
Identification of a Steroid Hormone-Associated Gene Signature Predicting the Prognosis of Prostate Cancer through an Integrative Bioinformatics Analysis

 , , , , , , , and
, , , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection and Preparation
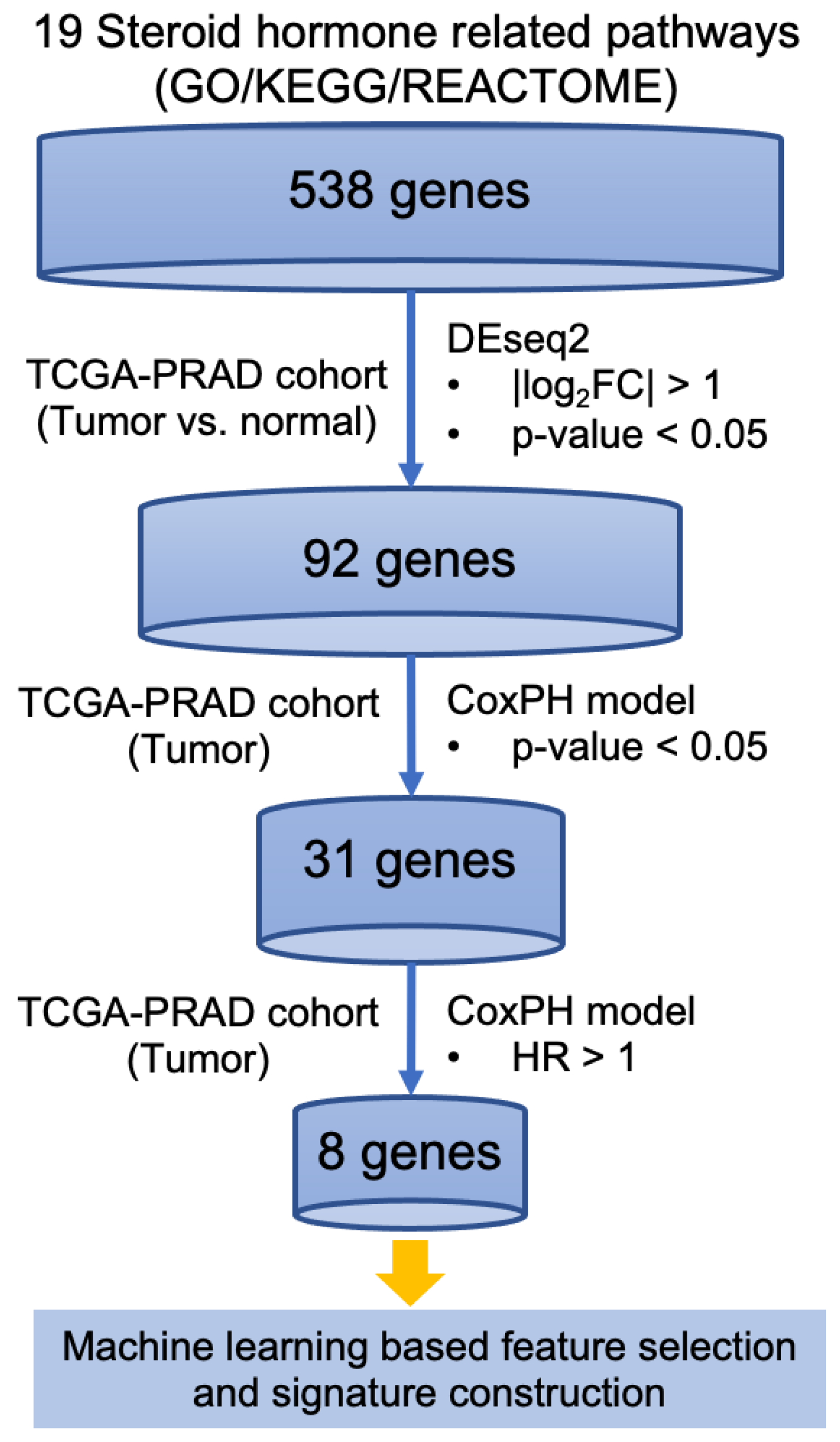
2.2. Steroid Hormone-Related Gene Selection
2.3. Differentially Expression Analysis
2.4. Survival Analysis
2.5. Feature Selection and Signature Construction
2.6. Independent Datasets Validation
2.7. Functional Annotation
3. Results
3.1. Identification of Steroid Hormone Genes Associated with Disease Progression in Prostate Cancer
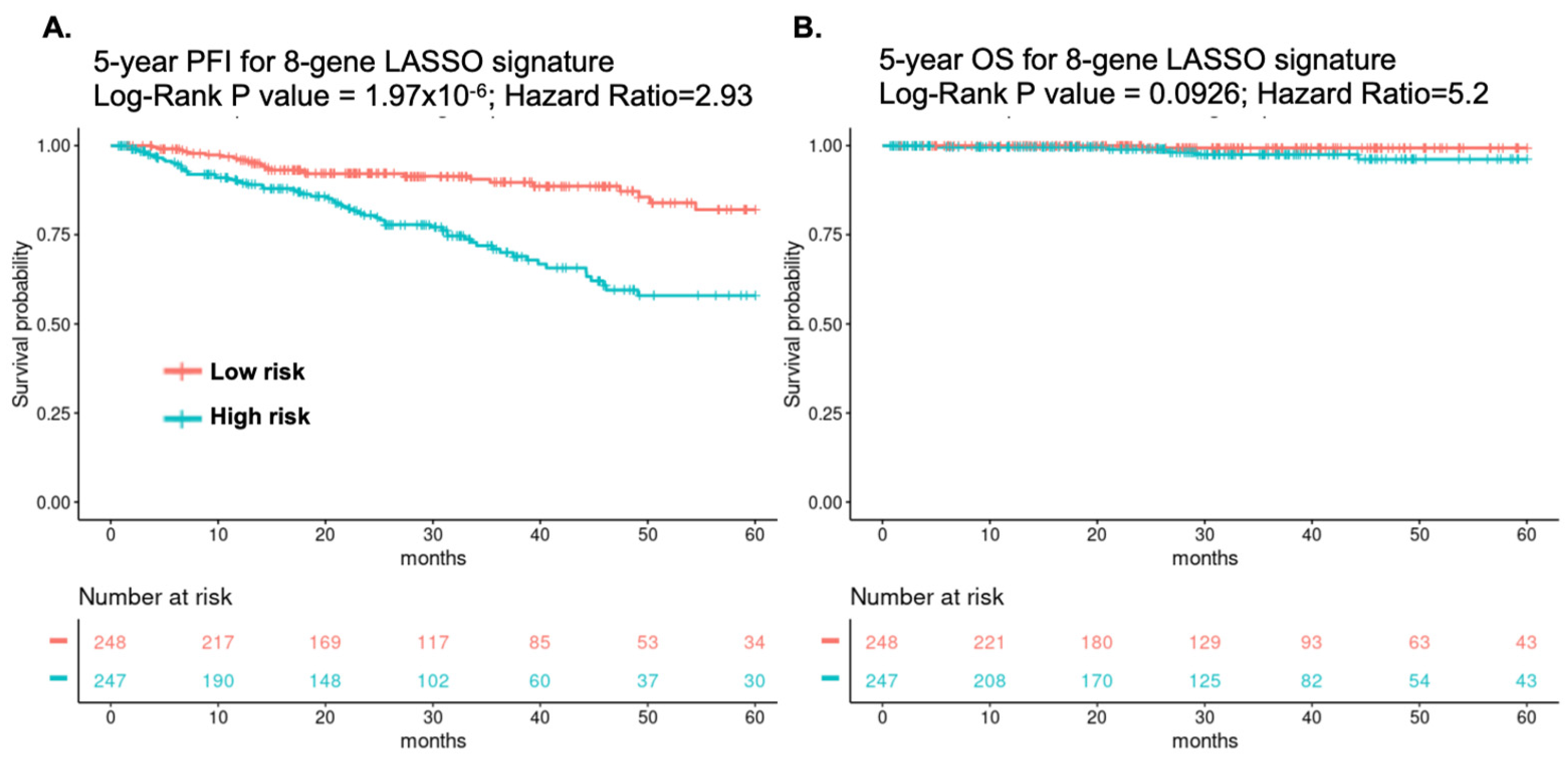
3.2. Identification of an Eight-Gene Signature Predicting PC Survival
3.3. Multivariate Cox Regression Analysis with Clinical Variables
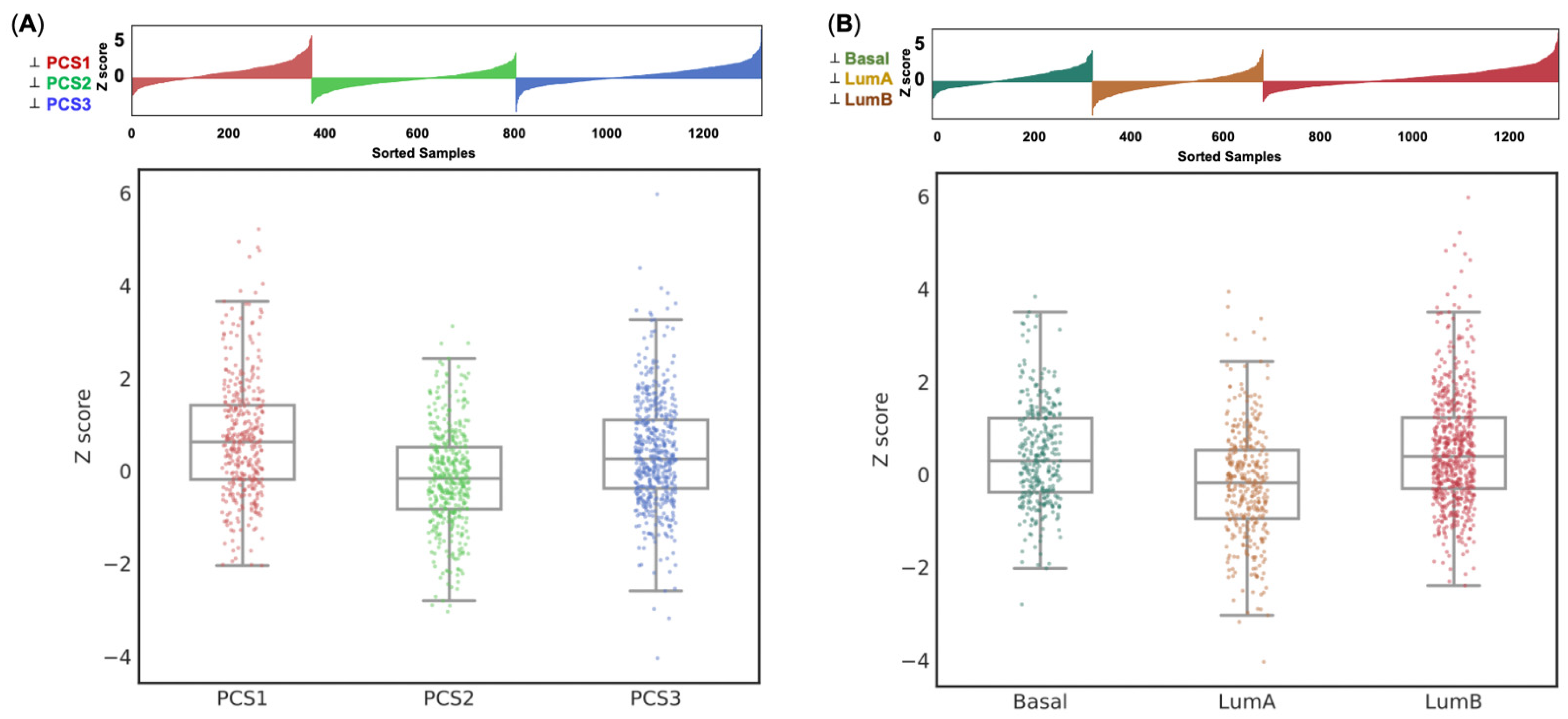
3.4. Expression of the Eight-Gene Panel Based on External PC Cohort’s Validation
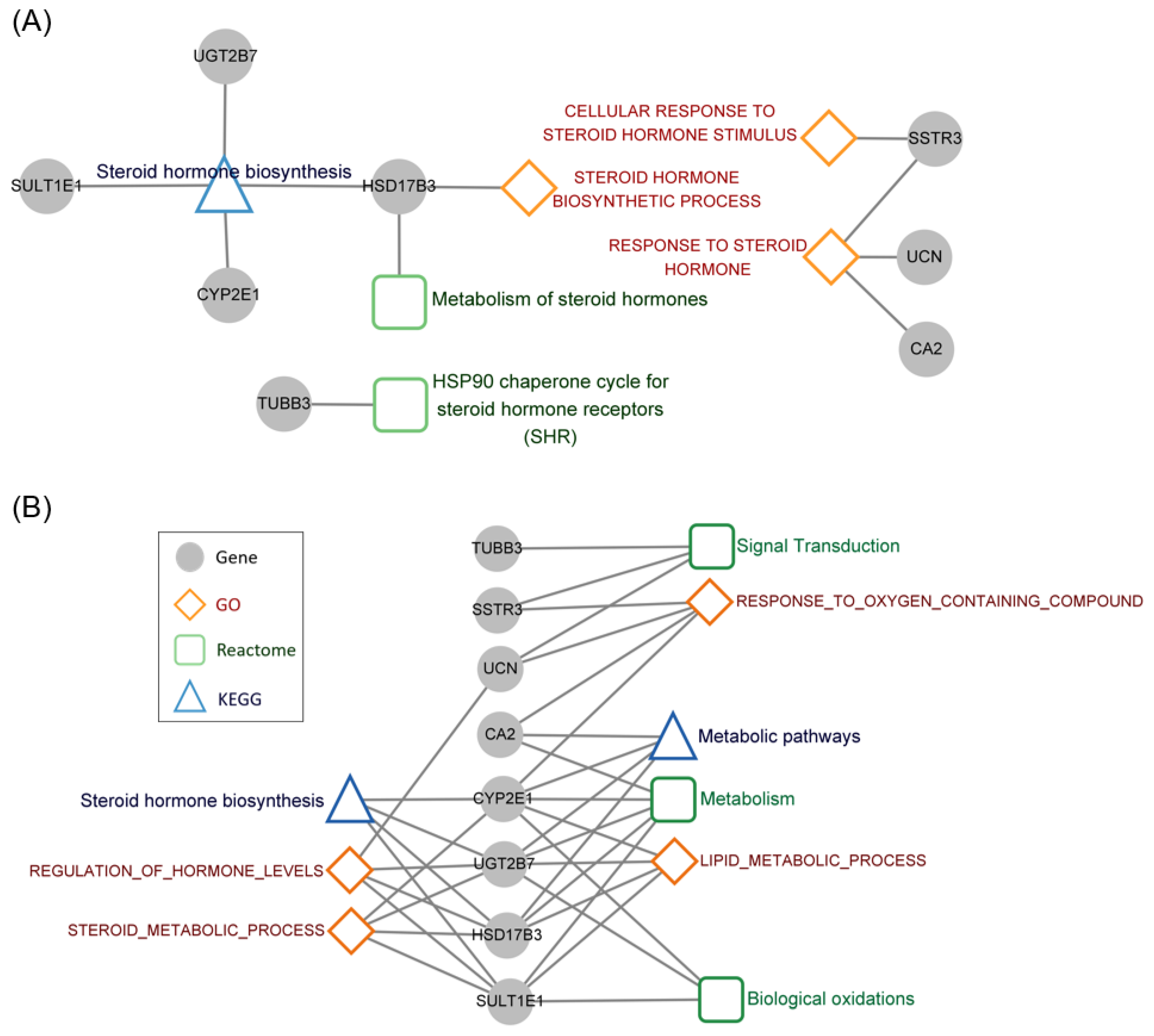
3.5. Functional Annotation of the Steroid Hormone Genes Associated with Prognosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bosland, M.C. The role of steroid hormones in prostate carcinogenesis. J. Natl. Cancer Inst. Monogr. 2000, 27, 39–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, A.M.; Pramanik, R.; Nicholson, L.J.; Dew, T.K.; Martin, F.L.; Muir, G.H.; Morris, J.D. Ras-MEK-ERK signaling cascade regulates androgen receptor element-inducible gene transcription and DNA synthesis in prostate cancer cells. Int. J. Cancer 2007, 121, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Network, N.C.C. Prostate Cancer (Version 2.2022). Available online: https://www.nccn.org/professionals/physician_gls/pdf/prostate.pdf (accessed on 15 January 2022).
- De Bono, J.S.; Logothetis, C.J.; Molina, A.; Fizazi, K.; North, S.; Chu, L.; Chi, K.N.; Jones, R.J.; Goodman, O.B., Jr.; Saad, F.; et al. Abiraterone and increased survival in metastatic prostate cancer. N. Engl. J. Med. 2011, 364, 1995–2005. [Google Scholar] [CrossRef]
- Smith, M.R.; Saad, F.; Chowdhury, S.; Oudard, S.; Hadaschik, B.A.; Graff, J.N.; Olmos, D.; Mainwaring, P.N.; Lee, J.Y.; Uemura, H.; et al. Apalutamide treatment and metastasis-free survival in prostate cancer. N. Engl. J. Med. 2018, 378, 1408–1418. [Google Scholar] [CrossRef]
- Scher, H.I.; Fizazi, K.; Saad, F.; Taplin, M.E.; Sternberg, C.N.; Miller, K.; de Wit, R.; Mulders, P.; Chi, K.N.; Shore, N.D.; et al. Increased survival with enzalutamide in prostate cancer after chemotherapy. N. Engl. J. Med. 2012, 367, 1187–1197. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, V.E.; Roviello, G.; D’Angelo, A.; Casadei, C.; De Giorgi, U.; Giorgione, R. Darolutamide in hormone-sensitive and castration-resistant prostate cancer. Expert Rev. Clin. Pharmacol. 2021, 14, 535–544. [Google Scholar] [CrossRef]
- Fizazi, K.; Tran, N.; Fein, L.; Matsubara, N.; Rodriguez-Antolin, A.; Alekseev, B.Y.; Ozguroglu, M.; Ye, D.; Feyerabend, S.; Protheroe, A.; et al. Abiraterone plus prednisone in metastatic, castration-sensitive prostate cancer. N. Engl. J. Med. 2017, 377, 352–360. [Google Scholar] [CrossRef]
- Robinson, D.; Van Allen, E.M.; Wu, Y.M.; Schultz, N.; Lonigro, R.J.; Mosquera, J.M.; Montgomery, B.; Taplin, M.E.; Pritchard, C.C.; Attard, G.; et al. Integrative clinical genomics of advanced prostate cancer. Cell 2015, 161, 1215–1228. [Google Scholar] [CrossRef] [Green Version]
- International Cancer Genome Consortium; Hudson, T.J.; Anderson, W.; Artez, A.; Barker, A.D.; Bell, C.; Bernabe, R.R.; Bhan, M.K.; Calvo, F.; Eerola, I.; et al. International network of cancer genome projects. Nature 2010, 464, 993–998. [Google Scholar]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An integrated TCGA pan-cancer clinical data resource to drive high-quality survival outcome analytics. Cell 2018, 173, 400–416.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Genome Atlas Research Network. The molecular taxonomy of primary prostate cancer. Cell 2015, 163, 1011–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bono, J.; Mateo, J.; Fizazi, K.; Saad, F.; Shore, N.; Sandhu, S.; Chi, K.N.; Sartor, O.; Agarwal, N.; Olmos, D.; et al. Olaparib for metastatic castration-resistant prostate cancer. N. Engl. J. Med. 2020, 382, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Kourou, K.; Exarchos, T.P.; Exarchos, K.P.; Karamouzis, M.V.; Fotiadis, D.I. Machine learning applications in cancer prognosis and prediction. Comput. Struct. Biotechnol. J. 2015, 13, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Tataru, O.S.; Vartolomei, M.D.; Rassweiler, J.J.; Virgil, O.; Lucarelli, G.; Porpiglia, F.; Amparore, D.; Manfredi, M.; Carrieri, G.; Falagario, U.; et al. Artificial intelligence and machine learning in prostate cancer patient management-current trends and future perspectives. Diagnostics 2021, 11, 354. [Google Scholar] [CrossRef]
- Cheng, W.C.; Chung, I.F.; Chen, C.Y.; Sun, H.J.; Fen, J.J.; Tang, W.C.; Chang, T.Y.; Wong, T.T.; Wang, H.W. DriverDB: An exome sequencing database for cancer driver gene identification. Nucl. Acids Res. 2014, 42, D1048–D1054. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.F.; Chen, C.Y.; Su, S.C.; Li, C.Y.; Wu, K.J.; Wang, H.W.; Cheng, W.C. DriverDBv2: A database for human cancer driver gene research. Nucl. Acids Res. 2016, 44, D975–D979. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.H.; Shen, P.C.; Chen, C.Y.; Hsu, A.N.; Cho, Y.C.; Lai, Y.L.; Chen, F.H.; Li, C.Y.; Wang, S.C.; Chen, M.; et al. DriverDBv3: A multi-omics database for cancer driver gene research. Nucl. Acids Res. 2019, 48, D863–D870. [Google Scholar] [CrossRef] [Green Version]
- Carbon, S.; Ireland, A.; Mungall, C.J.; Shu, S.; Marshall, B.; Lewis, S.; The AmiGO Hub; The Web Presence Working Group. AmiGO: Online access to ontology and annotation data. Bioinformatics 2009, 25, 288–289. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucl. Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Therneau, T.M.; Lumley, T. Package ‘survival’. R Top Doc. 2015, 128, 112. [Google Scholar]
- Simon, N.; Friedman, J.H.; Hastie, T.; Tibshirani, R. Regularization paths for Cox’s proportional hazards model via coordinate descent. J. Stat. Softw. 2011, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.H.; Hastie, T.; Tibshirani, R. Regularization paths for generalized linear models via coordinate descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, J.; Inazawa, J. Cancer-associated miRNAs and their therapeutic potential. J. Hum. Genet. 2021, 66, 937–945. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Knudsen, B.S.; Erho, N.; Alshalalfa, M.; Takhar, M.; Al-Deen Ashab, H.; Davicioni, E.; Karnes, R.J.; Klein, E.A.; Den, R.B.; et al. Integrated classification of prostate cancer reveals a novel luminal subtype with poor outcome. Cancer Res. 2016, 76, 4948–4958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.G.; Chang, S.L.; Erho, N.; Yu, M.; Lehrer, J.; Alshalalfa, M.; Speers, C.; Cooperberg, M.R.; Kim, W.; Ryan, C.J.; et al. Associations of luminal and basal subtyping of prostate cancer with prognosis and response to androgen deprivation therapy. JAMA Oncol. 2017, 3, 1663–1672. [Google Scholar] [CrossRef]
- Jacob, A.; Raj, R.; Allison, D.B.; Myint, Z.W. Androgen receptor signaling in prostate cancer and therapeutic strategies. Cancers 2021, 13, 5417. [Google Scholar] [CrossRef]
- Krebs, M.; Solimando, A.G.; Kalogirou, C.; Marquardt, A.; Frank, T.; Sokolakis, I.; Hatzichristodoulou, G.; Kneitz, S.; Bargou, R.; Kubler, H.; et al. miR-221-3p regulates VEGFR2 expression in high-risk prostate cancer and represents an escape mechanism from sunitinib in vitro. J. Clin. Med. 2020, 9, 670. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.B.; Mostaghel, E.A.; Vessella, R.; Hess, D.L.; Kalhorn, T.F.; Higano, C.S.; True, L.D.; Nelson, P.S. Maintenance of intratumoral androgens in metastatic prostate cancer: A mechanism for castration-resistant tumor growth. Cancer Res. 2008, 68, 4447–4454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostaghel, E.A.; Zhang, A.; Hernandez, S.; Marck, B.T.; Zhang, X.; Tamae, D.; Biehl, H.E.; Tretiakova, M.; Bartlett, J.; Burns, J.; et al. Contribution of adrenal glands to intratumor androgens and growth of castration-resistant prostate cancer. Clin. Cancer Res. 2019, 25, 426–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzucchelli, R.; Morichetti, D.; Scarpelli, M.; Bono, A.V.; Lopez-Beltran, A.; Cheng, L.; Kirkali, Z.; Montironi, R. Somatostatin receptor subtypes in hormone-refractory (castration-resistant) prostatic carcinoma. Asian J. Androl. 2011, 13, 242–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savelli, G.; Muni, A.; Falchi, R.; Zaniboni, A.; Barbieri, R.; Valmadre, G.; Minari, C.; Casi, C.; Rossini, P. Somatostatin receptors over-expression in castration resistant prostate cancer detected by PET/CT: Preliminary report of in six patients. Ann. Transl. Med. 2015, 3, 145. [Google Scholar] [PubMed]
- Liu, M.; Shi, H.; Yan, J.; Zhang, Y.; Ma, Y.; Le, K.; Li, Z.; Xing, N.; Li, G. Gene polymorphism-related differences in the outcomes of abiraterone for prostate cancer: A systematic overview. Am. J. Cancer Res. 2021, 11, 1873–1894. [Google Scholar] [PubMed]
- Agarwal, N.; Alex, A.B.; Farnham, J.M.; Patel, S.; Gill, D.; Buckley, T.H.; Stephenson, R.A.; Cannon-Albright, L. Inherited variants in SULT1E1 and response to abiraterone acetate by men with metastatic castration refractory prostate cancer. J. Urol. 2016, 196, 1112–1116. [Google Scholar] [CrossRef]
- Sekino, Y.; Han, X.; Babasaki, T.; Miyamoto, S.; Kobatake, K.; Kitano, H.; Ikeda, K.; Goto, K.; Inoue, S.; Hayashi, T.; et al. TUBB3 is associated with PTEN, neuroendocrine differentiation, and castration resistance in prostate cancer. Urol. Oncol. 2021, 39, 368.e1–368.e9. [Google Scholar] [CrossRef]
- Terry, S.; Ploussard, G.; Allory, Y.; Nicolaiew, N.; Boissiere-Michot, F.; Maille, P.; Kheuang, L.; Coppolani, E.; Ali, A.; Bibeau, F.; et al. Increased expression of class III beta-tubulin in castration-resistant human prostate cancer. Br. J. Cancer 2009, 101, 951–956. [Google Scholar] [CrossRef] [Green Version]
- Ploussard, G.; Terry, S.; Maille, P.; Allory, Y.; Sirab, N.; Kheuang, L.; Soyeux, P.; Nicolaiew, N.; Coppolani, E.; Paule, B.; et al. Class III beta-tubulin expression predicts prostate tumor aggressiveness and patient response to docetaxel-based chemotherapy. Cancer Res. 2010, 70, 9253–9264. [Google Scholar] [CrossRef] [Green Version]
- Belanger, A.; Pelletier, G.; Labrie, F.; Barbier, O.; Chouinard, S. Inactivation of androgens by UDP-glucuronosyltransferase enzymes in humans. Trends Endocrinol. Metab. 2003, 14, 473–479. [Google Scholar] [CrossRef]
- Li, H.; Xie, N.; Chen, R.; Verreault, M.; Fazli, L.; Gleave, M.E.; Barbier, O.; Dong, X. UGT2B17 expedites progression of castration-resistant prostate cancers by promoting ligand-independent AR signaling. Cancer Res. 2016, 76, 6701–6711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcuri, F.; Cintorino, M.; Florio, P.; Floccari, F.; Pergola, L.; Romagnoli, R.; Petraglia, F.; Tosi, P.; Teresa Del Vecchio, M. Expression of urocortin mRNA and peptide in the human prostate and in prostatic adenocarcinoma. Prostate 2002, 52, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Tezval, H.; Jurk, S.; Atschekzei, F.; Serth, J.; Kuczyk, M.A.; Merseburger, A.S. The involvement of altered corticotropin releasing factor receptor 2 expression in prostate cancer due to alteration of anti-angiogenic signaling pathways. Prostate 2009, 69, 443–448. [Google Scholar] [CrossRef]
- Ferreira, P.M.; Medeiros, R.; Vasconcelos, A.; Costa, S.; Pinto, D.; Morais, A.; Liveira, J.; Lopes, C. Association between CYP2E1 polymorphisms and susceptibility to prostate cancer. Eur. J. Cancer Prev. 2003, 12, 205–211. [Google Scholar] [CrossRef]
- Korhonen, K.; Parkkila, A.K.; Helen, P.; Valimaki, R.; Pastorekova, S.; Pastorek, J.; Parkkila, S.; Haapasalo, H. Carbonic anhydrases in meningiomas: Association of endothelial carbonic anhydrase II with aggressive tumor features. J. Neurosurg. 2009, 111, 472–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purohit, A.; Woo, L.W.; Chander, S.K.; Newman, S.P.; Ireson, C.; Ho, Y.; Grasso, A.; Leese, M.P.; Potter, B.V.; Reed, M.J. Steroid sulphatase inhibitors for breast cancer therapy. J. Steroid Biochem. Mol. Biol. 2003, 86, 423–432. [Google Scholar] [CrossRef]
- Pettersson, A.; Robinson, D.; Garmo, H.; Holmberg, L.; Stattin, P. Age at diagnosis and prostate cancer treatment and prognosis: A population-based cohort study. Ann. Oncol. 2018, 29, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Prensner, J.R.; Rubin, M.A.; Wei, J.T.; Chinnaiyan, A.M. Beyond PSA: The next generation of prostate cancer biomarkers. Sci. Transl. Med. 2012, 4, 127rv3. [Google Scholar] [CrossRef] [Green Version]
- Wallden, B.; Storhoff, J.; Nielsen, T.; Dowidar, N.; Schaper, C.; Ferree, S.; Liu, S.; Leung, S.; Geiss, G.; Snider, J.; et al. Development and verification of the PAM50-based Prosigna breast cancer gene signature assay. BMC Med. Genom. 2015, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Feng, F.Y.; Thomas, S.; Saad, F.; Gormley, M.; Yu, M.K.; Ricci, D.S.; Rooney, B.; Brookman-May, S.; McCarthy, S.; Olmos, D.; et al. Association of molecular subtypes with differential outcome to apalutamide treatment in nonmetastatic castration-resistant prostate cancer. JAMA Oncol. 2021, 7, 1005–1014. [Google Scholar] [CrossRef]
- Huntley, R.P.; Sawford, T.; Martin, M.J.; O’Donovan, C. Understanding how and why the Gene Ontology and its annotations evolve: The GO within UniProt. Gigascience 2014, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Zong, W.; Lin, C.W.; Fang, Z.; Ma, T.; Lewis, D.A.; Enwright, J.F.; Tseng, G.C. Comparative pathway integrator: A framework of meta-analytic integration of multiple transcriptomic studies for consensual and differential pathway analysis. Genes 2020, 11, 696. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Chung, Y.M.; Sergeeva, O.; Kepe, V.; Berk, M.; Li, J.; Ko, H.K.; Li, Z.; Petro, M.; DiFilippo, F.P.; et al. Loss of dihydrotestosterone-inactivation activity promotes prostate cancer castration resistance detectable by functional imaging. J. Biol. Chem. 2018, 293, 17829–17837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sten, T.; Kurkela, M.; Kuuranne, T.; Leinonen, A.; Finel, M. UDP-glucuronosyltransferases in conjugation of 5alpha- and 5beta-androstane steroids. Drug Metab. Dispos. 2009, 37, 2221–2227. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Differential Expression Analysis | Survival Analysis (PFI) | ||||
|---|---|---|---|---|---|
| Gene | Log2 Fold Change | Adjusted p-Value | HR | CI95 | p-Value |
| CA2 | −4.48699 | 2.20 × 10−78 | 2.14 | 1.36–3.37 | 0.001038 |
| CYP2E1 | −1.88521 | 1.31 × 10−25 | 1.55 | 1.01–2.38 | 0.043481 |
| HSD17B3 | 1.32350 | 4.89 × 10−11 | 2.19 | 1.40–3.40 | 0.000527 |
| SSTR3 | −1.21147 | 2.44 × 10−5 | 1.83 | 1.18–2.83 | 0.006554 |
| SULT1E1 | −1.23635 | 9.17 × 10−6 | 1.94 | 1.24–3.01 | 0.003371 |
| TUBB3 | 1.32113 | 2.14 × 10−10 | 2.27 | 1.45–3.54 | 0.000319 |
| UCN | 2.16915 | 4.31 × 10−41 | 1.94 | 1.24–3.01 | 0.003137 |
| UGT2B7 | −5.67669 | 1.25 × 10−54 | 1.64 | 1.07–2.52 | 0.023815 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, Y.-L.; Liu, C.-H.; Wang, S.-C.; Huang, S.-P.; Cho, Y.-C.; Bao, B.-Y.; Su, C.-C.; Yeh, H.-C.; Lee, C.-H.; Teng, P.-C.; et al. Identification of a Steroid Hormone-Associated Gene Signature Predicting the Prognosis of Prostate Cancer through an Integrative Bioinformatics Analysis. Cancers 2022, 14, 1565. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14061565
Lai Y-L, Liu C-H, Wang S-C, Huang S-P, Cho Y-C, Bao B-Y, Su C-C, Yeh H-C, Lee C-H, Teng P-C, et al. Identification of a Steroid Hormone-Associated Gene Signature Predicting the Prognosis of Prostate Cancer through an Integrative Bioinformatics Analysis. Cancers. 2022; 14(6):1565. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14061565
Chicago/Turabian StyleLai, Yo-Liang, Chia-Hsin Liu, Shu-Chi Wang, Shu-Pin Huang, Yi-Chun Cho, Bo-Ying Bao, Chia-Cheng Su, Hsin-Chih Yeh, Cheng-Hsueh Lee, Pai-Chi Teng, and et al. 2022. "Identification of a Steroid Hormone-Associated Gene Signature Predicting the Prognosis of Prostate Cancer through an Integrative Bioinformatics Analysis" Cancers 14, no. 6: 1565. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14061565