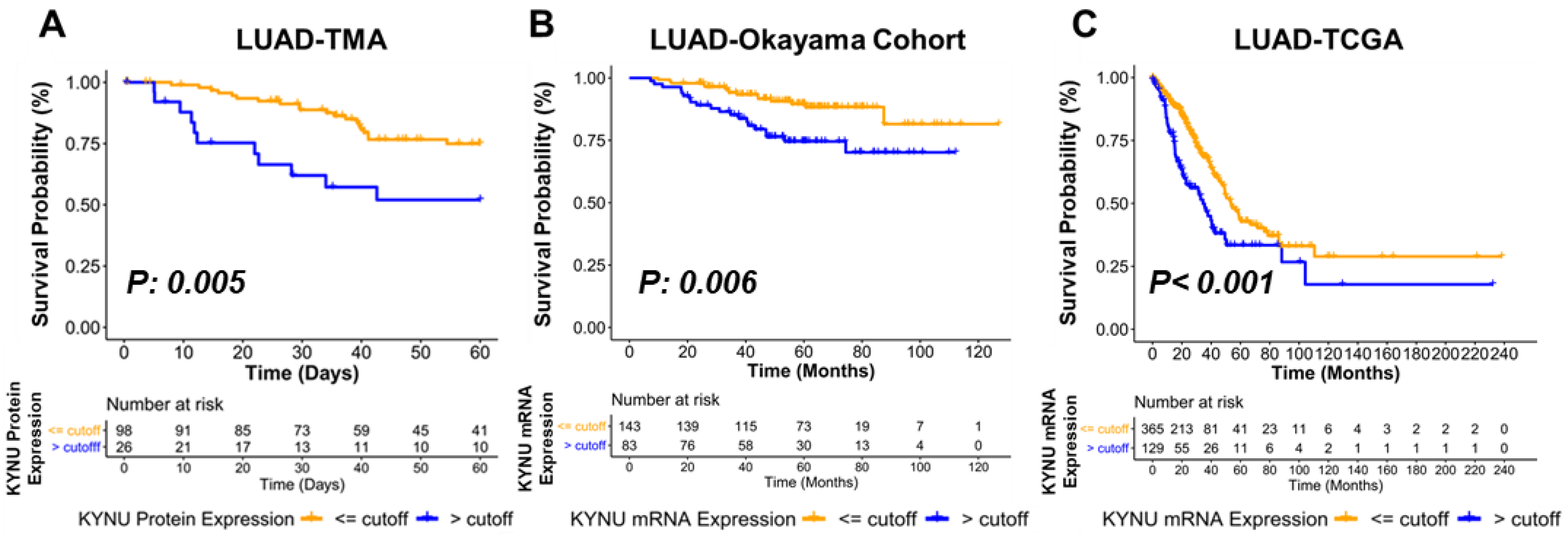
3.4. Association between KYNU Tumor Expression and Overall Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Cell Culture and Transfection
Appendix A.2. Western Blot Analysis
Appendix A.3. RT-PCR Analysis
Appendix A.4. Proteomic Analysis
Appendix A.5. Metabolomic Analysis
Appendix A.5.1. Exometabolome Experiments
Appendix A.5.2. Mass Spectrometry Data Acquisition
Appendix A.5.3. Data Processing
Appendix A.5.4. Data Normalization
Appendix A.6. Immunohistochemical Analysis
Appendix A.7. Statistical Analysis
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | KEAP1 | LKB1 (STK11) | TP53 | EGFR | KRAS | EML4ALK |
|---|---|---|---|---|---|---|
| H23 | Mutant | Mutant | Mutant | WT | Mutant | WT |
| H838 | Mutant | Mutant | Mutant | WT | WT | WT |
| H2030 | Mutant | Mutant | Mutant | WT | Mutant | WT |
| H1573 | Mutant | Mutant | Mutant | WT | Mutant | WT |
| H1792 | Mutant | WT | Mutant | WT | Mutant | WT |
| H1355 | Mutant | Mutant | Mutant | WT | Mutant | WT |
| H2122 | Mutant | Mutant | Mutant | WT | Mutant | WT |
| H647 | Mutant | Mutant | Mutant | WT | Mutant | WT |
| H920 | Mutant | WT | Mutant | WT | WT | WT |
| DFCI024 | Mutant | Mutant | Unknown | WT | Mutant | WT |
| H1944 | Mutant | Mutant | WT | WT | Mutant | WT |
| PC9 | WT | WT | Mutant | Mutant | WT | WT |
| H1299 | WT | WT | Mutant | WT | WT | WT |
| H1975 | WT | WT | Mutant | Mutant | WT | WT |
| H2009 | WT | WT | Mutant | WT | Mutant | WT |
| H1437 | WT | Mutant | Mutant | WT | WT | WT |
| H1650 | WT | WT | Mutant | Mutant | WT | WT |
| H1568 | WT | Mutant | Mutant | WT | WT | WT |
| H650 | WT | WT | Mutant | WT | Mutant | WT |
| H3255 | WT | WT | Mutant | Mutant | WT | WT |
| HCC4019 | WT | WT | Mutant | WT | Mutant | WT |
| H1373 | WT | WT | Mutant | WT | Mutant | WT |
| HCC827 | WT | WT | Mutant | Mutant | WT | WT |
| H1693 | WT | WT | Mutant | WT | WT | WT |
| H1993 | WT | Mutant | Mutant | WT | WT | WT |
| HCC4011 | WT | WT | Mutant | Mutant | WT | WT |
| HCC2279 | WT | WT | Mutant | Mutant | WT | WT |
| H2228 | WT | WT | Mutant | WT | WT | Fusion |
| HCC4006 | WT | WT | Mutant | Mutant | WT | WT |
| H2405 | WT | WT | Mutant | WT | WT | WT |
| H522 | WT | WT | Mutant | WT | WT | WT |
| H820 | WT | WT | Mutant | Mutant | WT | WT |
| H2291 | WT | WT | Mutant | WT | Mutant | WT |
| HCC4017 | WT | WT | Mutant | WT | Mutant | WT |
| HCC2935 | WT | WT | Mutant | Mutant | WT | WT |
| H1703 | WT | WT | Mutant | WT | WT | WT |
| H3255 | WT | WT | Mutant | Mutant | WT | WT |
| H1793 | WT | WT | Mutant | WT | WT | WT |
| H1651 | WT | WT | Mutant | WT | WT | WT |
| H1435 | WT | WT | Mutant | WT | WT | WT |
| H2342 | WT | WT | Mutant | WT | WT | WT |
| H1838 | WT | WT | Mutant | WT | WT | WT |
| DFCI032 | WT | WT | Unknown | WT | WT | Fusion |
| H1563 | WT | Mutant | Unknown | WT | WT | WT |
| H1395 | WT | Mutant | WT | WT | WT | WT |
| H969 | WT | WT | WT | WT | WT | WT |
| H1385 | WT | Mutant | WT | WT | Mutant | WT |
| KYNU Immunostaining | ||||
|---|---|---|---|---|
| Characteristic | Total | Positive, n (%) | Negative, n (%) | p |
| Total | 124 | 31 (25.0) | 93 (75.0) | |
| Sex | ||||
| Male | 67 | 19 | 48 | |
| Female | 57 | 12 | 45 | |
| Age | ||||
| >65 years | 58 | 14 | 44 | |
| ≤65 years | 66 | 17 | 49 | |
| Smoking Status | ||||
| Current | 44 | 17 | 37 | |
| Former | 44 | 13 | 41 | |
| Never | 16 | 1 | 15 | |
| Stage | ||||
| I | 69 | 16 | 53 | 0.2935 |
| II | 29 | 5 | 24 | |
| III | 24 | 9 | 15 | |
| IV | 2 | 1 | 1 | |
| Mutations | ||||
| KRAS | 24 | 11 | 22 | 0.1722 |
| EGFR | 11 | 1 | 13 | |
| KRAS and EGFR wildtype | 48 | 16 | 45 | |
| Cell Line | KEAP1 Status | Whole Lysate KYNU Protein Expression ‡ | Anthranilate †¥ | 3-Hydroxyanthranilate †¥ |
|---|---|---|---|---|
| H647 | Mutant | 368 | 21 ± 9 | - |
| DFCI024 | Mutant | 200 | 326 ± 87 | - |
| H1944 | Mutant | 417 | 30 ± 15 | - |
| H23 | Mutant | 0 | 119 ± 27 | - |
| H1792 | Mutant | 118 | 6 ± 2 | - |
| H2030 | Mutant | 52 | 31 ± 4 | 149 ± 6 |
| H2122 | Mutant | 182 | 18 ± 3 | 3 ± 2 |
| H1437 | WT | 183 | 6 ± 5 | - |
| H2009 | WT | 0 | 2 ± 2 | - |
| H1993 | WT | 0 | 4 ± 3 | - |
| H3255 | WT | 0 | 0 ± 2 | - |
| H2228 | WT | 362 | 1836 ± 279 | - |
| DFCI023 | WT | 18 | 2 ± 3 | - |
| H522 | WT | 0 | 4 ± 3 | - |
| H650 | WT | 28 | 4 ± 3 | - |
| HCC4011 | WT | 13 | 4 ± 3 | - |
| H1975 | WT | 0 | 3 ± 4 | - |
| HCC4019 | WT | 44 | 16 ± 3 | - |
| Variable | Tcga LUAD Dataset | |||||
|---|---|---|---|---|---|---|
| Univariable | Multivariable ¥ | |||||
| Hazard Ratio | 95% CI | Two-Sided p | Hazard Ratio | 95% CI | Two-Sided p | |
| Sex | ||||||
| Female | Reference | Reference | ||||
| Male | 1.05 | 0.78–1.42 | 0.73 | - | ||
| Age | ||||||
| <66 | Reference | Reference | ||||
| ≥66 | 1.28 | 0.95–1.73 | 0.10 | 1.25 | 0.92–1.69 | 0.16 |
| Stage | ||||||
| I | Reference | Reference | ||||
| II | 2.29 | 1.58–3.30 | <0.0001 | 2.04 | 1.40–2.97 | 0.0002 |
| III | 3.56 | 2.43–5.23 | <0.0001 | 3.12 | 2.11–4.59 | <0.0001 |
| IV | 3.61 | 2.05–6.36 | <0.0001 | 3.05 | 1.71–5.42 | 0.0001 |
| KYNU mRNA expression Cutoff ‡ | ||||||
| ≤cutoff | Reference | Reference | ||||
| >cutoff | 1.83 | 1.35–2.50 | 0.0001 | 1.53 | 1.11–2.12 | 0.01 |
| Variable | Okayama LUAD Dataset | |||||
|---|---|---|---|---|---|---|
| Univariable | Multivariable ¥ | |||||
| Hazard Ratio | 95% CI | Two-Sided p | Hazard Ratio | 95% CI | Two-Sided p | |
| Sex | ||||||
| Female | Reference | Reference | ||||
| Male | 1.52 | 0.78–2.96 | 0.22 | - | ||
| Age | ||||||
| <61 | Reference | Reference | ||||
| ≥61 | 1.43 | 0.73–2.78 | 0.29 | 1.68 | 0.86–3.29 | 0.13 |
| Stage | ||||||
| I | Reference | Reference | ||||
| II | 4.23 | 2.17–8.24 | <0.001 | 4.09 | 2.08–8.02 | <0.001 |
| Smoking | ||||||
| Never | Reference | Reference | ||||
| Former/Current | 1.64 | 0.84–3.20 | 0.15 | - | ||
| KYNU mRNA expression Cutoff ‡ | ||||||
| ≤cutoff | Reference | Reference | ||||
| >cutoff | 2.50 | 1.28–4.89 | 0.007 | 2.20 | 1.12–4.33 | 0.022 |








References
- Best, S.A.; Ding, S.; Kersbergen, A.; Dong, X.; Song, J.Y.; Xie, Y.; Reljic, B.; Li, K.; Vince, J.E.; Rathi, V.; et al. Distinct initiating events underpin the immune and metabolic heterogeneity of KRAS-mutant lung adenocarcinoma. Nat. Commun. 2019, 10, 4190. [Google Scholar] [CrossRef]
- Okayama, H.; Kohno, T.; Ishii, Y.; Shimada, Y.; Shiraishi, K.; Iwakawa, R.; Furuta, K.; Tsuta, K.; Shibata, T.; Yamamoto, S.; et al. Identification of genes upregulated in ALK-positive and EGFR/KRAS/ALK-negative lung adenocarcinomas. Cancer Res. 2012, 72, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Tosolini, M.; Kirilovsky, A.; Waldner, M.; Obenauf, A.C.; Angell, H.; Fredriksen, T.; Lafontaine, L.; Berger, A.; et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity 2013, 39, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [CrossRef]
- Skoulidis, F.; Byers, L.A.; Diao, L.; Papadimitrakopoulou, V.A.; Tong, P.; Izzo, J.; Behrens, C.; Kadara, H.; Parra, E.R.; Canales, J.R.; et al. Co-occurring genomic alterations define major subsets of KRAS-mutant lung adenocarcinoma with distinct biology, immune profiles, and therapeutic vulnerabilities. Cancer Discov. 2015, 5, 860–877. [Google Scholar] [CrossRef] [Green Version]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Jeong, Y.; Hellyer, J.A.; Stehr, H.; Hoang, N.T.; Niu, X.; Das, M.; Padda, S.K.; Ramchandran, K.; Neal, J.W.; Wakelee, H.; et al. Role of KEAP1/NFE2L2 Mutations in the Chemotherapeutic Response of Patients with Non-Small Cell Lung Cancer. Clin. Cancer Res. 2020, 26, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Goeman, F.; De Nicola, F.; Scalera, S.; Sperati, F.; Gallo, E.; Ciuffreda, L.; Pallocca, M.; Pizzuti, L.; Krasniqi, E.; Barchiesi, G.; et al. Mutations in the KEAP1-NFE2L2 Pathway Define a Molecular Subset of Rapidly Progressing Lung Adenocarcinoma. J. Thorac. Oncol. 2019, 14, 1924–1934. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hamada, S.; Matsumoto, R.; Taguchi, K.; Yamamoto, M.; Masamune, A. Nrf2 expression in pancreatic stellate cells promotes progression of cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G378–G388. [Google Scholar] [CrossRef]
- Sporn, M.B.; Liby, K.T. NRF2 and cancer: The good, the bad and the importance of context. Nat. Rev. Cancer 2012, 12, 564–571. [Google Scholar] [CrossRef]
- Shibata, T.; Ohta, T.; Tong, K.I.; Kokubu, A.; Odogawa, R.; Tsuta, K.; Asamura, H.; Yamamoto, M.; Hirohashi, S. Cancer related mutations in NRF2 impair its recognition by Keap1-Cul3 E3 ligase and promote malignancy. Proc. Natl. Acad. Sci. USA 2008, 105, 13568–13573. [Google Scholar] [CrossRef] [Green Version]
- Cykowiak, M.; Krajka-Kuźniak, V. Role of Nrf2 in Pancreatic Cancer. Antioxidants 2021, 11, 98. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Montanari, E.; Marzioni, D.; Goteri, G. Role of NRF2 in Ovarian Cancer. Antioxidants 2022, 11, 663. [Google Scholar] [CrossRef]
- Best, S.A.; De Souza, D.P.; Kersbergen, A.; Policheni, A.N.; Dayalan, S.; Tull, D.; Rathi, V.; Gray, D.H.; Ritchie, M.E.; McConville, M.J.; et al. Synergy between the KEAP1/NRF2 and PI3K Pathways Drives Non-Small-Cell Lung Cancer with an Altered Immune Microenvironment. Cell Metab. 2018, 27, 935–943.e934. [Google Scholar] [CrossRef] [Green Version]
- Galan-Cobo, A.; Sitthideatphaiboon, P.; Qu, X.; Poteete, A.; Pisegna, M.A.; Tong, P.; Chen, P.H.; Boroughs, L.K.; Rodriguez, M.L.M.; Zhang, W.; et al. LKB1 and KEAP1/NRF2 Pathways Cooperatively Promote Metabolic Reprogramming with Enhanced Glutamine Dependence in KRAS-Mutant Lung Adenocarcinoma. Cancer Res. 2019, 79, 3251–3267. [Google Scholar] [CrossRef]
- DeNicola, G.M.; Chen, P.H.; Mullarky, E.; Sudderth, J.A.; Hu, Z.; Wu, D.; Tang, H.; Xie, Y.; Asara, J.M.; Huffman, K.E.; et al. NRF2 regulates serine biosynthesis in non-small cell lung cancer. Nat. Genet. 2015, 47, 1475–1481. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Vander Heiden, M.G.; McCormick, F. The Metabolic Landscape of RAS-Driven Cancers from biology to therapy. Nat. Cancer 2021, 2, 271–283. [Google Scholar] [CrossRef]
- Romero, R.; Sayin, V.I.; Davidson, S.M.; Bauer, M.R.; Singh, S.X.; LeBoeuf, S.E.; Karakousi, T.R.; Ellis, D.C.; Bhutkar, A.; Sánchez-Rivera, F.J.; et al. Keap1 loss promotes Kras-driven lung cancer and results in dependence on glutaminolysis. Nat. Med. 2017, 23, 1362–1368. [Google Scholar] [CrossRef] [Green Version]
- Sayin, V.I.; LeBoeuf, S.E.; Singh, S.X.; Davidson, S.M.; Biancur, D.; Guzelhan, B.S.; Alvarez, S.W.; Wu, W.L.; Karakousi, T.R.; Zavitsanou, A.M.; et al. Activation of the NRF2 antioxidant program generates an imbalance in central carbon metabolism in cancer. Elife 2017, 6. [Google Scholar] [CrossRef]
- Phillips, R.S. Structure and mechanism of kynureninase. Arch. Biochem. Biophys. 2014, 544, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Triplett, T.A.; Garrison, K.C.; Marshall, N.; Donkor, M.; Blazeck, J.; Lamb, C.; Qerqez, A.; Dekker, J.D.; Tanno, Y.; Lu, W.C.; et al. Reversal of indoleamine 2,3-dioxygenase-mediated cancer immune suppression by systemic kynurenine depletion with a therapeutic enzyme. Nat. Biotechnol. 2018, 36, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Fahrmann, J.F.; Bantis, L.E.; Capello, M.; Scelo, G.; Dennison, J.B.; Patel, N.; Murage, E.; Vykoukal, J.; Kundnani, D.L.; Foretova, L.; et al. A Plasma-Derived Protein-Metabolite Multiplexed Panel for Early-Stage Pancreatic Cancer. J. Natl. Cancer Inst. 2019, 111, 372–379. [Google Scholar] [CrossRef]
- Fahrmann, J.F.; Vykoukal, J.; Fleury, A.; Tripathi, S.; Dennison, J.B.; Murage, E.; Wang, P.; Yu, C.Y.; Capello, M.; Creighton, C.J.; et al. Association between plasma diacetylspermine and tumor spermine synthase with outcome in triple negative breast cancer. J. Natl. Cancer Inst. 2019, 112, 607–616. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Parra, E.R.; Behrens, C.; Rodriguez-Canales, J.; Lin, H.; Mino, B.; Blando, J.; Zhang, J.; Gibbons, D.L.; Heymach, J.V.; Sepesi, B.; et al. Image Analysis-based Assessment of PD-L1 and Tumor-Associated Immune Cells Density Supports Distinct Intratumoral Microenvironment Groups in Non-small Cell Lung Carcinoma Patients. Clin. Cancer Res. 2016, 22, 6278–6289. [Google Scholar] [CrossRef] [Green Version]
- Cecile Contal, J.O.Q. An application of changepoint methods in studying the effect of age on survival in breast cancer. Comput. Stat. Data Anal. 1999, 30, 253–270. [Google Scholar] [CrossRef]
- Vykoukal, J.; Fahrmann, J.F.; Gregg, J.R.; Tang, Z.; Basourakos, S.; Irajizad, E.; Park, S.; Yang, G.; Creighton, C.J.; Fleury, A.; et al. Caveolin-1-mediated sphingolipid oncometabolism underlies a metabolic vulnerability of prostate cancer. Nat. Commun. 2020, 11, 4279. [Google Scholar] [CrossRef]
- Greulich, H. The genomics of lung adenocarcinoma: Opportunities for targeted therapies. Genes Cancer 2010, 1, 1200–1210. [Google Scholar] [CrossRef]
- Frank, R.; Scheffler, M.; Merkelbach-Bruse, S.; Ihle, M.A.; Kron, A.; Rauer, M.; Ueckeroth, F.; König, K.; Michels, S.; Fischer, R.; et al. Clinical and Pathological Characteristics of KEAP1- and NFE2L2-Mutated Non-Small Cell Lung Carcinoma (NSCLC). Clin. Cancer Res. 2018, 24, 3087–3096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [Green Version]
- Opitz, C.A.; Wick, W.; Steinman, L.; Platten, M. Tryptophan degradation in autoimmune diseases. Cell. Mol. Life Sci. CMLS 2007, 64, 2542–2563. [Google Scholar] [CrossRef] [PubMed]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terness, P.; Bauer, T.M.; Rose, L.; Dufter, C.; Watzlik, A.; Simon, H.; Opelz, G. Inhibition of allogeneic T cell proliferation by indoleamine 2,3-dioxygenase-expressing dendritic cells: Mediation of suppression by tryptophan metabolites. J. Exp. Med. 2002, 196, 447–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, B.; Lim, C.K.; Lovejoy, D.B.; Bessede, A.; Gluch, L.; Guillemin, G.J. Understanding the role of the kynurenine pathway in human breast cancer immunobiology. Oncotarget 2015, 7, 6506–6520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amobi, A.; Qian, F.; Lugade, A.A.; Odunsi, K. Tryptophan Catabolism and Cancer Immunotherapy Targeting IDO Mediated Immune Suppression. Adv. Exp. Med. Biol. 2017, 1036, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Fahrmann, J.F.; Vykoukal, J.V.; Ostrin, E.J. Amino Acid Oncometabolism and Immunomodulation of the Tumor Microenvironment in Lung Cancer. Front. Oncol. 2020, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Platten, M.; Wick, W.; Van den Eynde, B.J. Tryptophan catabolism in cancer: Beyond IDO and tryptophan depletion. Cancer Res. 2012, 72, 5435–5440. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Antonucci, L.; Karin, M. NRF2 as a regulator of cell metabolism and inflammation in cancer. Carcinogenesis 2020, 41, 405–416. [Google Scholar] [CrossRef]
- Gwinn, D.M.; Lee, A.G.; Briones-Martin-del-Campo, M.; Conn, C.S.; Simpson, D.R.; Scott, A.I.; Le, A.; Cowan, T.M.; Ruggero, D.; Sweet-Cordero, E.A. Oncogenic KRAS Regulates Amino Acid Homeostasis and Asparagine Biosynthesis via ATF4 and Alters Sensitivity to L-Asparaginase. Cancer Cell 2018, 33, 91–107.e106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunelli, L.; Caiola, E.; Marabese, M.; Broggini, M.; Pastorelli, R. Capturing the metabolomic diversity of KRAS mutants in non-small-cell lung cancer cells. Oncotarget 2014, 5, 4722–4731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najumudeen, A.K.; Ceteci, F.; Fey, S.K.; Hamm, G.; Steven, R.T.; Hall, H.; Nikula, C.J.; Dexter, A.; Murta, T.; Race, A.M.; et al. The amino acid transporter SLC7A5 is required for efficient growth of KRAS-mutant colorectal cancer. Nat. Genet. 2021, 53, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.H.; Sun, I.H.; Zhao, L.; Leone, R.D.; Sun, I.M.; Xu, W.; Collins, S.L.; Tam, A.J.; Blosser, R.L.; Patel, C.H.; et al. Targeting glutamine metabolism enhances tumor-specific immunity by modulating suppressive myeloid cells. J. Clin. Investig. 2020, 130, 3865–3884. [Google Scholar] [CrossRef]
- Wu, W.C.; Sun, H.W.; Chen, J.; OuYang, H.Y.; Yu, X.J.; Chen, H.T.; Shuang, Z.Y.; Shi, M.; Wang, Z.; Zheng, L. Immunosuppressive Immature Myeloid Cell Generation Is Controlled by Glutamine Metabolism in Human Cancer. Cancer Immunol. Res. 2019, 7, 1605–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, E.L.; Kelman, A.; Wu, G.S.; Gopaul, R.; Senkevitch, E.; Aghvanyan, A.; Turay, A.M.; Frauwirth, K.A. Glutamine uptake and metabolism are coordinately regulated by ERK/MAPK during T lymphocyte activation. J. Immunol. 2010, 185, 1037–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Raynor, J.; Nguyen, T.-L.M.; Chi, H. Nutrient and Metabolic Sensing in T Cell Responses. Front. Immunol. 2017, 8, 247. [Google Scholar] [CrossRef] [Green Version]
- Sonner, J.K.; Deumelandt, K.; Ott, M.; Thomé, C.M.; Rauschenbach, K.J.; Schulz, S.; Munteanu, B.; Mohapatra, S.; Adam, I.; Hofer, A.C.; et al. The stress kinase GCN2 does not mediate suppression of antitumor T cell responses by tryptophan catabolism in experimental melanomas. Oncoimmunology 2016, 5, e1240858. [Google Scholar] [CrossRef] [Green Version]
- Quintana, F.J.; Sherr, D.H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol. Rev. 2013, 65, 1148–1161. [Google Scholar] [CrossRef] [Green Version]
- Mándi, Y.; Vécsei, L. The kynurenine system and immunoregulation. J. Neural Transm. 2012, 119, 197–209. [Google Scholar] [CrossRef]
- Li, C.; Zhao, H. Tryptophan and Its Metabolites in Lung Cancer: Basic Functions and Clinical Significance. Front. Oncol. 2021, 11, 707277. [Google Scholar] [CrossRef] [PubMed]
- Long, G.V.; Dummer, R.; Hamid, O.; Gajewski, T.F.; Caglevic, C.; Dalle, S.; Arance, A.; Carlino, M.S.; Grob, J.J.; Kim, T.M.; et al. Epacadostat plus pembrolizumab versus placebo plus pembrolizumab in patients with unresectable or metastatic melanoma (ECHO-301/KEYNOTE-252): A phase 3, randomised, double-blind study. Lancet Oncol. 2019, 20, 1083–1097. [Google Scholar] [CrossRef]
- Weber, W.P.; Feder-Mengus, C.; Chiarugi, A.; Rosenthal, R.; Reschner, A.; Schumacher, R.; Zajac, P.; Misteli, H.; Frey, D.M.; Oertli, D.; et al. Differential effects of the tryptophan metabolite 3-hydroxyanthranilic acid on the proliferation of human CD8+ T cells induced by TCR triggering or homeostatic cytokines. Eur. J. Immunol. 2006, 36, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.P.; Fu, S.F.; Chen, X.; Ye, J.; Ye, Y.; Kong, L.D.; Zhu, Z. The Clinicopathological and Prognostic Significance of IDO1 Expression in Human Solid Tumors: Evidence from a Systematic Review and Meta-Analysis. Cell Physiol. Biochem. 2018, 49, 134–143. [Google Scholar] [CrossRef]
- Chuang, S.C.; Fanidi, A.; Ueland, P.M.; Relton, C.; Midttun, O.; Vollset, S.E.; Gunter, M.J.; Seckl, M.J.; Travis, R.C.; Wareham, N.; et al. Circulating biomarkers of tryptophan and the kynurenine pathway and lung cancer risk. Cancer Epidemiol. Biomarkers Prev. 2014, 23, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Huang, L.; Jin, J.Y.; Pi, W.; Ellsworth, S.G.; Jolly, S.; Mellor, A.L.; Machtay, M.; Kong, F.S. A Validation Study on IDO Immune Biomarkers for Survival Prediction in Non-Small Cell Lung Cancer: Radiation Dose Fractionation Effect in Early-Stage Disease. Clin. Cancer Res. 2020, 26, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Huang, L.; Jin, J.Y.; Jolly, S.; Zang, Y.; Wu, H.; Yan, L.; Pi, W.; Li, L.; Mellor, A.L.; et al. IDO Immune Status after Chemoradiation May Predict Survival in Lung Cancer Patients. Cancer Res. 2018, 78, 809–816. [Google Scholar] [CrossRef] [Green Version]
- Karayama, M.; Masuda, J.; Mori, K.; Yasui, H.; Hozumi, H.; Suzuki, Y.; Furuhashi, K.; Fujisawa, T.; Enomoto, N.; Nakamura, Y.; et al. Comprehensive assessment of multiple tryptophan metabolites as potential biomarkers for immune checkpoint inhibitors in patients with non-small cell lung cancer. Clin. Transl. Oncol. 2020, 23, 418–423. [Google Scholar] [CrossRef]
- Qian, Z.; Zhou, T.; Gurguis, C.I.; Xu, X.; Wen, Q.; Lv, J.; Fang, F.; Hecker, L.; Cress, A.E.; Natarajan, V.; et al. Nuclear factor, erythroid 2-like 2-associated molecular signature predicts lung cancer survival. Sci. Rep. 2015, 5, 16889. [Google Scholar] [CrossRef] [Green Version]
- Arbour, K.C.; Jordan, E.; Kim, H.R.; Dienstag, J.; Yu, H.A.; Sanchez-Vega, F.; Lito, P.; Berger, M.; Solit, D.B.; Hellmann, M.; et al. Effects of Co-occurring Genomic Alterations on Outcomes in Patients with KRAS-Mutant Non-Small Cell Lung Cancer. Clin. Cancer Res. 2018, 24, 334–340. [Google Scholar] [CrossRef] [Green Version]





| Variable | LUAD TMA | |||||
|---|---|---|---|---|---|---|
| Univariable | Multivariable ‡ | |||||
| HR | 95% C.I. | p | HR | 95% C.I. | p | |
| Sex | ||||||
| Female | Reference | Reference | ||||
| Male | 1.42 | 0.70–2.89 | 0.330 | - | ||
| Age ¥ | ||||||
| <65 | Reference | Reference | ||||
| ≥65 | 0.92 | 0.45–1.86 | 0.810 | - | ||
| Stage | ||||||
| I | Reference | Reference | ||||
| II | 3.69 | 1.43–9.53 | 0.007 | 4.12 | 1.59–10.69 | 0.004 |
| III | 7.74 | 2.98–20.10 | <0.001 | 8.73 | 3.31–22.99 | <0.001 |
| IV | 12.28 | 2.54–59.38 | 0.002 | 6.46 | 1.27–32.78 | 0.024 |
| Smoking | ||||||
| Never | Reference | Reference | ||||
| Former | 1.11 | 0.36–3.40 | 0.860 | - | ||
| Current | 1.23 | 0.40–3.73 | 0.720 | - | ||
| KYNU Staining † | ||||||
| ≤Cutoff | Reference | Reference | ||||
| >Cutoff | 2.77 | 1.33–5.80 | 0.007 | 3.11 | 1.41–6.84 | 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahrmann, J.F.; Tanaka, I.; Irajizad, E.; Mao, X.; Dennison, J.B.; Murage, E.; Casabar, J.; Mayo, J.; Peng, Q.; Celiktas, M.; et al. Mutational Activation of the NRF2 Pathway Upregulates Kynureninase Resulting in Tumor Immunosuppression and Poor Outcome in Lung Adenocarcinoma. Cancers 2022, 14, 2543. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102543
Fahrmann JF, Tanaka I, Irajizad E, Mao X, Dennison JB, Murage E, Casabar J, Mayo J, Peng Q, Celiktas M, et al. Mutational Activation of the NRF2 Pathway Upregulates Kynureninase Resulting in Tumor Immunosuppression and Poor Outcome in Lung Adenocarcinoma. Cancers. 2022; 14(10):2543. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102543
Chicago/Turabian StyleFahrmann, Johannes F., Ichidai Tanaka, Ehsan Irajizad, Xiangying Mao, Jennifer B. Dennison, Eunice Murage, Julian Casabar, Jeffrey Mayo, Qian Peng, Muge Celiktas, and et al. 2022. "Mutational Activation of the NRF2 Pathway Upregulates Kynureninase Resulting in Tumor Immunosuppression and Poor Outcome in Lung Adenocarcinoma" Cancers 14, no. 10: 2543. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102543