
Immune Landscape and an RBM38-Associated Immune Prognostic Model with Laboratory Verification in Malignant Melanoma


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. RBM38 Expression in Different Datasets
2.2. Survival Prognosis and Genetic Alteration of RBM38
2.3. Cell Culture and Transfection
2.4. Real-Time Quantitative RT-PCR (qRT-PCR)
2.5. CCK-8 and Cell Colony Assay
2.6. Xenograft Tumor in Nude Mice
2.7. Transwell Cell Migration and Invasion Experiment
2.8. Immunohistochemical Staining
2.9. RNA Sequencing Analysis
2.10. GO and KEGG Analyses
2.11. Selection and Analysis of Immune-Related Differential Expressed Genes
2.12. Construction and Validation of Prognostic Models
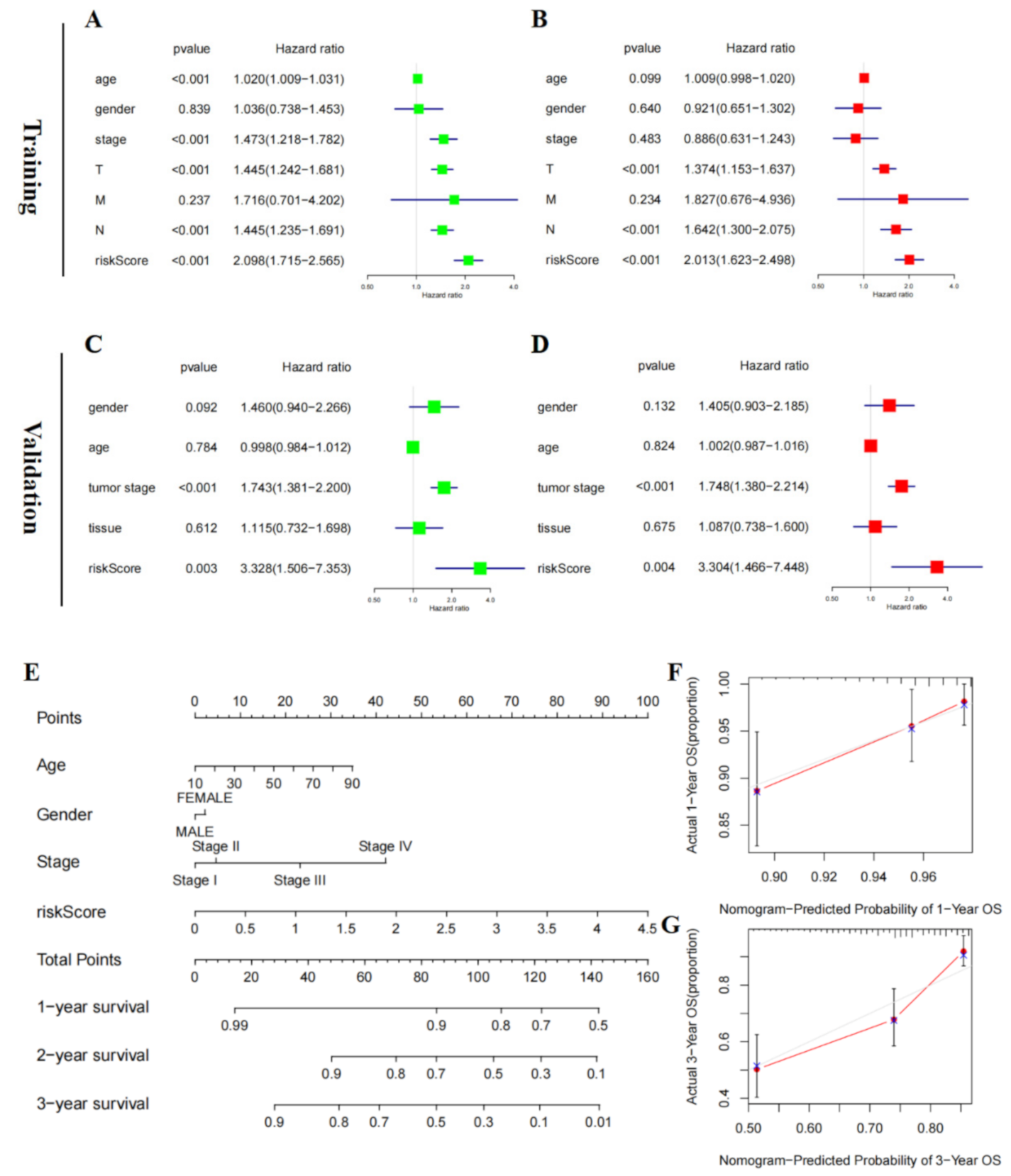
2.13. Independent Prognostic Ability of the Five-Gene Signature
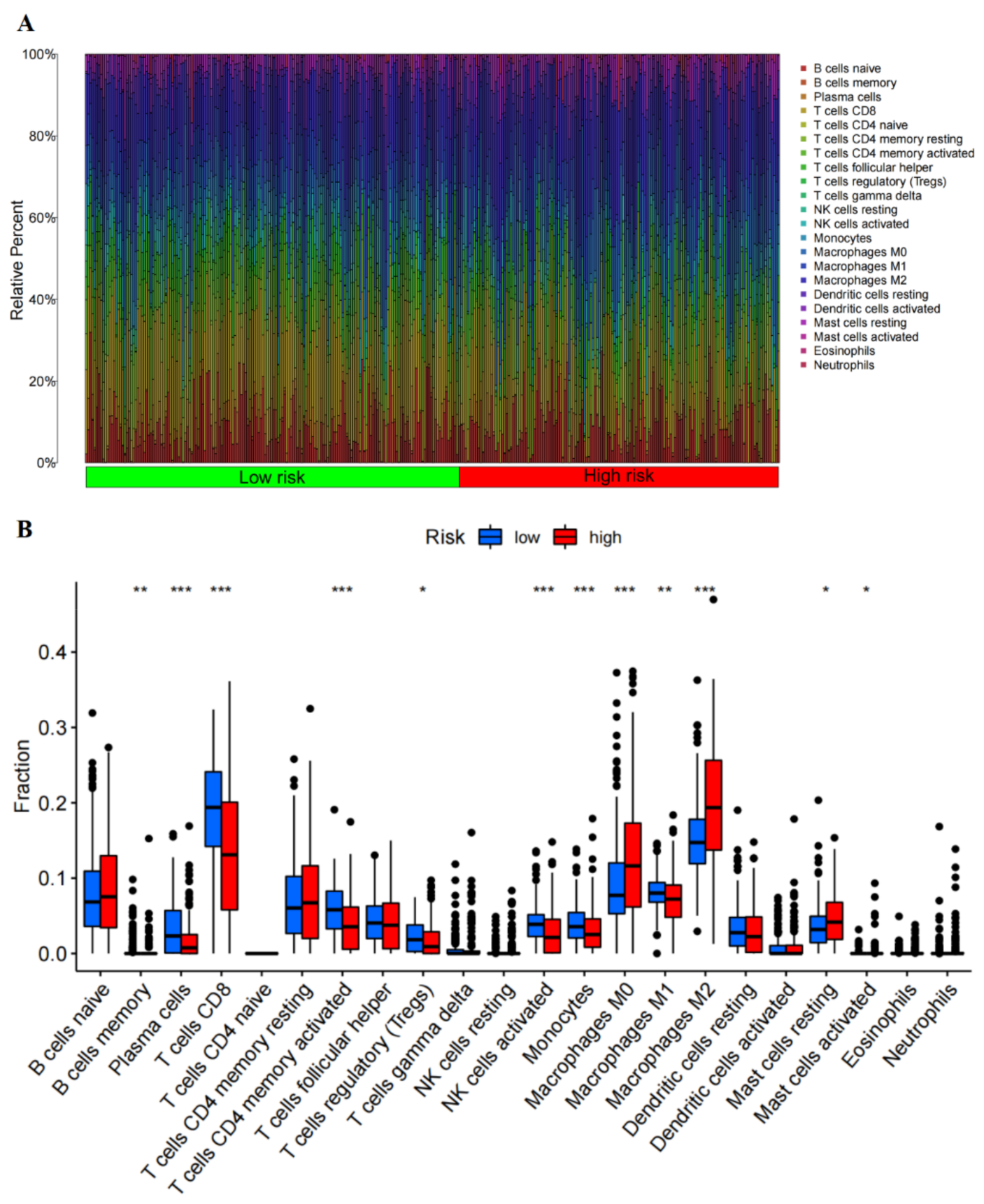
2.14. Immune Infiltration Analysis
2.15. Immunophenoscore
2.16. Statistical Analysis
3. Results
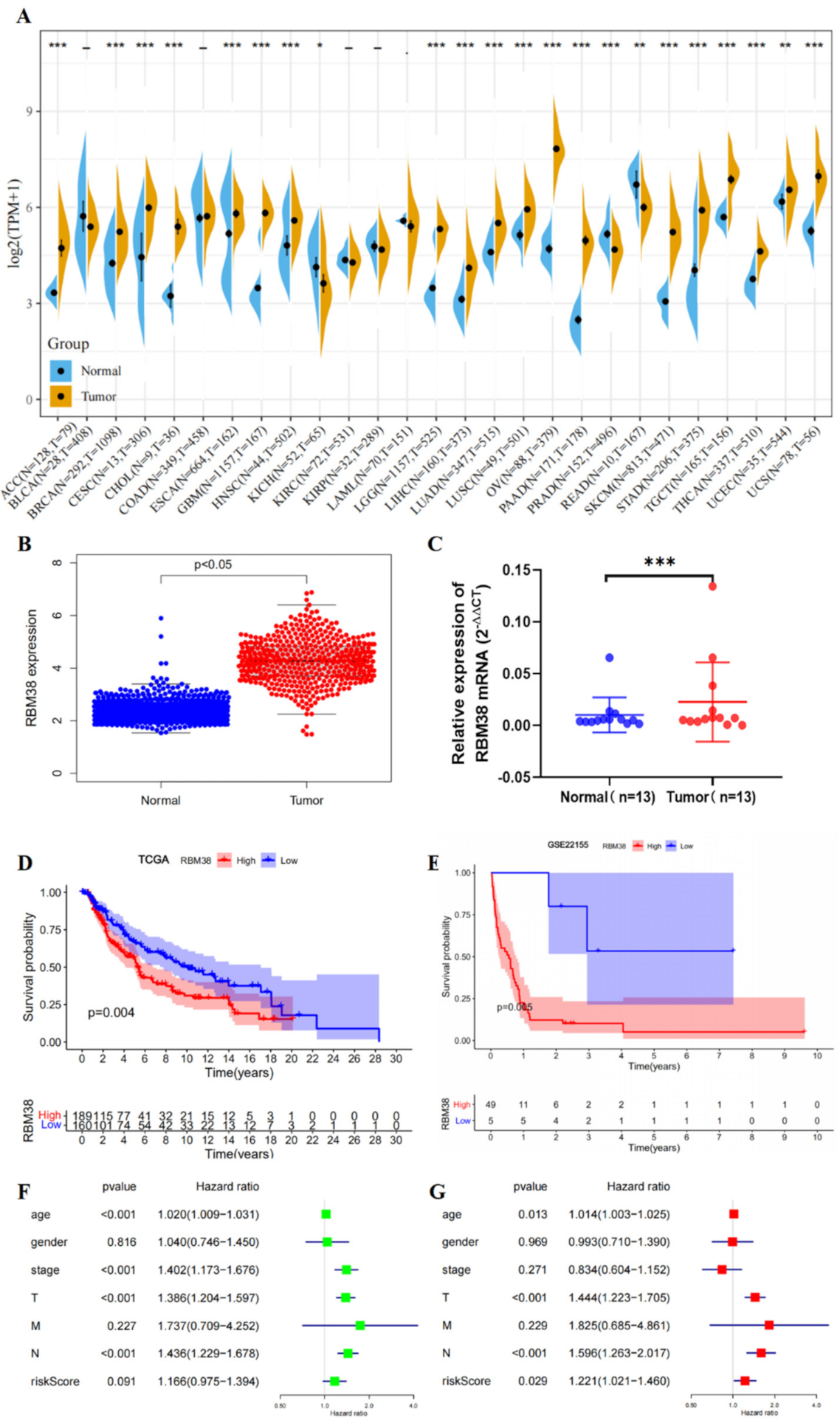
3.1. RBM38 Expression, Alteration, and Prognosis Analysis
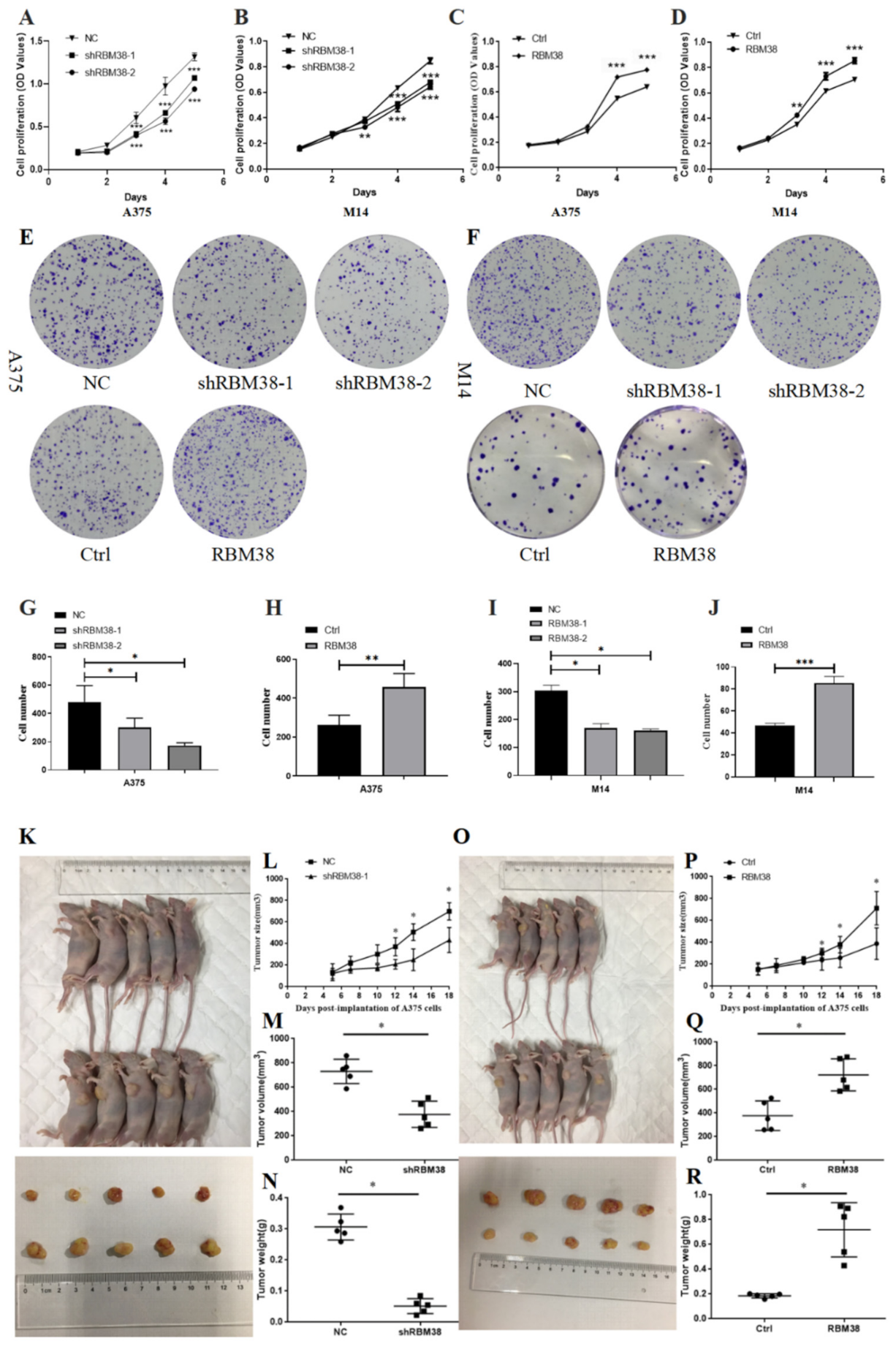
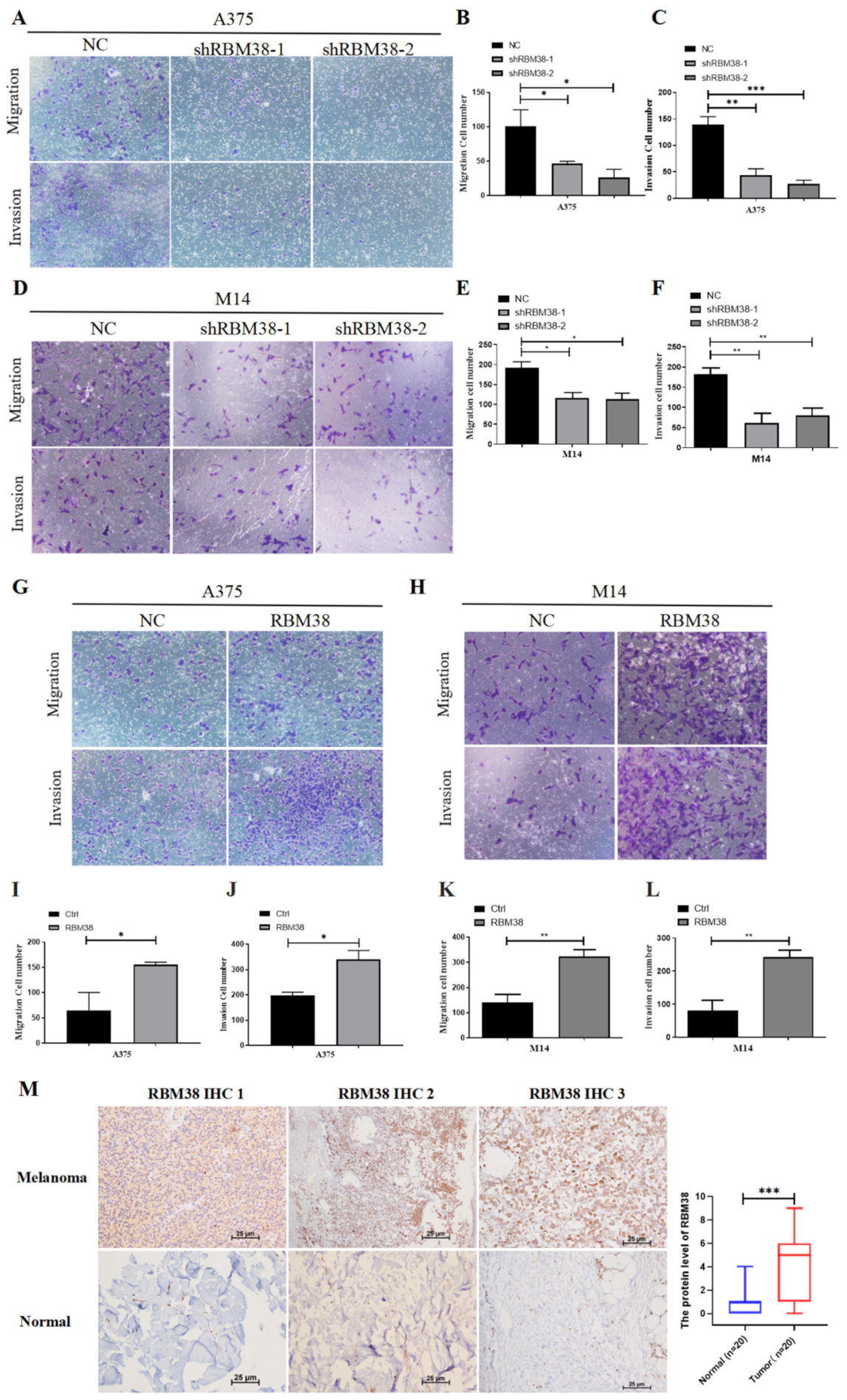
3.2. RBM38 Promotes Melanoma Cell Proliferation, Migration, and Invasion and Influences Immune Cell Infiltration
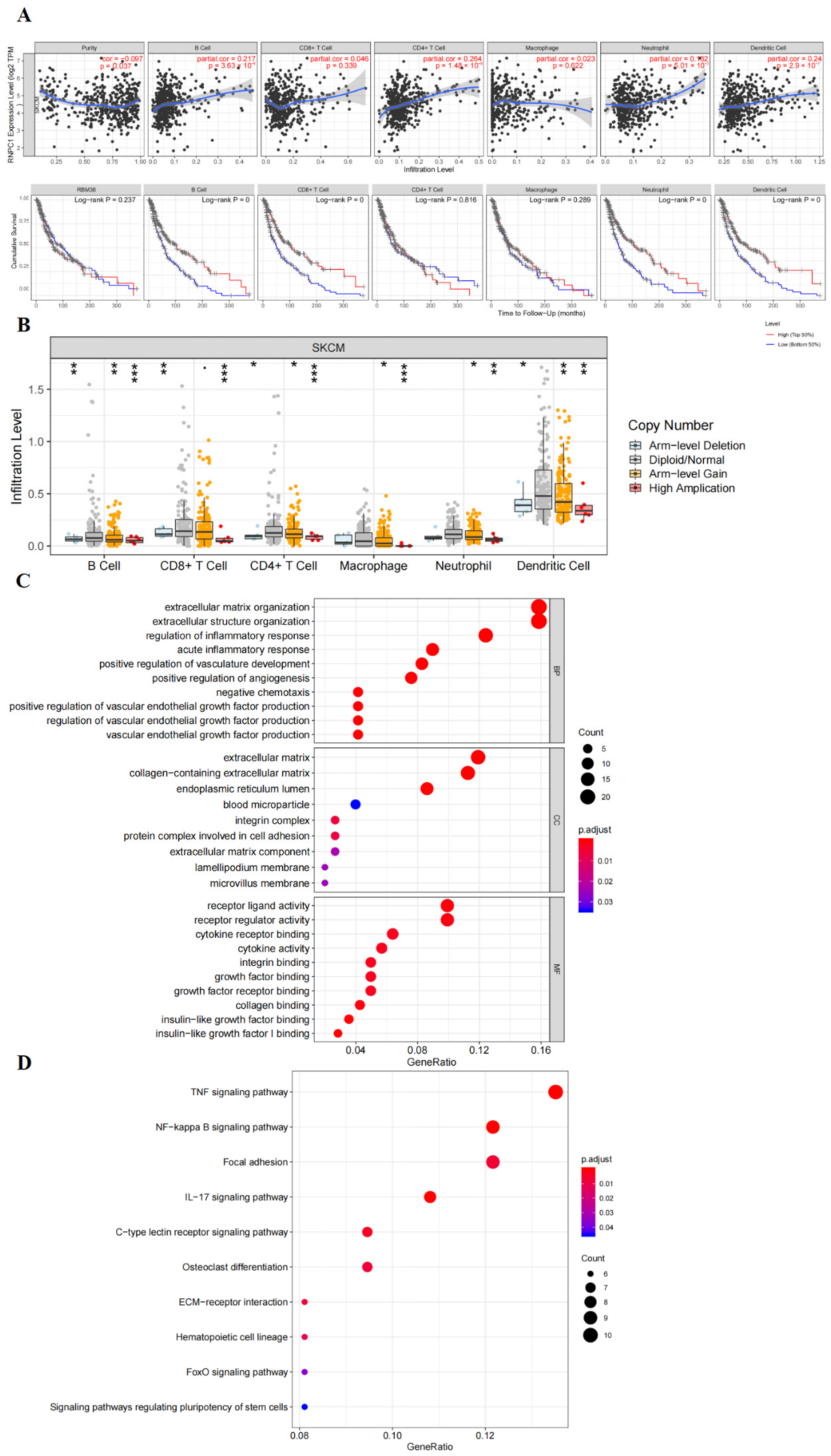
3.3. RBM38 Influences Immune Cell Infiltration and Promotes the Proliferation and Migration by Immune Pathway
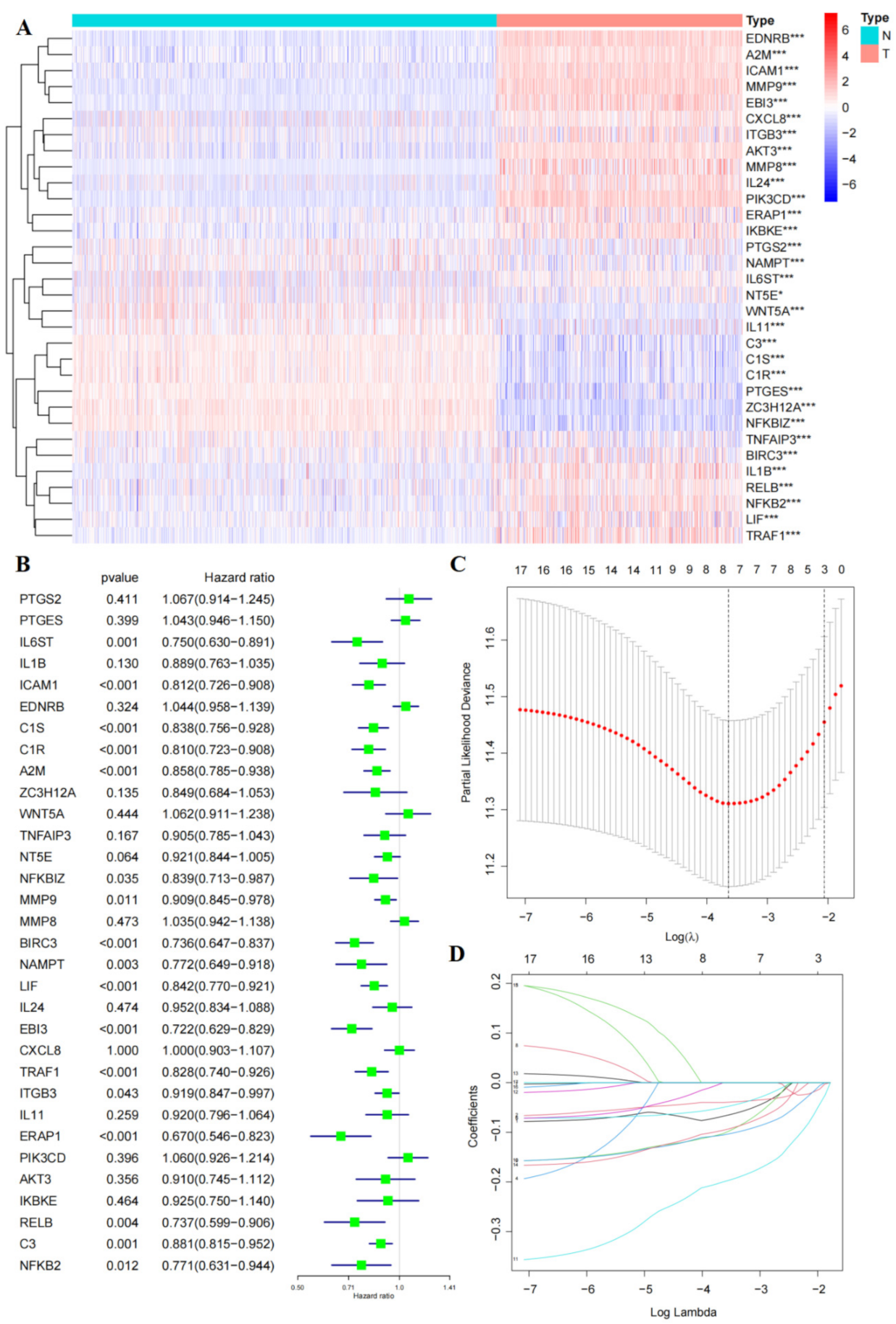
3.4. Construct a Five-Gene Prognostic Signature
3.5. Risk Score of RAIPM Is an Independent Prognostic Factor
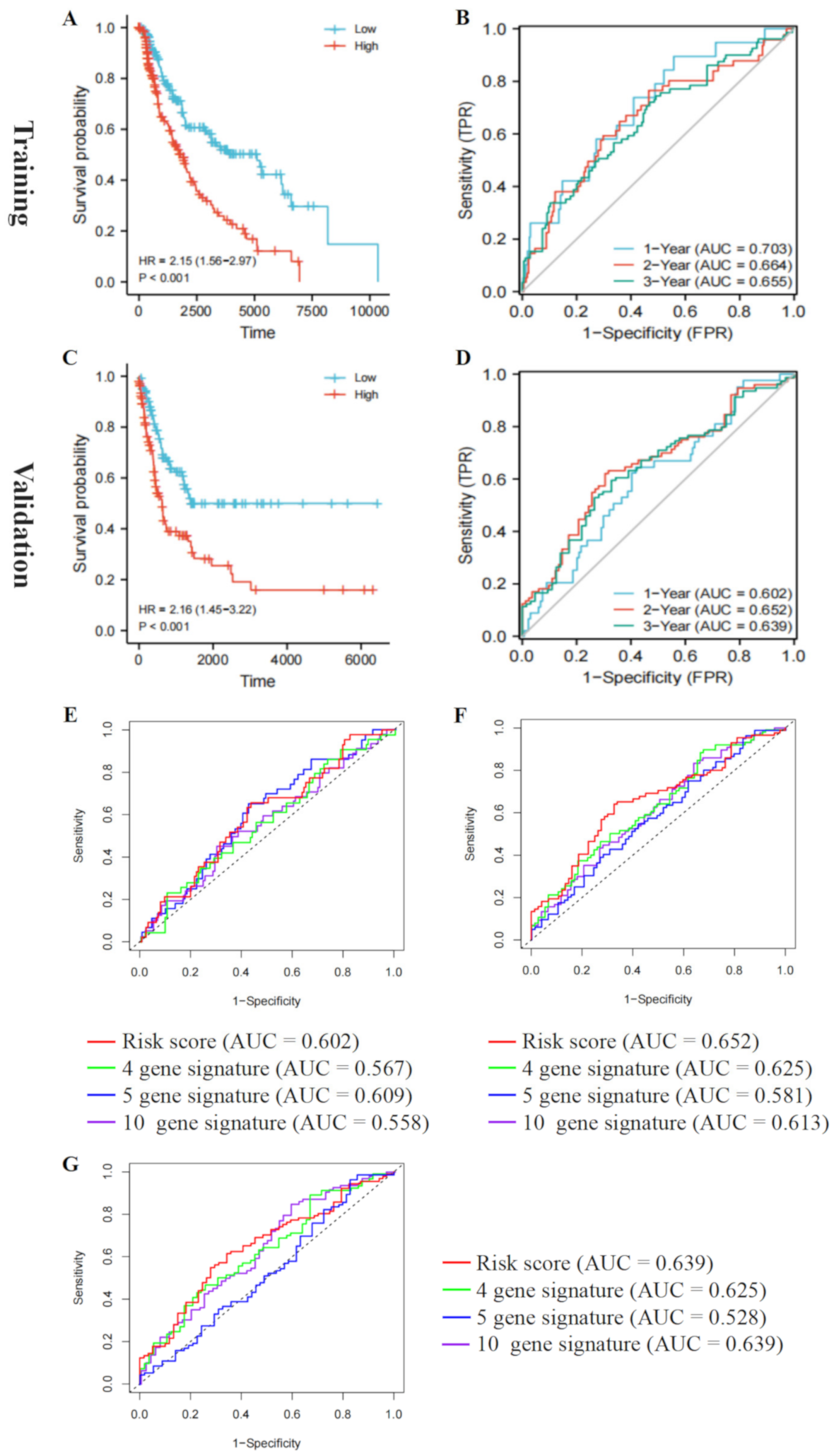
3.6. Validate the Prognostic Value of the Five-Gene Risk Signature
3.7. Association between Immunosuppressive Molecules and Risk Scores in This Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Balch, C.; Gershenwald, J.; Soong, S.; Thompson, J.; Atkins, M.; Byrd, D.; Buzaid, A.; Cochran, A.; Coit, D.; Ding, S.; et al. Final version of 2009 AJCC melanoma staging and classification. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2009, 27, 6199–6206. [Google Scholar] [CrossRef] [Green Version]
- Saranga-Perry, V.; Ambe, C.; Zager, J.; Kudchadkar, R. Recent developments in the medical and surgical treatment of melanoma. CA Cancer J. Clin. 2014, 64, 171–185. [Google Scholar] [CrossRef]
- Leach, D.; Krummel, M.; Allison, J. Enhancement of antitumor immunity by CTLA-4 blockade. Science 1996, 271, 1734–1736. [Google Scholar] [CrossRef] [Green Version]
- Hodi, F.; O'Day, S.; McDermott, D.; Weber, R.; Sosman, J.; Haanen, J.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.; et al. Improved survival with ipilimumab in patients with metastatic melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef]
- Wolchok, J.; Kluger, H.; Callahan, M.; Postow, M.; Rizvi, N.; Lesokhin, A.; Segal, N.; Ariyan, C.; Gordon, R.; Reed, K.; et al. Nivolumab plus ipilimumab in advanced melanoma. N. Engl. J. Med. 2013, 369, 122–133. [Google Scholar] [CrossRef] [Green Version]
- Postow, M.; Chesney, J.; Pavlick, A.; Robert, C.; Grossmann, K.; McDermott, D.; Linette, G.; Meyer, N.; Giguere, J.; Agarwala, S.; et al. Nivolumab and ipilimumab versus ipilimumab in untreated melanoma. N. Engl. J. Med. 2015, 372, 2006–2017. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Lécuyer, E.; Blanchette, M. Prediction of mRNA subcellular localization using deep recurrent neural networks. Bioinformatics 2019, 35, i333–i342. [Google Scholar] [CrossRef] [Green Version]
- Castello, A.; Fischer, B.; Hentze, M.; Preiss, T. RNA-binding proteins in Mendelian disease. Trends Genet. TIG 2013, 29, 318–327. [Google Scholar] [CrossRef]
- Shu, L.; Yan, W.; Chen, X. RNPC1, an RNA-binding protein and a target of the p53 family, is required for maintaining the stability of the basal and stress-induced p21 transcript. Genes Dev. 2006, 20, 2961–2972. [Google Scholar] [CrossRef] [Green Version]
- Krackhardt, A.; Witzens, M.; Harig, S.; Hodi, F.; Zauls, A.; Chessia, M.; Barrett, P.; Gribben, J. Identification of tumor-associated antigens in chronic lymphocytic leukemia by SEREX. Blood 2002, 100, 2123–2131. [Google Scholar] [CrossRef] [PubMed]
- Tanner, M.; Grenman, S.; Koul, A.; Johannsson, O.; Meltzer, P.; Pejovic, T.; Borg, A.; Isola, J. Frequent amplification of chromosomal region 20q12-q13 in ovarian cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2000, 6, 1833–1839. [Google Scholar]
- Zheng, S.; Xu, J.; Isaacs, S.; Wiley, K.; Chang, B.; Bleecker, E.; Walsh, P.; Trent, J.; Meyers, D.; Isaacs, W. Evidence for a prostate cancer linkage to chromosome 20 in 159 hereditary prostate cancer families. Hum. Genet. 2001, 108, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shira, A.; Pinthus, J.; Rozovsky, U.; Goldstein, M.; Sellers, W.; Yaron, Y.; Eshhar, Z.; Orr-Urtreger, A. Multiple genes in human 20q13 chromosomal region are involved in an advanced prostate cancer xenograft. Cancer Res. 2002, 62, 6803–6807. [Google Scholar]
- Cho, S.; Jung, Y.; Zhang, J.; Chen, X. The RNA-binding protein RNPC1 stabilizes the mRNA encoding the RNA-binding protein HuR and cooperates with HuR to suppress cell proliferation. J. Biol. Chem. 2012, 287, 14535–14544. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zhang, Z.; Zhang, S.; Guo, Q.; Zhang, F.; Gao, L.; Ni, H.; Guo, X.; Xiang, C.; Xi, T. RNA Binding Protein RNPC1 Inhibits Breast Cancer Cell Metastasis via Activating STARD13-Correlated ceRNA Network. Mol. Pharm. 2018, 15, 2123–2132. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Y.; Ling, C.; Heng, W. RNPC1 inhibits non-small cell lung cancer progression via regulating miR-181a/CASC2 axis. Biotechnol. Lett. 2018, 40, 543–550. [Google Scholar] [CrossRef]
- Hötte, G.; Linam-Lennon, N.; Reynolds, J.; Maher, S. Radiation sensitivity of esophageal adenocarcinoma: The contribution of the RNA-binding protein RNPC1 and p21-mediated cell cycle arrest to radioresistance. Radiat. Res. 2012, 177, 272–279. [Google Scholar] [CrossRef]
- Ye, W.; Luo, C.; Liu, F.; Liu, Z.; Chen, F. CD96 Correlates With Immune Infiltration and Impacts Patient Prognosis: A Pan-Cancer Analysis. Front. Oncol. 2021, 11, 634617. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, Z.; Ma, L.; Li, C.; Lin, Y.; Yu, T.; Wei, J.; Zhu, L.; Yao, G. Effects of RNA methylation N6-methyladenosine regulators on malignant progression and prognosis of melanoma. Cancer Cell Int. 2021, 21, 453. [Google Scholar] [CrossRef]
- Simon, N.; Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Cox's Proportional Hazards Model via Coordinate Descent. J. Stat. Softw. 2011, 39, 1–13. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, W. Recruitment of bone marrow CD11bGr-1 cells by polymeric nanoparticles for antigen cross-presentation. Sci. Rep. 2017, 7, 44691. [Google Scholar] [CrossRef] [Green Version]
- Avula, L.; Knapen, D.; Buckinx, R.; Vergauwen, L.; Adriaensen, D.; Van Nassauw, L.; Timmermans, J. Whole-genome microarray analysis and functional characterization reveal distinct gene expression profiles and patterns in two mouse models of ileal inflammation. BMC Genom. 2012, 13, 377. [Google Scholar] [CrossRef] [Green Version]
- Bakhshi, T.; Georgel, P. Genetic and epigenetic determinants of diffuse large B-cell lymphoma. Blood Cancer J. 2020, 10, 123. [Google Scholar] [CrossRef]
- Fa, Z.; Xie, Q.; Fang, W.; Zhang, H.; Zhang, H.; Xu, J.; Pan, W.; Xu, J.; Olszewski, M.; Deng, X.; et al. RIPK3/Fas-Associated Death Domain Axis Regulates Pulmonary Immunopathology to Cryptococcal Infection Independent of Necroptosis. Front. Immunol. 2017, 8, 1055. [Google Scholar] [CrossRef] [Green Version]
- Brunner, G.; Reitz, M.; Heinecke, A.; Lippold, A.; Berking, C.; Suter, L.; Atzpodien, J. A nine-gene signature predicting clinical outcome in cutaneous melanoma. J. Cancer Res. Clin. Oncol. 2013, 139, 249–258. [Google Scholar] [CrossRef]
- Liu, N.; Liu, Z.; Liu, X.; Duan, X.; Huang, Y.; Jin, Z.; Niu, Y.; Zhang, L.; Chen, H. Identification of an Immune-Related Prognostic Signature Associated With Immune Infiltration in Melanoma. Front. Genet. 2020, 11, 1002. [Google Scholar] [CrossRef]
- Meyer, S.; Fuchs, T.J.; Bosserhoff, A.K.; Hofstadter, F.; Pauer, A.; Roth, V.; Buhmann, J.M.; Moll, I.; Anagnostou, N.; Brandner, J.M.; et al. A seven-marker signature and clinical outcome in malignant melanoma: A large-scale tissue-microarray study with two independent patient cohorts. PLoS ONE 2012, 7, e38222. [Google Scholar] [CrossRef] [Green Version]
- Givechian, K.; Wnuk, K.; Garner, C.; Benz, S.; Garban, H.; Rabizadeh, S.; Niazi, K.; Soon-Shiong, P. Identification of an immune gene expression signature associated with favorable clinical features in Treg-enriched patient tumor samples. NPJ Genom. Med. 2018, 3, 14. [Google Scholar] [CrossRef]
- Jung, T.; Haist, M.; Kuske, M.; Grabbe, S.; Bros, M. Immunomodulatory Properties of BRAF and MEK Inhibitors Used for Melanoma Therapy-Paradoxical ERK Activation and Beyond. Int. J. Mol. Sci. 2021, 22, 9890. [Google Scholar] [CrossRef]
- Bhatia, A.; Kumar, Y. Cancer-immune equilibrium: Questions unanswered. Cancer Microenviron. Off. J. Int. Cancer Microenviron. Soc. 2011, 4, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Shi, L.; Zhou, X.; Wu, J.; Xia, T.; Zhou, W.; Sun, X.; Zhu, L.; Wei, J.; Ding, Q. The role of c-Myc-RBM38 loop in the growth suppression in breast cancer. J. Exp. Clin. Cancer Res. CR 2017, 36, 49. [Google Scholar] [CrossRef]
- Ye, J.; Liang, R.; Bai, T.; Lin, Y.; Mai, R.; Wei, M.; Ye, X.; Li, L.; Wu, F. RBM38 plays a tumor-suppressor role via stabilizing the p53-mdm2 loop function in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. CR 2018, 37, 212. [Google Scholar] [CrossRef]
- Xue, J.; Xia, T.; Liang, X.; Zhou, W.; Cheng, L.; Shi, L.; Wang, Y.; Ding, Q. RNA-binding protein RNPC1: Acting as a tumor suppressor in breast cancer. BMC Cancer 2014, 14, 322. [Google Scholar] [CrossRef] [Green Version]
- Wampfler, J.; Federzoni, E.; Torbett, B.; Fey, M.; Tschan, M. The RNA binding proteins RBM38 and DND1 are repressed in AML and have a novel function in APL differentiation. Leuk. Res. 2016, 41, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Wei, X.; Ni, W.; Cao, M.; Meng, L.; Yang, H. The expression of RNA-binding protein RBM38 decreased in renal cell carcinoma and represses renal cancer cell proliferation, migration, and invasion. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2017, 39, 1010428317701635. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Wu, J.; Shi, L.; Li, X.; Zhu, L.; Sun, X.; Qian, J.; Wang, Y.; Wei, J.; Ding, Q. PTEN expression is upregulated by a RNA-binding protein RBM38 via enhancing its mRNA stability in breast cancer. J. Exp. Clin. Cancer Res. CR 2017, 36, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Gu, J.; Li, X.; Wang, Q.; Ding, Y. RNA-binding protein RBM38 acts as a tumor suppressor in gastric cancer. Int. J. Clin. Exp. Pathol. 2017, 10, 11130–11136. [Google Scholar]
- Zhang, J.; Cho, S.; Shu, L.; Yan, W.; Guerrero, T.; Kent, M.; Skorupski, K.; Chen, H.; Chen, X. Translational repression of p53 by RNPC1, a p53 target overexpressed in lymphomas. Genes Dev. 2011, 25, 1528–1543. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Ma, C.; Shao, J.; Cao, J. Prognostic Implications of Novel Ten-Gene Signature in Uveal Melanoma. Front. Oncol. 2020, 10, 567512. [Google Scholar] [CrossRef]
- Guo, W.; Zhu, L.; Zhu, R.; Chen, Q.; Wang, Q.; Chen, J. A four-DNA methylation biomarker is a superior predictor of survival of patients with cutaneous melanoma. eLife 2019, 8, e44310. [Google Scholar] [CrossRef] [PubMed]
- Roszik, J.; Mustachio, L.; Livingston, J.; Groisberg, R.; Carmagnani Pestana, R.; Subbiah, V.; Conley, A. Landscape of Immune-Related Markers and Potential Therapeutic Targets in Soft Tissue Sarcoma. Cancers 2021, 13, 5249. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Ma, C.; Yang, L.; Sun, Y.; Zhang, Y. Prognostic Value and Immunological Characteristics of a Novel RNA Binding Protein Signature in Cutaneous Melanoma. Front. Genet. 2021, 12, 723796. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Chen, Y. Tumor microenvironment: Sanctuary of the devil. Cancer Lett. 2015, 368, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.; Zitvogel, L.; Sautès-Fridman, C.; Kroemer, G. The immune contexture in cancer prognosis and treatment. Nat. Rev. Clin. Oncol. 2017, 14, 717–734. [Google Scholar] [CrossRef]
- Cipponi, A.; Wieers, G.; van Baren, N.; Coulie, P. Tumor-infiltrating lymphocytes: Apparently good for melanoma patients. But why? Cancer Immunol. Immunother. CII 2011, 60, 1153–1160. [Google Scholar] [CrossRef]
- McGranahan, N.; Furness, A.; Rosenthal, R.; Ramskov, S.; Lyngaa, R.; Saini, S.; Jamal-Hanjani, M.; Wilson, G.; Birkbak, N.; Hiley, C.; et al. Clonal neoantigens elicit T cell immunoreactivity and sensitivity to immune checkpoint blockade. Science 2016, 351, 1463–1469. [Google Scholar] [CrossRef] [Green Version]
- Clemente, C.; Mihm, M.; Bufalino, R.; Zurrida, S.; Collini, P.; Cascinelli, N. Prognostic value of tumor infiltrating lymphocytes in the vertical growth phase of primary cutaneous melanoma. Cancer 1996, 77, 1303–1310. [Google Scholar] [CrossRef]
- van Houdt, I.; Sluijter, B.; Moesbergen, L.; Vos, W.; de Gruijl, T.; Molenkamp, B.; van den Eertwegh, A.; Hooijberg, E.; van Leeuwen, P.; Meijer, C.; et al. Favorable outcome in clinically stage II melanoma patients is associated with the presence of activated tumor infiltrating T-lymphocytes and preserved MHC class I antigen expression. Int. J. Cancer 2008, 123, 609–615. [Google Scholar] [CrossRef]
- Nsengimana, J.; Laye, J.; Filia, A.; O'Shea, S.; Muralidhar, S.; Poźniak, J.; Droop, A.; Chan, M.; Walker, C.; Parkinson, L.; et al. β-Catenin-mediated immune evasion pathway frequently operates in primary cutaneous melanomas. J. Clin. Investig. 2018, 128, 2048–2063. [Google Scholar] [CrossRef] [Green Version]
- McArthur, G.; Chapman, P.; Robert, C.; Larkin, J.; Haanen, J.; Dummer, R.; Ribas, A.; Hogg, D.; Hamid, O.; Ascierto, P.; et al. Safety and efficacy of vemurafenib in BRAF(V600E) and BRAF(V600K) mutation-positive melanoma (BRIM-3): Extended follow-up of a phase 3, randomised, open-label study. Lancet Oncol. 2014, 15, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Hauschild, A.; Grob, J.; Demidov, L.; Jouary, T.; Gutzmer, R.; Millward, M.; Rutkowski, P.; Blank, C.; Miller, W.; Kaempgen, E.; et al. Dabrafenib in BRAF-mutated metastatic melanoma: A multicentre, open-label, phase 3 randomised controlled trial. Lancet 2012, 380, 358–365. [Google Scholar] [CrossRef]
- Robert, C.; Thomas, L.; Bondarenko, I.; O’Day, S.; Weber, J.; Garbe, C.; Lebbe, C.; Baurain, J.; Testori, A.; Grob, J.; et al. Ipilimumab plus dacarbazine for previously untreated metastatic melanoma. N. Engl. J. Med. 2011, 364, 2517–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knox, T.; Sahakian, E.; Banik, D.; Hadley, M.; Palmer, E.; Noonepalle, S.; Kim, J.; Powers, J.; Gracia-Hernandez, M.; Oliveira, V.; et al. Selective HDAC6 inhibitors improve anti-PD-1 immune checkpoint blockade therapy by decreasing the anti-inflammatory phenotype of macrophages and down-regulation of immunosuppressive proteins in tumor cells. Sci. Rep. 2019, 9, 6136. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Id | Coef | HR | HR.95L | HR.95H | p-Value |
|---|---|---|---|---|---|
| A2M | −0.097 | 0.908 | 0.824 | 1.000 | 0.050 |
| NAMPT | −0.185 | 0.831 | 0.688 | 1.004 | 0.055 |
| LIF | −0.123 | 0.885 | 0.809 | 0.967 | 0.007 |
| EBI3 | −0.247 | 0.781 | 0.674 | 0.905 | 0.001 |
| ERAP1 | −0.170 | 0.844 | 0.668 | 1.066 | 0.155 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Xu, J.; Luo, B.; Tang, J.; Hou, Z.; Zhu, Z.; Zhu, L.; Yao, G.; Li, C. Immune Landscape and an RBM38-Associated Immune Prognostic Model with Laboratory Verification in Malignant Melanoma. Cancers 2022, 14, 1590. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14061590
Liu J, Xu J, Luo B, Tang J, Hou Z, Zhu Z, Zhu L, Yao G, Li C. Immune Landscape and an RBM38-Associated Immune Prognostic Model with Laboratory Verification in Malignant Melanoma. Cancers. 2022; 14(6):1590. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14061590
Chicago/Turabian StyleLiu, Jinfang, Jun Xu, Binlin Luo, Jian Tang, Zuoqiong Hou, Zhechen Zhu, Lingjun Zhu, Gang Yao, and Chujun Li. 2022. "Immune Landscape and an RBM38-Associated Immune Prognostic Model with Laboratory Verification in Malignant Melanoma" Cancers 14, no. 6: 1590. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14061590