
ADCK2 Knockdown Affects the Migration of Melanoma Cells via MYL6

 , , and
, , and
Abstract
:Simple Summary
Abstract
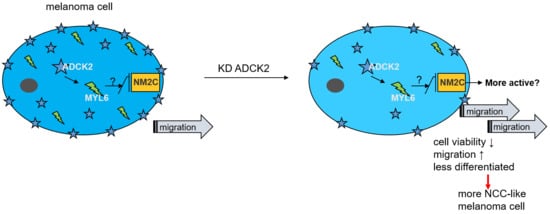
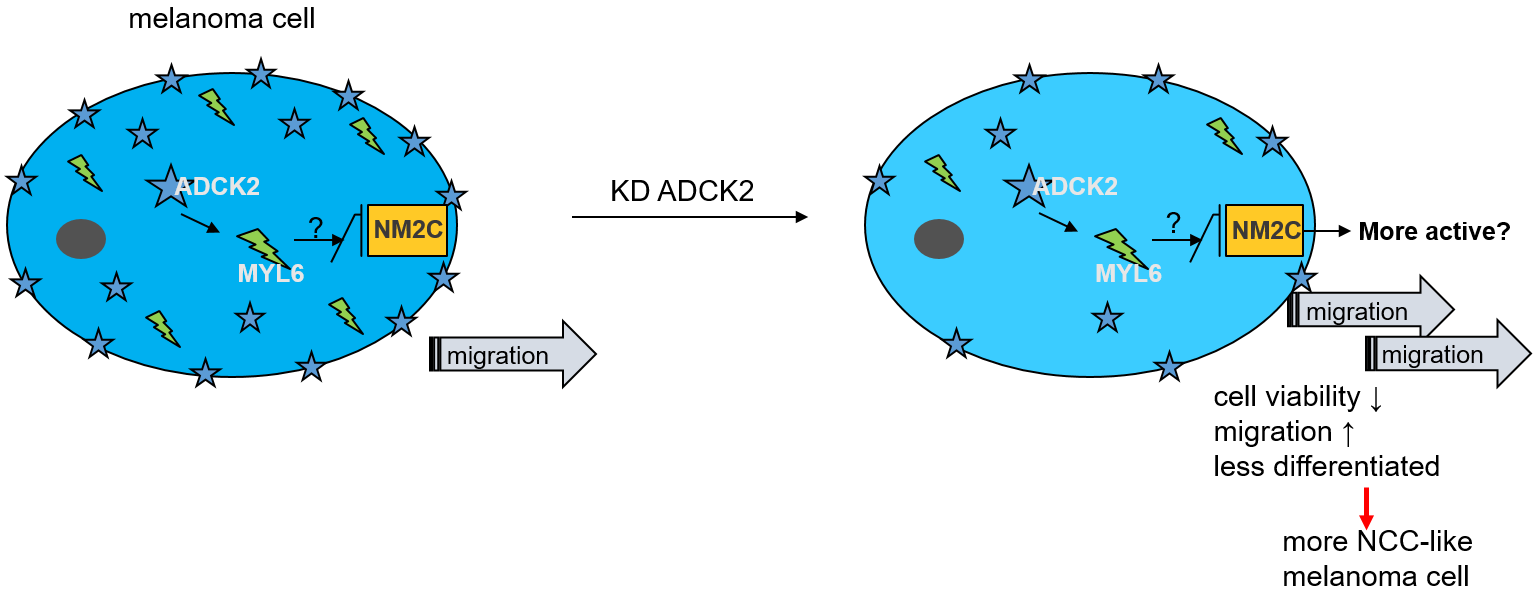{kind=link}
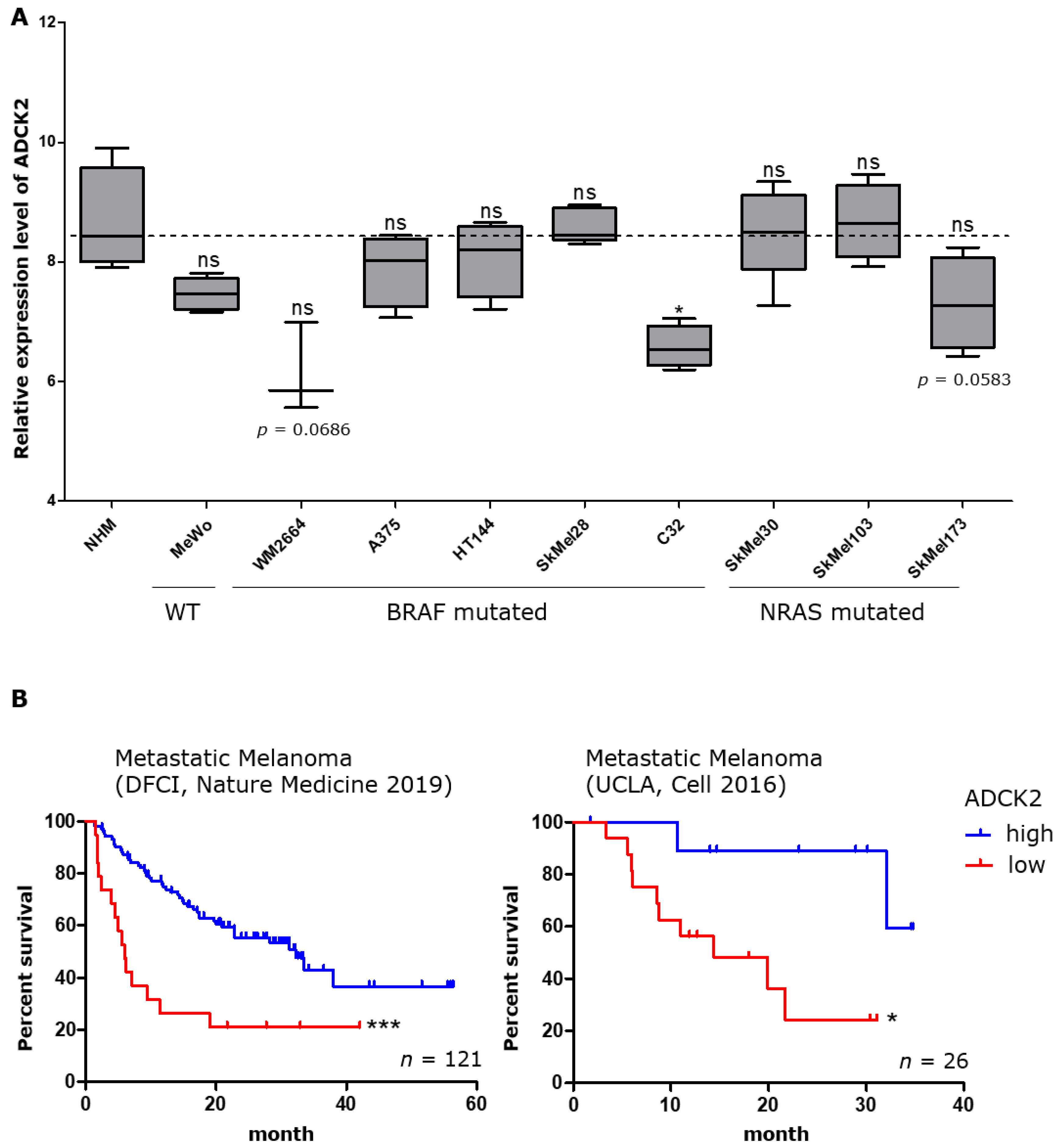{kind=link}
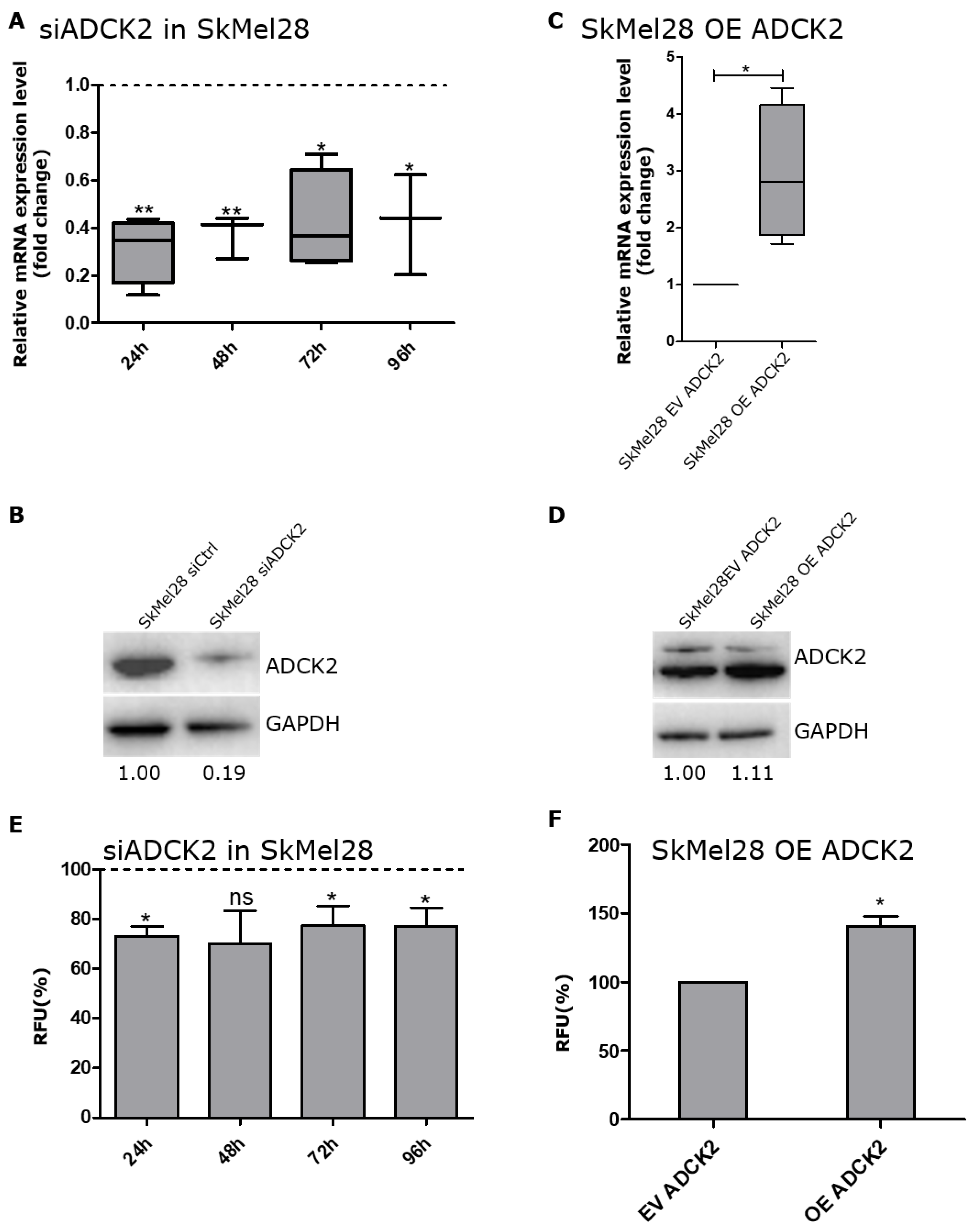{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. siRNA Transfection and Overexpression
2.3. qPCR
2.4. Western Blot
2.5. Immunofluorescence Staining (IF)
2.6. Cell Viability Assay
2.7. Migration Assay
2.8. Invasion Assay
2.9. Gene Expression Analysis
2.10. Statistical Analysis
3. Results
3.1. Patients with High Intratumoral ADCK2 Expression Have a Better Prognosis
3.2. ADCK2 Is Important for the Cell Viability of Melanoma Cells
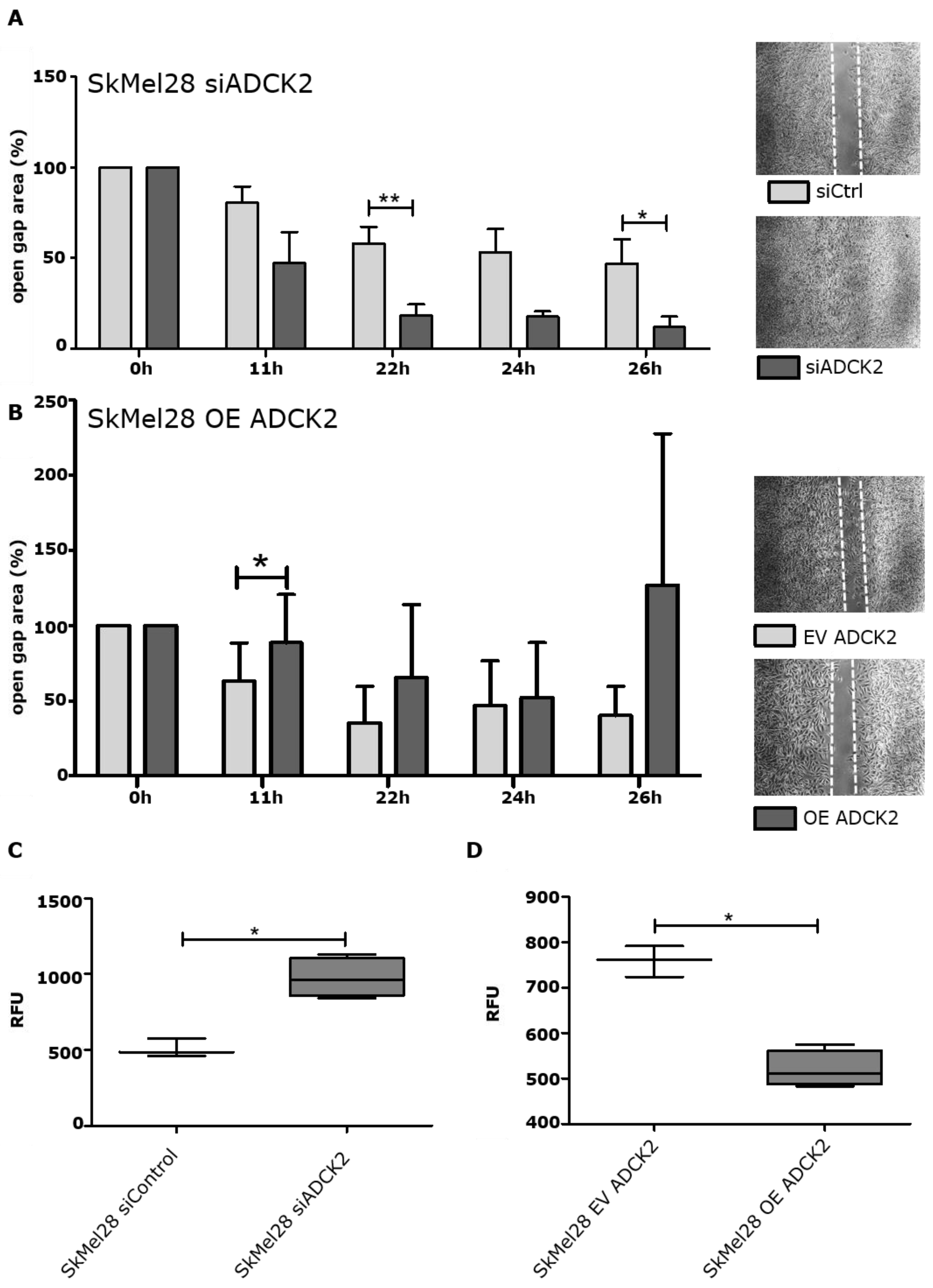
3.3. ADCK2 Limits the Migration and Invasion of Melanoma Cells
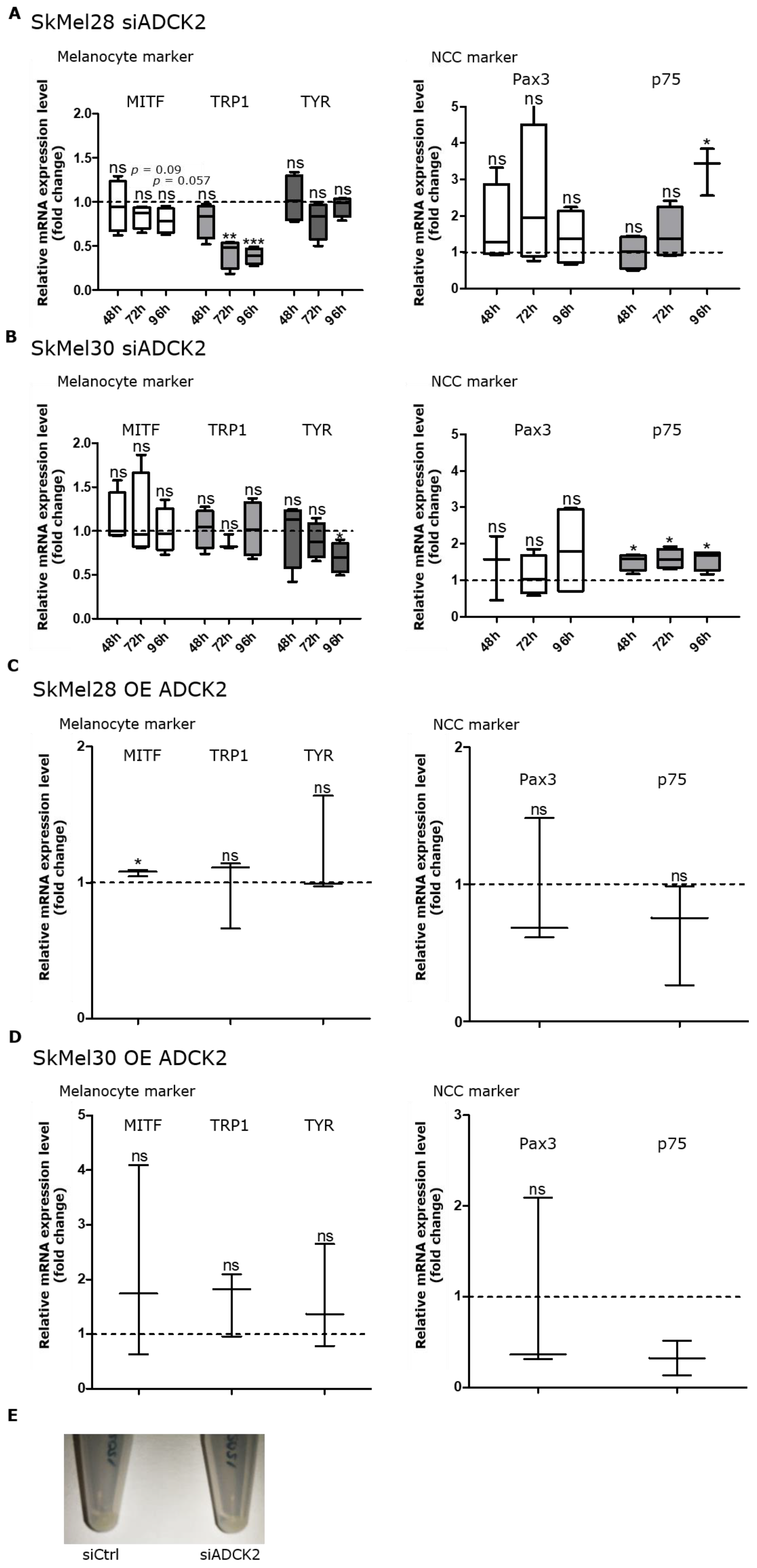
3.4. ADCK2 Affects the Differentiation Status of Melanoma Cells
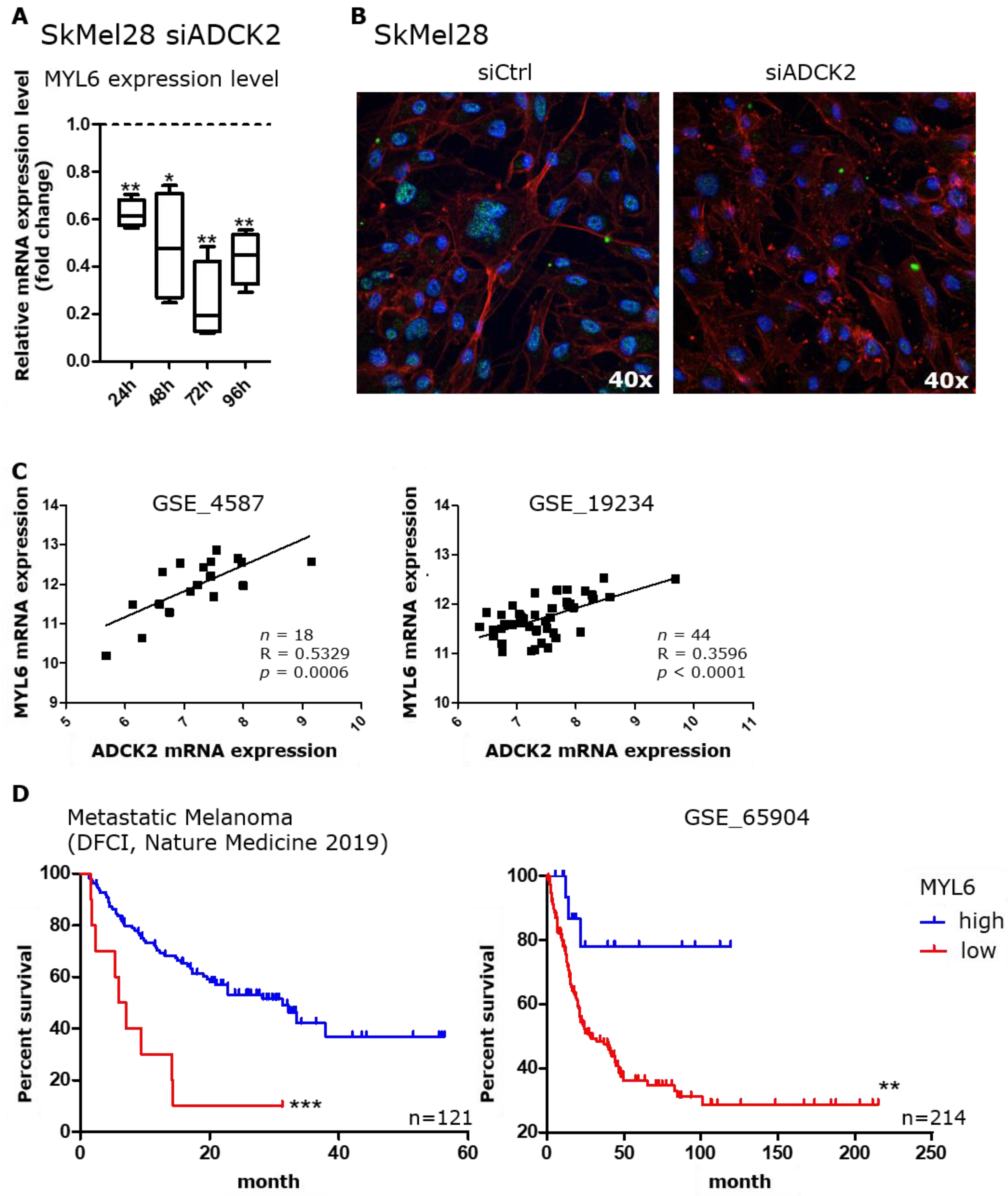
3.5. MYL6 Is Functionally Connected to ADCK2
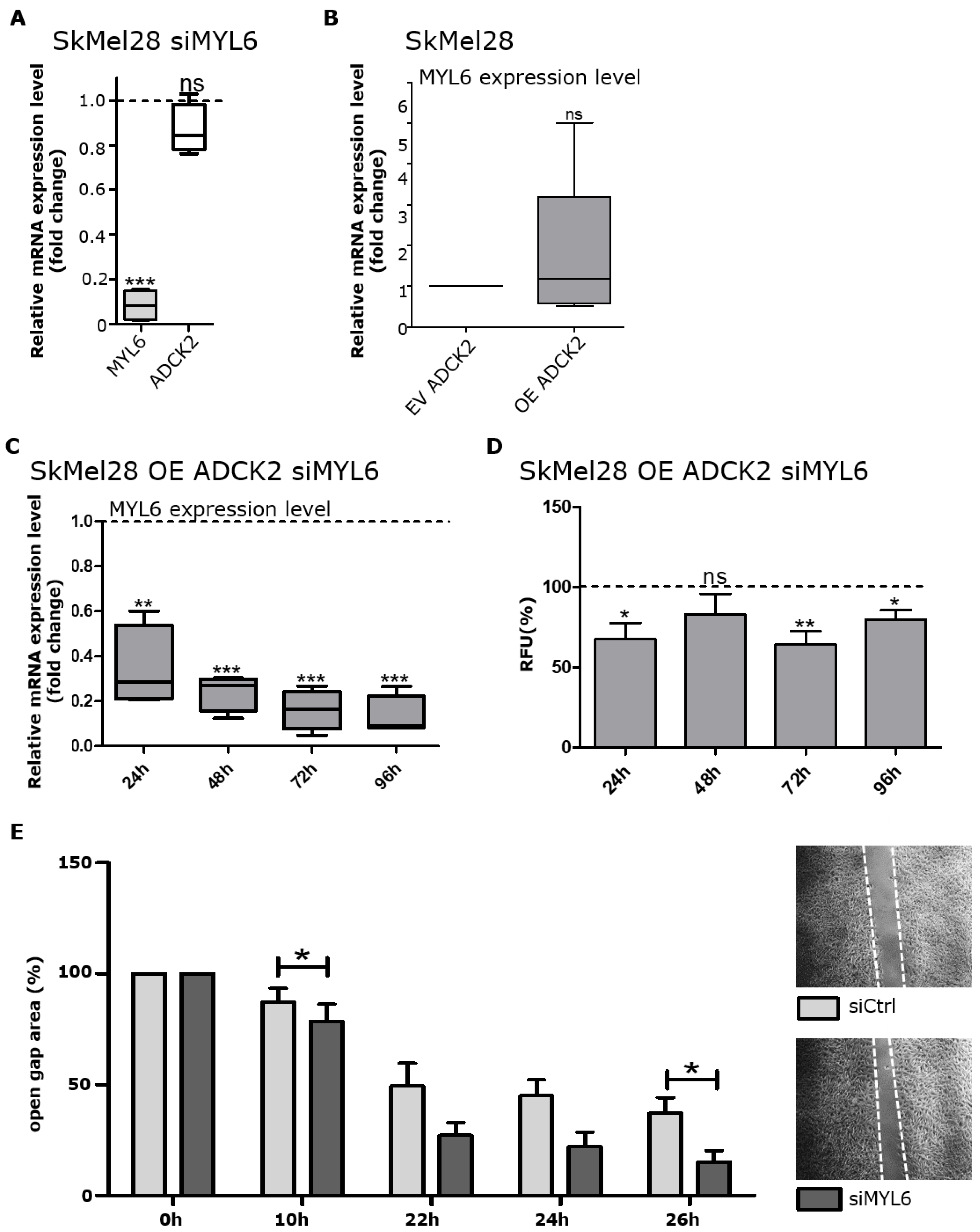
3.6. Knockdown of MYL6 Negates the Effects of ADCK2 Overexpression on Melanoma Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, M.Z.; Guo, C.; Yin, Y.; Lin, J.; Liu, S. Responding proteins for hacat cells against 2,4-dinitrobenzene sulfonic acid stimulation-A proteomic study. J. Anim. Plant Sci. 2016, 26, 233–241. [Google Scholar]
- Bakos, R.M.; Maier, T.; Besch, R.; Mestel, D.S.; Ruzicka, T.; Sturm, R.A.; Berking, C. Nestin and SOX9 and SOX10 transcription factors are coexpressed in melanoma. Exp. Dermatol. 2010, 19, e89–e94. [Google Scholar] [CrossRef] [PubMed]
- Wessely, A.; Steeb, T.; Berking, C.; Heppt, M.V. How neural crest transcription factors contribute to melanoma heterogeneity, cellular plasticity, and treatment resistance. Int. J. Mol. Sci. 2021, 22, 5761. [Google Scholar] [CrossRef] [PubMed]
- De Wit, N.J.W.; Rijntjes, J.; Diepstra, J.H.S.; Van Kuppevelt, T.H.; Weidle, U.H.; Ruiter, D.J.; Van Muijen, G.N.P. Analysis of differential gene expression in human melanocytic tumour lesions by custom made oligonucleotide arrays. Br. J. Cancer 2005, 92, 2249–2261. [Google Scholar] [CrossRef] [PubMed]
- Rathore, M.; Girard, C.; Ohanna, M.; Tichet, M.; Ben Jouira, R.; Garcia, E.; Larbret, F.; Gesson, M.; Audebert, S.; Lacour, J.P.; et al. Cancer cell-derived long pentraxin 3 (PTX3) promotes melanoma migration through a toll-like receptor 4 (TLR4)/NF-κB signaling pathway. Oncogene 2019, 38, 5873–5889. [Google Scholar] [CrossRef] [PubMed]
- Leonard, C.J.; Aravind, L.; Koonin, E.V. Novel families of putative protein kinases in bacteria and archaea: Evolution of the “eukaryotic” protein kinase superfamily. Genome Res. 1998, 8, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Lagier-Tourenne, C.; Tazir, M.; López, L.C.; Quinzii, C.M.; Assoum, M.; Drouot, N.; Busso, C.; Makri, S.; Ali-Pacha, L.; Benhassine, T.; et al. ADCK3, an Ancestral Kinase, Is Mutated in a Form of Recessive Ataxia Associated with Coenzyme Q 10 Deficiency. Am. J. Hum. Genet. 2008, 82, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Brough, R.; Frankum, J.R.; Sims, D.; Mackay, A.; Mendes-pereira, A.M.; Bajrami, I.; Costa-cabral, S.; Rafiq, R.; Ahmad, A.S.; Antonietta, M.; et al. Functional Viability Profiles of Breast Cancer. Cancer Discov. 2011, 1, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, B.; Jia, Z. Preparation and characterization of human ADCK3, a putative atypical kinase. Protein Expr. Purif. 2015, 108, 13–17. [Google Scholar] [CrossRef]
- Goto, L.; Mutsuko, Y.; Ibusuki, Y.; Yamamoto, Y.; Fujiki, Y.; Tomiguchi, M. Therapeutic predictors of neoadjuvant endocrine therapy response in estrogen receptor-positive breast cancer with reference to optimal gene expression profiling. Breast Cancer Res. Treat. 2018, 172, 353–362. [Google Scholar] [CrossRef]
- Kakiuchi, T.; Ohtsuka, Y.; Sato, T.; Nakayama, A.; Jinnouchi, K.; Oka, M. Association between Crohn’s disease and AarF domain-containing kinase 4 glomerulopathy. Clin. J. Gastroenterol. 2019, 12, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Fonseca, L.; Schaefer, J.; Navas-Enamorado, I.; Santos-Ocaña, C.; Hernández-Camacho, J.D.; Guerra, I.; Cascajo, M.V.; Sánchez-Cuesta, A.; Horvath, Z.; Siendones, E.; et al. ADCK2 Haploinsufficiency Reduces Mitochondrial Lipid Oxidation and Causes Myopathy Associated with CoQ Deficiency. J. Clin. Med. 2019, 8, 1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egashira, S.; Jinnin, M.; Makino, K.; Ajino, M.; Shimozono, N.; Okamoto, S.; Tazaki, Y.; Hirano, A. Recurrent Fusion Gene ADCK4-NUMBL in Cutaneous Squamous Cell Carcinoma Mediates Cell Proliferation. J. Investig. Dermatol. 2019, 139, 954–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorns, E.; Lord, C.J.; Grigoriadis, A.; McDonald, S.; Fenwick, K.; MacKay, A.; Mein, C.A.; Natrajan, R.; Savage, K.; Tamber, N.; et al. Integrated functional, gene expression and genomic analysis for the identification of cancer targets. PLoS ONE 2009, 4, e5120. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Li, G.; Wang, P.; Li, X.; Lai, F.; Luo, R.; Liu, B.; Lin, J. aarF domain containing kinase 5 gene promotes invasion and migration of lung cancer cells through ADCK5-SOX9-PTTG1 pathway. Exp. Cell Res. 2020, 392, 112002. [Google Scholar] [CrossRef] [PubMed]
- Wiedemeyer, W.R.; Dunn, I.F.; Quayle, S.N.; Zhang, J.; Chheda, M.G.; Dunn, G.P.; Zhuang, L.; Rosenbluh, J.; Chen, S.; Xiao, Y.; et al. Pattern of retinoblastoma pathway inactivation dictates response to CDK4/6 inhibition in GBM. Proc. Natl. Acad. Sci. USA 2010, 107, 11501–11506. [Google Scholar] [CrossRef] [Green Version]
- Brito, C.; Sousa, S. Non-Muscle Myosin 2A (NM2A): Structure, Regulation and Function. Cells 2020, 9, 1590. [Google Scholar] [CrossRef]
- Li, Y.R.; Yang, W.X. Myosins as fundamental components during tumorigenesis: Diverse and indispensable. Oncotarget 2016, 7, 46785–46812. [Google Scholar] [CrossRef] [Green Version]
- Sellers, J.R.; Heissler, S.M. Nonmuscle myosin-2 isoforms. Curr. Biol. 2019, 29, R275–R278. [Google Scholar] [CrossRef]
- Chaves, L.D.; Ostroski, B.J.; Reed, K.M. Myosin light chain genes in the turkey (Meleagris gallopavo). Cytogenet. Genome Res. 2003, 102, 340–346. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K. Limma: Linear models for microarray data. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer: New York, NY, USA, 2005. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; The R Foundation: Vienna, Austria, 2019; Available online: https://www.r-project.org/ (accessed on 3 November 2021).
- Schoolmeesters, A.; Brown, D.D.; Fedorov, Y. Kinome-Wide Functional Genomics Screen Reveals a Novel Mechanism of TNFa-Induced Nuclear Accumulation of the HIF-1a Transcription Factor in Cancer Cells. PLoS ONE 2012, 7, e31270. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.J.; Selfors, L.M.; Bui, J.; Reynolds, A.; Leake, D.; Khvorova, A.; Brugge, J.S. Identification of genes that regulate epithelial cell migration using an siRNA screening approach. Nat. Cell Biol. 2008, 10, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Montagnani, V.; Benelli, M.; Apollo, A.; Pescucci, C.; Licastro, D.; Urso, C.; Gerlini, G.; Borgognoni, L.; Luzzatto, L.; Stecca, B. Thin and thick primary cutaneous melanomas reveal distinct patterns of somatic copy number alterations. Oncotarget 2016, 7, 30365–30378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penning, A.J.; Al-Ibraheemi, A.; Michal, M.; Larsen, B.T.; Cho, S.J.; Lockwood, C.M.; Paulson, V.A.; Liu, Y.J.; Plank, L.; Fritchie, K.; et al. Novel BRAF gene fusions and activating point mutations in spindle cell sarcomas with histologic overlap with infantile fibrosarcoma. Mod. Pathol. 2021, 34, 1530–1540. [Google Scholar] [CrossRef]
- Cheng, P.F.; Shakhova, O.; Widmer, D.S.; Eichhoff, O.M.; Zingg, D.; Frommel, S.C.; Belloni, B.; Raaijmakers, M.I.G.; Goldinger, S.M.; Santoro, R.; et al. Methylation-dependent SOX9 expression mediates invasion in human melanoma cells and is a negative prognostic factor in advanced melanoma. Genome Biol. 2015, 16, 42. [Google Scholar] [CrossRef] [Green Version]
- Heissler, S.M.; Manstein, D.J. Nonmuscle myosin-2: Mix and match. Cell. Mol. Life Sci. 2013, 70, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Middelbeek, J.; Vrenken, K.; Visser, D.; Lasonder, E.; Koster, J.; Jalink, K.; Clark, K.; van Leeuwen, F.N. The TRPM7 interactome defines a cytoskeletal complex linked to neuroblastoma progression. Eur. J. Cell Biol. 2016, 95, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.; Shen, L.; Wang, H.; Zhang, L. Classify multicategory outcome in patients with lung adenocarcinoma using clinical, transcriptomic and clinico-transcriptomic data: Machine learning versus multinomial models. Am. J. Cancer Res. 2020, 10, 4624–4639. [Google Scholar]
- Schramek, D.; Sendoel, A.; Segal, J.P.; Beronja, S.; Heller, E.; Oristian, D.; Reva, B.; Fuchs, E. Direct in vivo RNAi screen unveils myosin IIa as a tumor suppressor of squamous cell carcinomas. Science 2014, 343, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Shutova, M.S.; Svitkina, T.M. Common and Specific Functions of Nonmuscle Myosin II Paralogs in Cells. Biochemistry 2018, 83, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Han, C.; Jin, S.; Lee, B.; Choi, H.; Kwon, J.T.; Kim, D.; Kim, J.; Lifirsu, E.; Park, W.J.; et al. Myosin regulatory light chains are required to maintain the stability of myosin II and cellular integrity. Biochem. J. 2011, 434, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vierthaler, M.; Sun, Q.; Wang, Y.; Steinfass, T.; Poelchen, J.; Hielscher, T.; Novak, D.; Umansky, V.; Utikal, J. ADCK2 Knockdown Affects the Migration of Melanoma Cells via MYL6. Cancers 2022, 14, 1071. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14041071
Vierthaler M, Sun Q, Wang Y, Steinfass T, Poelchen J, Hielscher T, Novak D, Umansky V, Utikal J. ADCK2 Knockdown Affects the Migration of Melanoma Cells via MYL6. Cancers. 2022; 14(4):1071. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14041071
Chicago/Turabian StyleVierthaler, Marlene, Qian Sun, Yiman Wang, Tamara Steinfass, Juliane Poelchen, Thomas Hielscher, Daniel Novak, Viktor Umansky, and Jochen Utikal. 2022. "ADCK2 Knockdown Affects the Migration of Melanoma Cells via MYL6" Cancers 14, no. 4: 1071. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14041071