Cascade Biocatalysis Designed for the Allylic Oxidation of α-Pinene



Department of Organic Chemistry, Biochemistry and Catalysis, Faculty of Chemistry, University of Bucharest, 030018 Bucharest, Romania
*
Author to whom correspondence should be addressed.
Catalysts 2021, 11(1), 134; https://0-doi-org.brum.beds.ac.uk/10.3390/catal11010134
Submission received: 29 December 2020
/
Revised: 13 January 2021
/
Accepted: 15 January 2021
/
Published: 18 January 2021
(This article belongs to the Special Issue Biocatalysis for Green Chemistry)
Abstract
:A biocatalytic cascade system using a cocktail of oxidoreductase enzymes (2-1B peroxidase and M120 laccase) was designed for the allylic oxidation of (+)-α-pinene into value-added products (e.g., verbenol and verbenone). The oxidative transformation involved a two-step process as follows: (+)-α-pinene was (i) oxidized on the allylic position with H2O2 mainly assisted by 2-1B peroxidase leading to verbenol as the principal reaction product, and (ii) directed to verbenone in the presence of M120 laccase responsible for further oxidation of verbenol to verbenone. The reaction environment was ensured by the acetate buffer (0.1 M, pH = 5). Optimum values for the experimental parameters (e.g., concentration of 2-1B peroxidase, M120 laccase, and H2O2) were set up. The biocatalytic cascade process was monitored for 24 h in order to evaluate the process pathway. Maximum performance under optimum conditions was reached after 5 h incubation time (e.g., 80% (+)-α-pinene conversion and 70% yield in verbenol). Therefore, the developed biocatalytic cascade system offered promising perspectives for (+)-α-pinene valorization.
1. Introduction
Currently, terpenes, especially monoterpenes, represent one of the most chemically approached class of natural compounds with extensive therapeutic properties and, generally, many examples of involvement in industrial applications. Therefore, monoterpenes have been brought into the spotlight due to their role in the prevention or therapy of many diseases [1,2]; in insecticidal and antimicrobial activity, as being useful in preserving crops and soil cultures [3]; or in the mass production of valuable natural precursors, flavors, and fragrances [4]. α-Pinene is one of these monoterpenoid compounds that can be found in turpentine resin together with β-pinene, limonene, 3-carene, myrcene, camphene, and p-cymene [5]. More than 160,000 tons per year of α-pinene accumulate as a basic component of turpentine from wood and citrus oil processing [6]. Additionally, the pulp and paper industry offers generous turpentine waste with a high content of α-pinene [7]. Thus, the valorization of the industrial residues of α-pinene, which have demonstrated good volatility and organoleptic properties, is a real requirement of today’s industry.
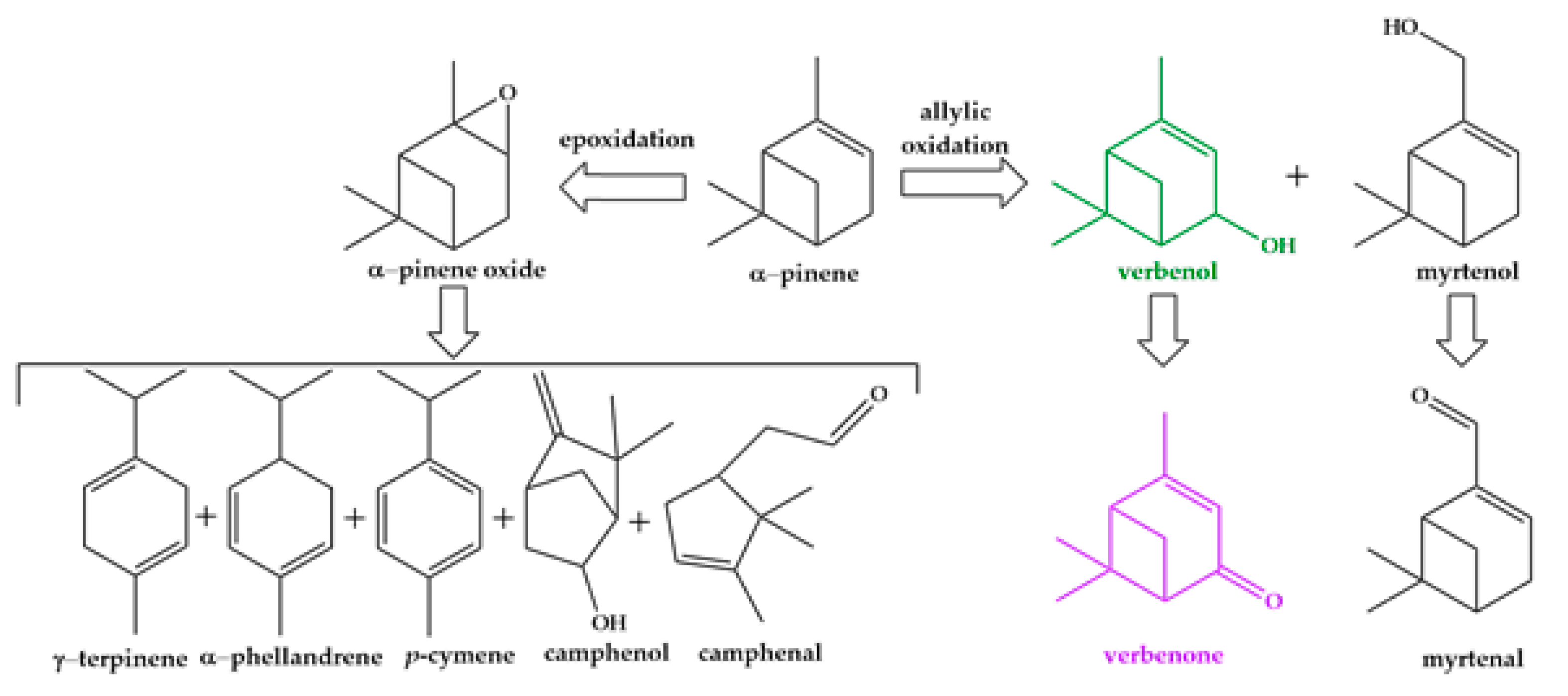
Oxidative functionalization of α-pinene leading to chemical products capitalized on the natural derivatives useful as flavors and fragrances is considered an alternative with high potential in industrial applications [8]. Oxidative transformations of α-pinene can take the following two competitive routes: (1) epoxidation of the double bond of the cycle leading to pinene epoxide and further to varied monoterpenoid derivatives and (2) allylic oxidation of α-pinene by the production of verbenol–verbenone and myrtenol–myrtenal mixtures [9] (Scheme 1). Both pathways could be achieved based on conventional catalysis [8,10,11]. However, the drawbacks of a catalytic process, such as technological and environmental problems resulting from the (re)generation of the catalyst and the use of a large number of effluents [12], launch the challenge of developing new alternatives. In this context, biocatalysis is a valuable solution offering a green alternative for α-pinene oxidation and leading to (oxi)derivatives valuable for the flavors and fragrances industry.
Taking into account the allylic oxidation based on biocatalytic transformation, the reported systems are mainly based on a whole-cell biocatalyst [5,7,13,14,15,16]. An intergeneric hybrid produced by the protoplastic fusion of Aspergillus niger (A. niger) and Penicillium digitatum (P. digitatum) biotransformed (−)-α-pinene into (−)-verbenol, with an overall yield of 60%, compared to the strains of the parental cells (10% and 15% yield in (−)-verbenol for A. niger and P. digitatum, respectively) [13]. In plant cell cultures, Penicillium brachyceras showed substrate specificity for (−)-α-pinene, oxidizing it to verbenone with a conversion of 81%, while Rauwolfia sellowii was capable of 38% conversion of both α-pinene enantiomers [14]. Bacteria formed by the degradation of citrus fruits with a strain coming from Gluconobacter japonicus proved the ability to convert 96% of (+)-α-pinene into verbenone during eight days of incubation [7]. Additionally, both enantiomers of α-pinene and their racemic mixture allowed to sequentially produce verbenol by treatment with Picea abies cell suspension (35% substrate conversion). The prolongation of the reaction time led to verbenone as the main product (68% substrate conversion) [15]. Interestingly, Picea abies-immobilized cells tend to preferably produce pinocarveol and α-terpineol over verbenol [5]. Allylic hydroxylation of triterpenes was achieved by incubating the substrate with bacterial culture of Bacillus megaterium (considering P450s monooxygenase activity). Erythrodiol, uvaol, and hederagenin as substrates allowed to produce the corresponding hydroxy derivatives with yields ranging between 10% and 30% [16]. Therefore, there is a great diversity of microorganisms that contribute to the biotransformation of terpenes (e.g., α-pinene) into natural flavoring compounds. However, several shortcomings (e.g., the cytotoxicity of precursors and products, the difference in hydrophobicity between organic compounds and the growth medium, etc., [17]) have been also noticed as limiting the applicability of microorganism-based biocatalysis.
Enzyme biocatalysis could be a solution for avoiding the drawbacks of microorganism-based biocatalysis presented previously. Cytochrome hem-monooxygenase P450cam from Pseudomonas putida has been reported in the literature for the specific oxidation of α-pinene. The major product was verbenol (86% selectivity in verbenol with only 5% selectivity in verbenone) [18]. Our research group reported previously the biocatalytic activity of the modified unspecific peroxidase (PaDa-I as unspecific peroxidase variant expressed in Saccharomyces cerevisiae, characterized by the intermediate catalytic behavior between P450 monooxygenase and hem-peroxidase) [19] for the allylic oxidation of α-pinene [20]. The biocatalytic oxidation was performed in a biphasic system in order to avoid the inconvenience of low solubility of the substrate or products in aqueous phase. System performance reached maximum 68% α-pinene conversion and 44% yield in verbenol [20].
Recently, a two-step biocatalytic process of allylic oxidation based on the cascading action of modified P450 monooxygenase and alcohol dehydrogenase enzyme has been reported [21]. Isophorone substrate as one valuable monooxygenated terpenoid was transformed into ketoisophorone, useful as a building block in vitamin synthesis. The P450 mutant successfully contributes to the formation of 4-hydroxy-α-isophorone (99% catalytic performance expressed in total turnover numbers), further oxidized by alcohol dehydrogenase to the corresponding keto derivative (maximum 92% conversion of substrate) [21].
In this study, we report the development of a linear bi-enzymatic cascade process based on the synergistic communication between versatile peroxidase (2-1B peroxidase) and laccase (M120 laccase) enzymes for the allylic oxidation of (+)-α-pinene leading predominantly to verbenol. The oxidative process involves the following two steps: (i) the allylic oxidation of (+)-α-pinene mainly assisted by 2-1B peroxidase to verbenol as the principal reaction product and (ii) the oxidation of verbenol directed to verbenone in the presence of M120 laccase. H2O2 was added as oxidation reagent for the peroxidase enzyme, while dissolved O2 in the reaction phase was required by the laccase enzyme. The 2-1B peroxidase mutant is an extracellular versatile peroxidase found in Pleurotus eryngii, expressed in Saccharomices cerevisiae (S. cerevisiae) as a stable and highly reactive enzyme, requiring stoichiometric concentrations of H2O2. 2-1B peroxidase was prepared by the modification of the hydrophobic core of the putative peptide, contributing to the improvement of the catalytic efficiency for peroxidative reactions and oxygen transfer [22]. The ability to retain its catalytic activity even at higher concentrations of H2O2 was improved due to the implied structural modifications [19]. Furthermore, the introduced mutations increased enzyme thermostability and oxidation activity even in alkaline conditions. Maximum activity for the 2-1B peroxidase variant was reported at pH 5 and was taken as reference value. Additionally, a residual activity was noticed up to pH 9 [23].
In contrast, laccases represent a particular group of blue copper oxidative enzymes. They are glycoproteins, ubiquitous in nature, found in plants and in almost every type of fungus. Laccase-catalyzed reactions are carried out through the mono-electronic oxidation of a substrate molecule (aromatic or aliphatic phenols/amines) to a corresponding reactive radical [24]. The redox process is performed by assisting the cluster of four copper atoms, which forms the catalytic core of the enzyme. However, frequently, the substrate of interest cannot be oxidized directly by laccases, either because the substrate is too large to be integrated into the active site of this enzyme, or because the enzyme has high redox potential. These limitations might be corrected by mediators, while the oxidized radical state will be able to interact with the enzyme with high redox potential. Specifically, laccases may be used in connection with peroxidases, to produce mild oxidation reactions with the formation of alcohol or aldehyde/ketone products [25].
In the present study, the use of a cascade process catalyzed by the enzymatic mixture (1-2B peroxidase and M120 laccase) was strongly related to the idea of improving the catalytic activity of the peroxidase enzyme in oxidative conditions with the support of the laccase enzyme. Experimental approaches on setting up the cascade system were performed for the oxidative biotransformation of (+)-α-pinene directed preponderantly to the verbenol product and further oxidized to verbenone. Besides these, myrtenol and myrtenal were also considered as secondary products of the process. Optimization of the biocatalytic process was performed considering the enzymatic (peroxidase and laccase) biocatalyst and oxidant (H2O2) concentrations. Additionally, monitoring in time of the (+)-α-pinene biotransformation was achieved.
2. Results and Discussion
Cascade biocatalysis was set up and evaluated for (+)-α-pinene oxidation. The biocatalytic system contained an enzyme cocktail of 2-1B peroxidase and M120 laccase used as bi-enzyme biocatalyst and H2O2 as oxidation reagent specific for the peroxidase enzyme. Additionally, 0.1 M acetate buffer (pH = 5) guaranteed the acidic environment suitable for optimum catalytic activity of 2-1B peroxidase. Kinetic measurements were performed for both enzymes separately and in mixture. Kinetic/catalytic constants (Km, kcat and kcat/Km) were calculated in order to evaluate the activity of both enzymes (Table 1). 2-1B peroxidase exhibited higher catalytic efficiency (kcat/Km) than M120 laccase (5.30 ± 0.92 and 0.91 ± 0.09 s−1 mM−1 of 2-1B peroxidase and M120 laccase, respectively). Noticeably, the catalytic efficiency of 2-1B peroxidase was improved when M120 laccase was added to the reaction mixture.
2.1. Influence of 2-1B Peroxidase on the Biocatalytic Cascade System
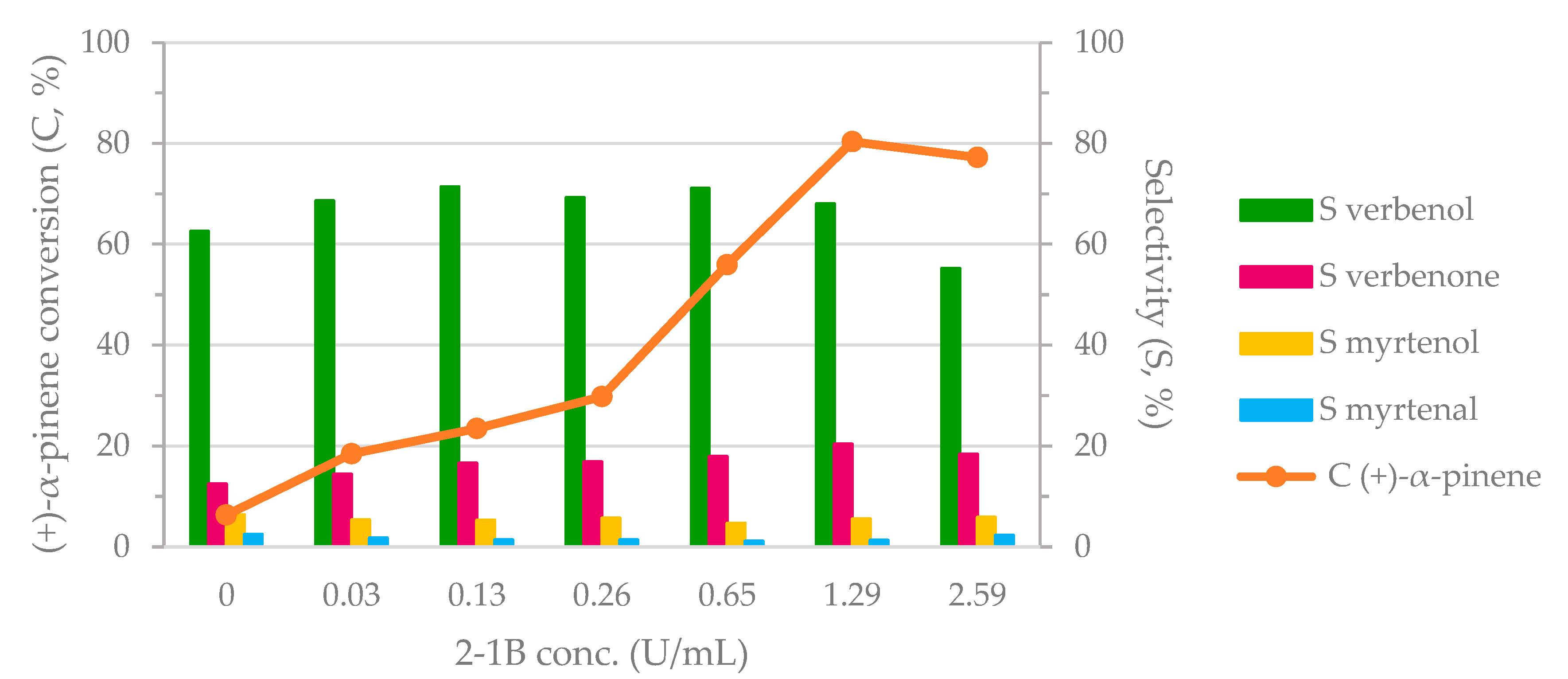
Different concentrations of 2-1B peroxidase were tested in the proposed biocatalytic system. The experimental results are presented in Figure 1. An upward trend was observed for substrate conversion when the concentration of 2-1B peroxidase increased from 0 to 1.29 U/mL (12.97 U/mL of stock solution of 2-1B peroxidase). A higher concentration than 1.29 U/mL (2.59 U/mL) did not substantially affect the substrate conversion. The content in verbenol remained constant (~70% selectivity in verbenol) up to 0.65 U/mL of 2-1B peroxidase. Then, the formation of verbenol decreased up to 2.59 U/mL 2-1B peroxidase concentration. Over the same time, the production of verbenone and the other oxidative derivatives, such as myrtenol and myrtenal, was favored for high concentrations of the peroxidase enzyme (>0.65 U/mL). However, secondary oxidation products (e.g., myrtenol and myrtenal) cumulated no more than 10–15% abundance in the products’ mixture. All the reaction products were identified based on GC–MS (gas chromatography coupled with mass spectrometry detection) analysis and quantified using HPLC-DAD (high performance liquid chromatography coupled with diode array detection) analysis. It can be seen that the allylic oxidation of (+)-α-pinene depended directly on the 2-1B peroxidase concentration (i.e., the total number of peroxidase catalytic sites). In this context, 1.29 U/mL was considered the optimal concentration of 2-1B peroxidase as a compromise between (+)-α-pinene conversion and verbenol production (80% (+)-α-pinene conversion and 68% selectivity in verbenol, as the favored product). This peroxidase concentration was used for the next experiments. It must be noted that, for 0 U/mL 2-1B peroxidase, only 6% of substrate was transformed. Therefore, M120 laccase (216 POU/mL) exhibited low catalytic capacity for the oxidation of (+)-α-pinene in the absence of the peroxidase enzyme.
2.2. Influence of M120 Laccase on the Biocatalytic Cascade System
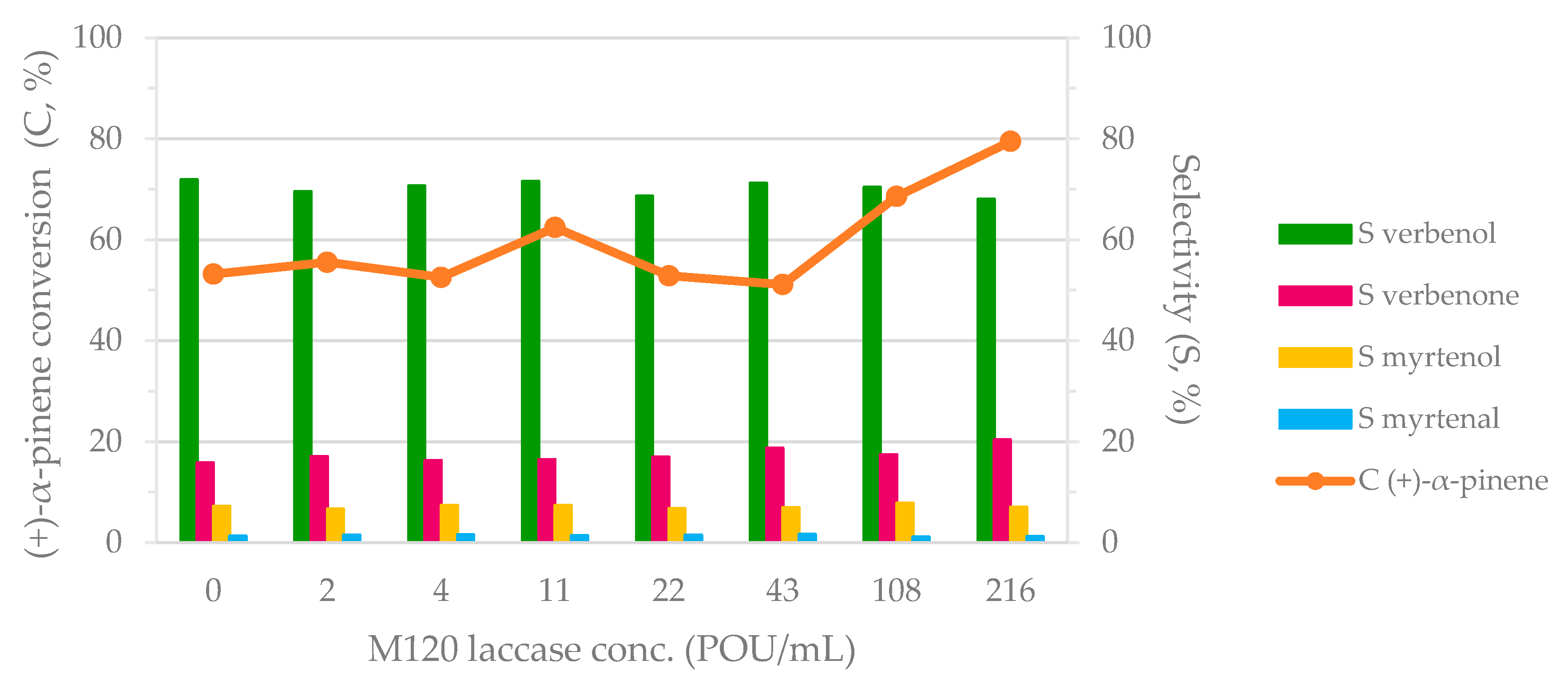
The biocatalytic cascade process was performed varying the concentration of laccase in the range of 0–216 POU/mL. The results are shown in Figure 2. Low concentrations of M120 laccase (<43 POU/mL M120 laccase) did not exhibit any effects on the (+)-α-pinene transformation. High laccase concentrations (43–216 POU/mL) led to the strong improvement of (+)-α-pinene oxidation. Therefore, the conversion of (+)-α-pinene was improved from 51% to 80% (+)-α-pinene when the M120 laccase concentration varied in the range of 43–216 POU/mL. No visible modification was observed in terms of verbenol production (~70% selectivity in verbenol for all the cases). However, a slight increase in selectivity in myrtenol and myrtenal was observed for the variation of M120 laccase concentration. In the absence of M120 laccase, 52% (+)-α-pinene was converted with the preferential formation of verbenol, indicating the capacity of the 2-1B peroxidase to catalyze the (+)-α-pinene oxidation (Figure 2).
The use of M120 laccase for (+)-α-pinene oxidative biotransformation allowed a significant improvement in the substrate conversion (maximum to 80% (+)-α-pinene conversion), preserving the preferred product as verbenol (around 70% selectivity in verbenol for 216 POU/mL M120 laccase activity). This means that M120 laccase supported the peroxidase activity. Additionally, the evaluation of the catalytic efficiency of 2-1B peroxidase in the presence and absence of M120 laccase confirms the laccase contribution (Table 1). Based on the experimental results, we presumed that the allylic oxidation of (+)-α-pinene to verbenol was catalyzed predominantly by 2-1B peroxidase, whereas M120 laccase favoring the oxidation of verbenol to verbenone allowed the equilibrium of the allylic oxidation to move to a greater substrate consumption. Similarly, the myrtenol conversion into myrtenal could also have an influence on this process, but with a small contribution since the myrtenol–myrtenal transformation was achieved with low efficiency.
2.3. Influence of H2O2 on the Biocatalytic Cascade System
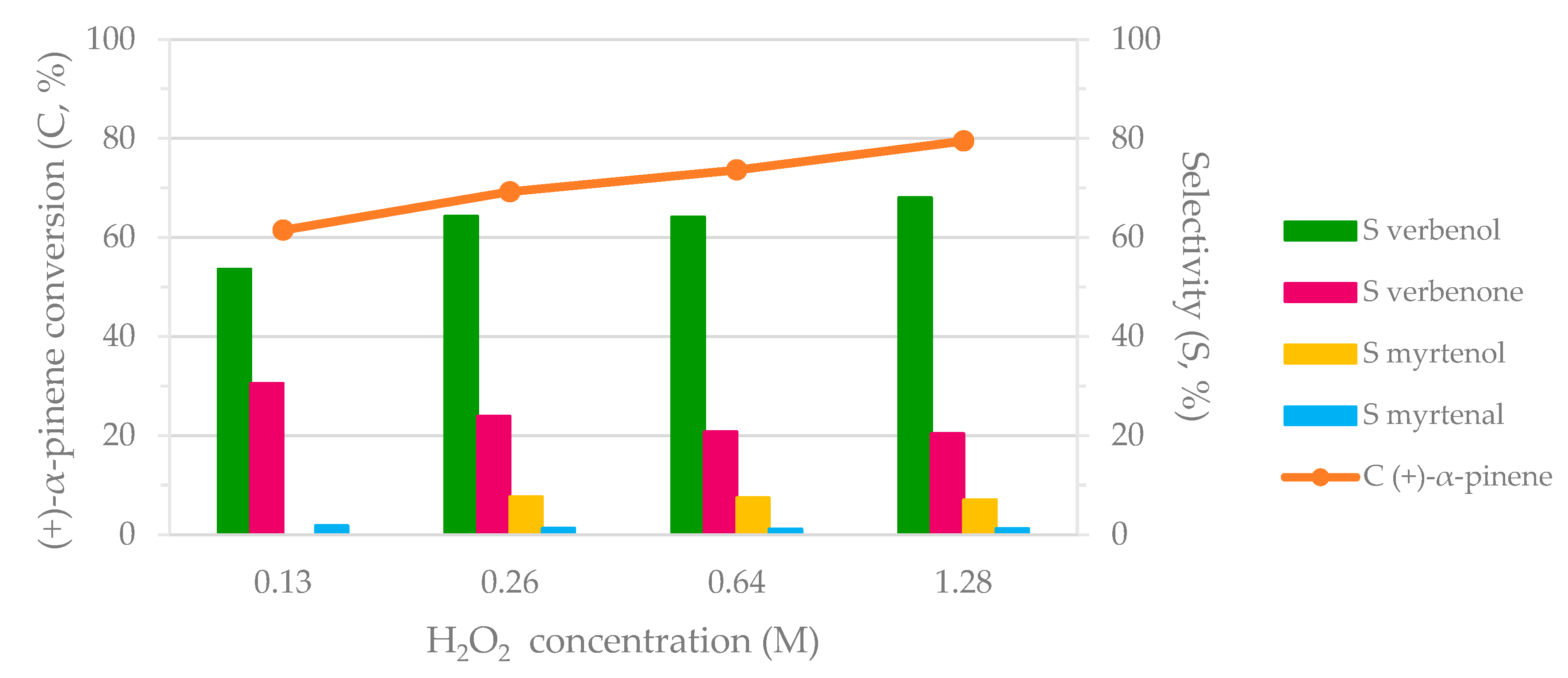
H2O2 concentration was also varied in the biocatalytic system. The experimental results in terms of (+)-α-pinene conversion and verbenol/verbenone formation were considered. The corresponding results are presented in Figure 3. When H2O2 concentration was raised from 0.13 to 1.28 M, the (+)-α-pinene conversion exhibited an ascending trend varying from 62% to 80%, respectively. Additionally, an increase in the selectivity for verbenol was achieved, whereas the selectivity in verbenone decreased. The ratio between verbenol and verbenone indicated that the peroxidase step gained in the presence of laccase when H2O2 concentration was raised to 1.28 M. No visible variation of selectivity in myrtenol/myrtenal (<10% selectivity in myrtenol/myrtenal) was noted for all the cases.
2-1B peroxidase used H2O2 as the source of OH• radicals in order to insert the OH group onto the substrate structure in the proximity of the carbon double bound (allylic oxidation). It is already known that high concentrations of H2O2 can usually inhibit the catalytic activity of the common peroxidase enzyme. However, 2-1B peroxidase is a mutant of versatile peroxidase that involves structural modifications in order to suppress the enzyme inhibition at high concentrations of H2O2 [19]. Consequently, 2-1B peroxidase preserved its catalytic activity for H2O2 concentrations up to 1.28 M. Stability tests confirmed the resistance of 2-1B peroxidase at high H2O2 concentrations (Figure S1, Supplementary Materials). It should be mentioned that H2O2 is also a competitive inhibitor of M120 laccase. However, M120 laccase exhibited appreciable catalytic efficiency at high H2O2 concentrations. Consequently, the enzymatic cooperativity of both enzymes was observed, leading to the promising degree of (+)-α-pinene biotransformation (80% (+)-α-pinene conversion, and 68% and 20% selectivity in verbenol and verbenone, respectively).
2.4. Influence of Reaction Time on the Biocatalytic Cascade System
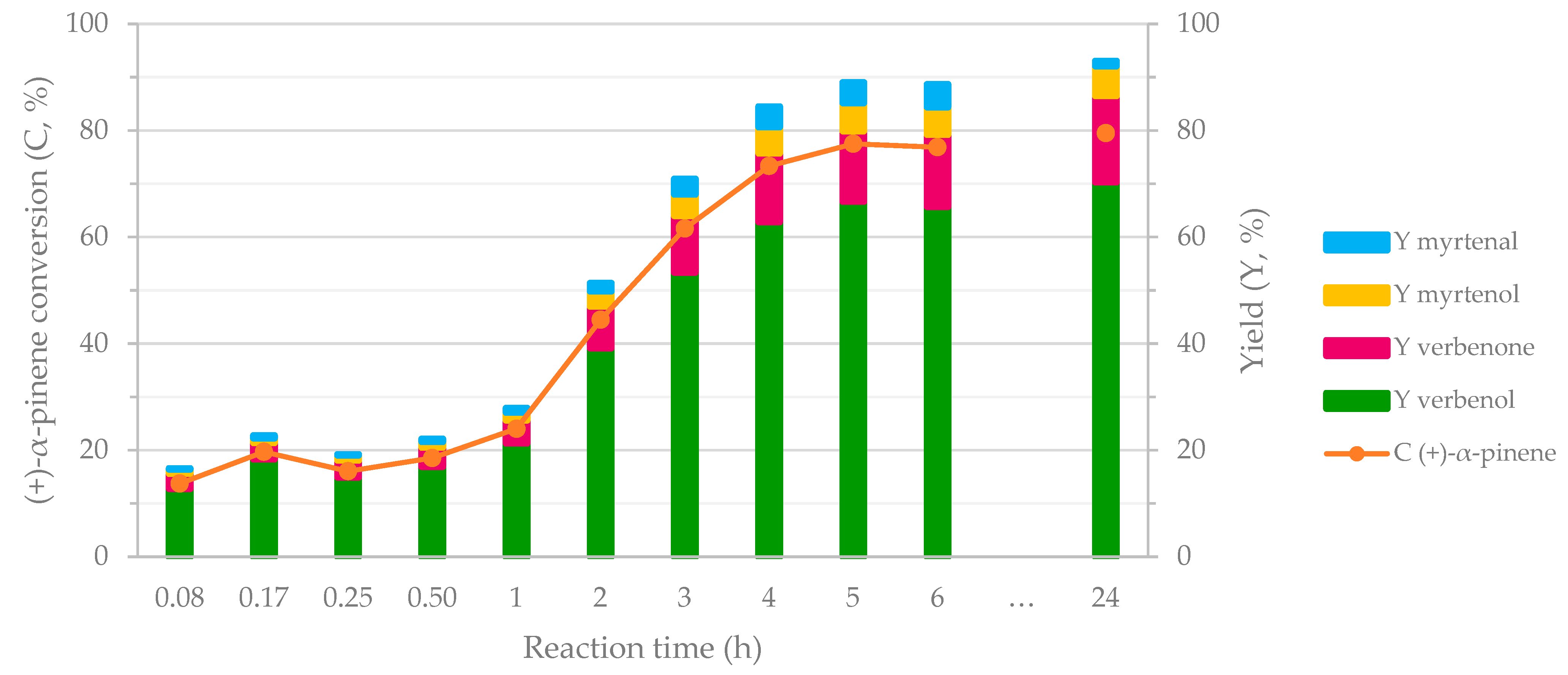
To evaluate the global performance of the biocatalytic cascade process, 10 reaction mixtures with the same composition were prepared and incubated under similar experimental conditions. The samples were analyzed at different moments over 24 h. Figure 4 presents the corresponding results. For the first hour of reaction, no visible modifications in the (+)-α-pinene conversion and product yield were noticed. Subsequently, a rapid increase in (+)-α-pinene conversion was registered for the next 4 h. Therefore, the maximum transformation of (+)-α-pinene (around 80% (+)-α-pinene conversion) was achieved in 5 h incubation time. An increase in verbenol production was also noticed up to 67% yield. Simultaneously, the same ascending trend was observed for verbenone, myrtenol, and myrtenal. Subsequently, the substrate conversion remained constant followed by a slight increase in verbenol (up to 70% yield in verbenol) during the next 19 h. It should be noted that verbenol was the favored product from the first hour of the reaction time. These are clear indications that the allylic oxidation of (+)-α-pinene dominated the cascade process from the beginning. The 2-1B peroxidase enzyme was mainly responsible for this behavior. Additionally, the cascade established the reactions’ equilibrium after 5 h. Therefore, 5 h were enough to efficiently yield verbenol from (+)-α-pinene (around 70% yield of verbenol). Noticeably, both enzymes preserved their catalytic activity under the system conditions (88% and 95% recovery of the catalytic activity for 2-1B peroxidase and M120 laccase after 5 h incubation in the cascade system) (Figure S1, Supplementary Materials).
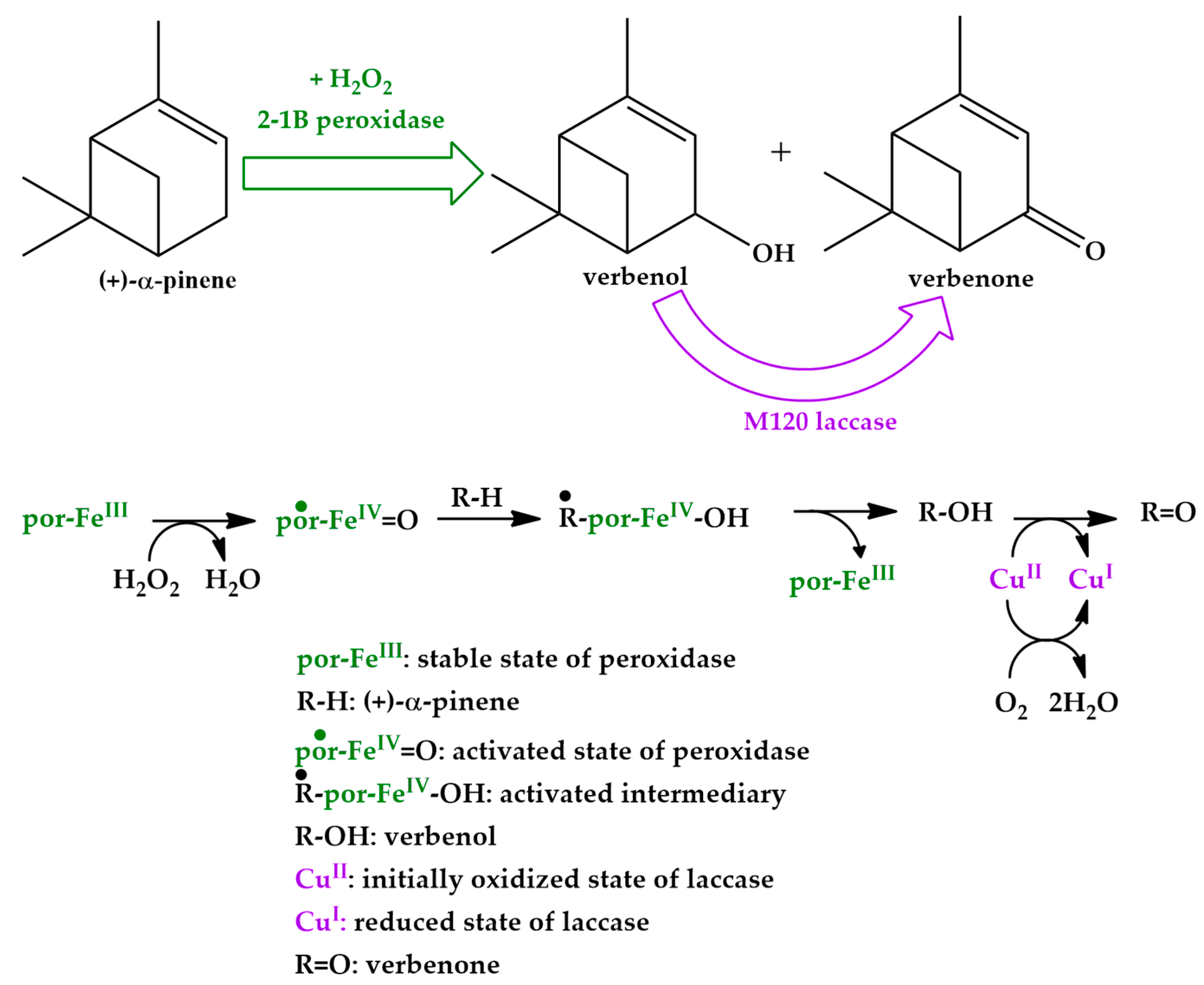
Based on the experimental results presented in Figure 4, the mechanism of the developed process was proposed (Scheme 2). The setup system is a typical biocatalytic cascade design (as “domino” reactions [26]) with a bi-enzymatic biocatalyst (2-1B peroxidase and M120 laccase). The mechanism involves the linking of individual stages of the entire chemical transformation process of (+)-α-pinene oxidation. (+)-α-Pinene, H2O2, and the enzymes were added at the beginning, leading to the formation of the verbenol product, and continuing the biocatalytic process further to verbenone formation. Moreover, myrtenol and myrtenal were produced, but with low selectivity (<10%).
The hypothetical mechanism involving the electron exchange steps is also detailed in Scheme 2. The catalytic site of 2-1B versatile peroxidase, which is essentially provided with a trivalent porphyrinic FeIII, transforms into the radical FeIV complex after H2O2 contribution and captures the substrate susceptible to oxidation, (+)-α-pinene [27]. M120 laccase focuses on the freshly formed verbenol with a proton and electron transfer mechanism supported by acid pH 5 and the rearrangement onto the energy levels drives to the formation of verbenone [28]. Therefore, the presence of M120 laccase allowed an apparent enhancement of 2-1B peroxidase activity by favoring the substrate consumption. The conclusion is supported by both experimental sets for 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) and (+)-α-pinene substrates.
3. Materials and Methods
3.1. Chemicals and Solutions
The reagents used in the experiments were all purchased from Sigma-Aldrich, Taufkirchen, Germany: (+)-α-pinene (ρ = 0.86g/cm3), H2O2 (30%), acetonitrile (99.8% purity for HPLC), sodium acetate, and glacial acetic acid for the preparation of the acetate buffer solution. The enzyme mixture contains 2-1B peroxidase and M120 laccase. 2-1B peroxidase was kindly offered by Dr. Miguel Alcalde from the Biocatalysis Department affiliated with the Catalysis Institute of CSIC, Madrid, Spain. 2-1B peroxidase was prepared as a mutant of versatile peroxidase from Pleurotus eryngii expressed in Saccharomyces cerevisiae cells. It was obtained synthetically by modifying the hydrophobic part around the catalytic zone of the putative peroxidase enzyme. The enzymatic activity was estimated at 12.97 U/mL. M120 laccase (thermostable, extracted from the Trametes sp. fungal family) was offered by Dr. Fumiaki Ito from Amano Enzyme Europe Limited (Oxfordshire, UK). The enzyme activity indicated by the provider was 108,000 POU/g. Acetate buffer was prepared by dissolving 1.36 g of sodium acetate in 100 mL deionized water. The pH of the solution was adjusted to 5 by adding glacial acetic acid.
3.2. Enzyme Activity
Kinetic constants of both enzymes (2-1B peroxidase and M120 laccase) were determined spectrophotometrically using ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt) as substrate according to the standard method [29]. The activity assays were carried out in 0.1 M acetate buffer (pH = 5) for substrate concentrations in the range of 10–600 μM and 8 mM H2O2 (for 2-1B peroxidase). The absorbance was recorded at 420 nm (ε = 36,000 M−1 cm−1). Kinetic constants (Km) for both 2-1B peroxidase and M120 laccase (Table 1) were calculated assuming that a one-side saturation ligand was binding to the active site of the enzyme. For the mixture of both enzymes, an apparent kinetic constant of 2-1B peroxidase was calculated using the ABTS-based protocol as in the previous cases and mimicking the conditions of the biocatalytic cascade system in terms of the 2-1B peroxidase/M120 laccase ratio. Furthermore, kcat and kcat/Km were calculated using the results of the ABTS analysis (Table 1). All experiments were performed in triplicate.
3.3. Preparation of the Reaction Mixture
The reaction sample was prepared by mixing the reactants and biocatalysts at certain amounts in the solution of 0.1 M acetate buffer (pH = 5), leading to a composition of 0.63 M (+)-α-pinene, 0–2.59 U/mL 2-1B peroxidase, 0.13–1.28 M H2O2, and 0–216 POU/mL M120 laccase. The total volume of the reaction mixture was 500 µL. All experiments were performed in duplicate (relative standard deviation (RDS) < 20%).
The reaction mixture was incubated at 40 °C for a maximum of 24 h under stirring (1000 rpm) conditions. After the reaction, the sample was cooled to room temperature. Before high-performance liquid chromatographic (HPLC) analysis, the sample was treated with 250 µL acetonitrile to precipitate the enzymes and achieve the proper phase compatible with the mobile phase of the HPLC system. Then, the obtained mixture was vortexed for 10 min. The enzymes precipitated in the presence of the organic solvent. The centrifugation step allowed to remove the precipitate and to collect the supernatant for HPLC analysis. Moreover, the supernatant was passed through 0.2 µm porosity microfilters just before analysis in order to ensure a homogeneous sample phase.
Stability tests of both enzymes separately and in mixture were performed in order to evaluate the recovery of the enzyme activity. The enzyme(s) and H2O2 were added to the acetate buffer (0.1 M, pH = 5) at the same ratio as for the reaction mixture and incubated for 24 h. Aliquots were collected from time to time and analyzed using the ABTS protocol. Recovered activity of the enzymes was calculated as the ratio between detected activity at the specific time and the initial activity.
3.4. Analysis Method
The reacted mixture prepared as indicated previously (Section 3.3 Preparation of the Reaction Mixture) was analyzed by the HPLC-DAD method using a 1260 Infinity HPLC modular system from Agilent Technologies equipped with a Poroshell 120 EC-C18 column and a diode array-type detector (DAD). The experimental parameters of the analysis were as follows: acetonitrile and water (75/25, v/v) mixture as mobile phase, 0.8 mL/min flow rate, and 10 μL volume of injection. The chromatogram was recorded for 30 min (retention time of 6.2 min for (+)-α-pinene, 22.4 min for verbenol and 17.2 min for verbenone). The recorded signals were read at the specific wavelength for each analyte as follows: λα-pinene = 227 nm, λverbenol = 210 nm, and λverbenone = 260 nm. Quantitative analysis of the reacted mixture allowed to calculate the (+)-α-pinene conversion (C, %), selectivity (S, %), and yield (Y, %) in the reaction products (e.g., verbenol, verbenone, myrtenol, and myrtenal).
The GC–MS system (Thermo Electron Scientific Corporation, Waltham, MA, USA) containing a TR-WAX capillary column was used for the identification of the reaction products (e.g., verbenol, verbenone, myrtenal, and myrtenal). Helium was the carrier gas (1 mL min−1), while menthol was used as internal standard. The injection temperature was set to 200 °C and the column temperature varied with 5 °C min−1 (from 50 to 230 °C).
4. Conclusions
A cascade biocatalytic system with a bi-enzymatic biocatalyst was developed for (+)-α-pinene oxidation specific to the verbenol product. The biocatalyst cocktail containing the 2-1B peroxidase enzyme (a versatile peroxidase mutant) and M120 laccase separately catalyzed the steps of the cascade process. The peroxidase was responsible for the allylic oxidation of (+)-α-pinene mostly to verbenol, whereas laccase assisted in the oxidation of the alcoholic unit of verbenol to the corresponding ketone (verbenone). In this way, the laccase enzyme supported the catalytic activity of peroxidase. Therefore, a cooperative connection was established between 2-1B peroxidase and M120 laccase, thereby increasing the performance of the overall oxidation process. The conclusion was confirmed by kinetic measurements.
Optimization of the reaction conditions (e.g., the concentrations of 2-1B peroxidase, M120 laccase, and H2O2) allowed to achieve the maximum conversion of 80% (+)-α-pinene with 68% and 20% selectivity in verbenol and verbenone, respectively. Myrtenol and myrtenal were also detected in the products’ mixture, but with low abundance (<10% selectivity in myrtenol/myrtenal). Regarding the reaction time, 5 h were enough under optimum experimental conditions to reach 67% yield in verbenol. Over the same time, good recovery of both enzymes’ activity was noticed (88% and 95% recovery of the catalytic activity for 2-1B peroxidase and M120 laccase, respectively).
The setup system is a promising alternative for (+)-α-pinene valorization due to several advantages: (i) combining the catalytic activities of two oxidoreductase enzymes based on a cooperative mechanism; (ii) using a peroxide-resistant enzyme (2-1B peroxidase) to avoid the catalytic inhibition by H2O2; (iii) producing verbenol with high efficiency (around 70% yield in verbenol); and (iv) providing an example of allylic oxidation in line with the rules of green chemistry.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2073-4344/11/1/134/s1, Figure S1: Stability tests for 2-1B peroxidase, M120 laccase, and their mixture. Experimental conditions: 1.29 U/mL 2-1B peroxidase and/or 216 POU/mL M120 laccase, and 1.28 M H2O2 in acetate buffer (0.1 M, pH = 5) at 40 °C and 1000 rpm stirring rate; ABTS protocol for the determination of the enzyme activity.
Author Contributions
Conceptualization, M.T. and V.I.P.; methodology, M.T. and C.S.; formal analysis, G.R.G., C.S., and S.I.; investigation, G.R.G., C.S., and S.I.; resources, M.T.; writing—original draft preparation, G.R.G.; writing—review and editing, M.T. and G.R.G.; supervision, M.T.; project administration, M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the PNCDI III PED project (contract no. 376PED/2020) from UEFISCDI, Bucharest, Romania.
Institutional Review Board Statement
Note applicable.
Informed Consent Statement
Note applicable.
Acknowledgments
We thank Miguel Alcalde from the Institute of Catalysis, CSIC, Madrid, Spain for supplying the 2-1B peroxidase mutant. We also offer many thanks to Fumiaki Ito from Amano Enzyme Company who kindly provided us with the M120 laccase enzyme.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Wender, P.; Mucciaro, T. A New and Practical Approach to the Synthesis. J. Am. Chem. Soc. 1992, 114, 5879–5881. [Google Scholar] [CrossRef]
- Paduch, R.; Trytek, M.; Król, S.K.; Kud, J.; Frant, M.; Szerszeń, M.K.; Fiedurek, J. Biological activity of terpene compounds produced by biotechnological methods. Pharm. Biol. 2016, 54, 1096–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, C.; da Fonseca, M. Biotransformation of terpenes. Biotechnol. Adv. 2006, 24, 134–142. [Google Scholar] [CrossRef]
- Fahlbusch, K.-G.; Hammerschmidt, F.-J.; Panten, J.; Pickenhagen, W.; Schatkowski, D.; Bauer, K.; Garbe, D.; Surburg, H. Flavors and Fragrances. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH: Weinheim, Germany, 2003; Volume 15, pp. 74–198. [Google Scholar]
- Vanek, T.; Halik, J.; Vankova, R.; Valterova, I. Formation of trans-Verbenol and Verbenone from α-Pinene Catalysed by Immobilised Picea abies Cells. Biosci. Biotechnol. Biochem. 2005, 69, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Deepthi, P.; Manish, V.; Chowdary, G. Microbial biotransformation of α-(+)-pinene by newly identified strain of Gluconobacter japonicus MTCC 12284. Int. J. Dev. Res. 2015, 5, 5270–5275. [Google Scholar]
- Golets, M. Heterogeneously Catalyzed Valorization of Monoterpenes to High Value-Added Chemicals; VMC-KBC Umeå: Umeå, Sweden, 2014. [Google Scholar]
- Canepa, A.; Herrero, E.; Crivello, M.; Eimer, G.; Casuscelli, S. H2O2 based α-pinene oxidation over Ti-MCM-41. A kinetic study. J. Mol. Catal. A Chem. 2011, 347, 1–7. [Google Scholar] [CrossRef]
- Becerra, J.-A.; Gonzalez, L.-M.; Villa, A.-L. Kinetic study of α-pinene allylic oxidation over FePcCl16-NH2-SiO2 catalyst. J. Mol. Catal. Chem. 2016, 423, 12–21. [Google Scholar] [CrossRef]
- Fdil, N.; Romane, A.; Allaoud, S.; Karim, A.; Castanet, Y.; Mortreux, A. Terpenic olefin epoxidation using metals acetylacetonates as catalysts. J. Mol. Catal. A Chem. 1996, 108, 15–21. [Google Scholar] [CrossRef]
- Maksimchuk, N.V.; Melgunov, M.S.; Chesalov, Y.A.; Bialon, J.M.; Jarzebski, A.B.; Kholdeeva, O.A. Aerobic oxidations of α-pinene over cobalt-substitued polyoxometalate supported on amino-modified mesoporous silicates. J. Catal. 2007, 246, 241–248. [Google Scholar] [CrossRef]
- Dutenhefner, P.A.R.; da Silva, M.J.; Sales, L.S.; Sousa, E.M.B.; Gusevskaya, E.V. Solvent-free liquid-phase autoxidation of monoterpenes catalyzed by sol-gel Co/SiO2. J. Mol. Catal. A Chem. 2004, 217, 139–144. [Google Scholar] [CrossRef]
- Rao, S.; Rao, R.; Agrawal, R. Enhanced production of verbenol, a highly valued food flavourant, by an intergeneric fusant strain of Aspergillus niger and Penicillium digitatum. Biotechnol. Appl. Biochem. 2003, 37, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Limberger, R.; Aleixo, A.; Neto, A.F.; Henriques, A. Bioconversion of (+)- and (−)-alpha-pinene to (+)- and (−)-verbenone by plant cell cultures of Psychotria brachyceras and Rauvolfia sellowii. Electron. J. Biotechnol. 2007, 10, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Henriksson, M.L.; Isaksson, D.; Sjodin, K.; Hogberg, H.-E.; Vanek, T.; Valterova, I. Transformation of α-Pinene Using Picea abies Suspension Culture. J. Nat. Prod. 2003, 66, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, W.; Ge, H.; Li, G.; Shen, P.; Xu, S.; Yu, B.; Zhang, J. Biocatalytic allylic hydroxylation of unsatured triterpenes and steroids by Bacillus megaterium CGMCC 1.1741. Bioorganic Chem. 2020, 99, 2–6. [Google Scholar]
- Schrader, J. Microbial Flavour Production. In Flavors and Fragrances; Springer: Berlin, Germany, 2007; pp. 507–574. [Google Scholar]
- Bell, S.; Chen, X.; Sowden, R.; Xu, F.; Williams, J.; Wong, L.-L.; Rao, Z. Molecular Recognition in (+)-α-Pinene Oxidation by Cytochrome P450cam. J. Am. Chem. Soc. 2003, 125, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, E.G.; Perez, D.G.; Duenas, F.R.; Martinez, A.; Alcalde, M. Directed evolution of a temperature, peroxide and alkaline pH tolerant versatile peroxidase. Biochem. J. 2012, 441, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Negoi, A.; Parvulescu, V.; Tudorache, M. Peroxidase-based biocatalysis in a two-phase system for allylic oxidation of α-pinene. Catal. Today 2018, 306, 199–206. [Google Scholar] [CrossRef]
- Tavanti, M.; Parmeggiani, F.; Castellanos, J.R.G.; Mattevi, A.; Turner, N.J. One-pot Biocatalytic Double Oxidation of α-Isophorone for the synthesis of Ketoisophorone. ChemCatChem 2017, 9, 3338–3348. [Google Scholar] [CrossRef]
- Espeja, P.M.; Ruiz, E.G.; Perez, D.G.; Ullrich, R.; Hofrichter, M.; Alcalde, M. Directed Evolution of Unspecific Peroxygenase from Agrocybe aegerita. Appl. Environ. Microbiol. 2014, 80, 3496–3507. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, V.S.; Acebes, S.; Ruiz, E.G.; Romero, A.; Guallar, V.; Alcalde, M.; Medrano, F.; Martinez, A.; Duenas, F.R. Unveiling the basis of alkaline stability of an evolved versatile peroxidase. Biochem. J. 2016, 473, 1917–1928. [Google Scholar] [CrossRef]
- Andreasson, L.-E.; Reinhammar, B. The Mechanism of Electron Transfer in Laccase-catalysed Reactions. Biochim. Biophys. Acta 1979, 568, 145–156. [Google Scholar] [CrossRef]
- Riva, S. Laccase: Blue enzymes for green chemistry. Trends Biotechnol. 2006, 24, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Faber, K. Cascade-Reactions. In Biotransformations in Organic Chemistry; Springer: Graz, Austria, 2018; pp. 357–369. [Google Scholar]
- Valderrama, B.; Ayala, M.; Duhalt, R.V. Suicide Inactivation of Peroxidases and the Challenge of Engineering More Robust Enzymes. Chem. Biol. 2002, 9, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Galli, C.; Madzak, C.; Vadala, R.; Jolivalt, C.; Gentili, P. Concerted Electron/Proton Transfer Mechanism in the Oxidation of Phenols by Laccase. ChemBioChem 2013, 14, 2500–2505. [Google Scholar] [CrossRef]
- Ander, P.; Messner, K. Oxidation of 1-hydroxybenzotriazole by laccase and lignin peroxidase. Biotechnol. Tech. 1998, 12, 191–195. [Google Scholar] [CrossRef]
Scheme 1.
Alternatives of α-pinene oxidation.

Figure 1.
The influence of 2-1B peroxidase on the biocatalytic cascade system. Reaction conditions: 0.63 M (+)-α-pinene, 1.28 M H2O2, 216 POU/mL M120 laccase, and 2-1B peroxidase in acetate buffer (0.1 M, pH = 5) for 24 h at 40 °C and 1000 rpm stirring rate.
Figure 1.
The influence of 2-1B peroxidase on the biocatalytic cascade system. Reaction conditions: 0.63 M (+)-α-pinene, 1.28 M H2O2, 216 POU/mL M120 laccase, and 2-1B peroxidase in acetate buffer (0.1 M, pH = 5) for 24 h at 40 °C and 1000 rpm stirring rate.

Figure 2.
The influence of M120 laccase on the biocatalytic cascade system. Reaction conditions: 0.63 M (+)-α-pinene, 1.28 M H2O2, 1.29 U/mL 2-1B peroxidase, and M120 laccase in acetate buffer (0.1 M, pH = 5) for 24 h at 40 °C and 1000 rpm stirring rate.
Figure 2.
The influence of M120 laccase on the biocatalytic cascade system. Reaction conditions: 0.63 M (+)-α-pinene, 1.28 M H2O2, 1.29 U/mL 2-1B peroxidase, and M120 laccase in acetate buffer (0.1 M, pH = 5) for 24 h at 40 °C and 1000 rpm stirring rate.

Figure 3.
The influence of H2O2 on the biocatalytic cascade system. Reaction conditions: 0.63 M (+)-α-pinene, 1.29 U/mL 2-1B peroxidase, 216 POU/mL M120 laccase, and H2O2 in acetate buffer (0.1 M, pH = 5) for 24 h at 40 °C and 1000 rpm stirring rate.
Figure 3.
The influence of H2O2 on the biocatalytic cascade system. Reaction conditions: 0.63 M (+)-α-pinene, 1.29 U/mL 2-1B peroxidase, 216 POU/mL M120 laccase, and H2O2 in acetate buffer (0.1 M, pH = 5) for 24 h at 40 °C and 1000 rpm stirring rate.

Figure 4.
Monitoring the biocatalytic cascade transformation of (+)-α-pinene in time. Reaction conditions: 0.63 M (+)-α-pinene, 1.29 U/mL 2-1B peroxidase, 216 POU/mL M120 laccase, and 1.28 M H2O2 in acetate buffer (0.1 M, pH = 5) at 40 °C and 1000 rpm stirring rate.
Figure 4.
Monitoring the biocatalytic cascade transformation of (+)-α-pinene in time. Reaction conditions: 0.63 M (+)-α-pinene, 1.29 U/mL 2-1B peroxidase, 216 POU/mL M120 laccase, and 1.28 M H2O2 in acetate buffer (0.1 M, pH = 5) at 40 °C and 1000 rpm stirring rate.

Scheme 2.
Proposed biocatalytic route of the (+)-α-pinene biotransformation leading to verbenol as principal product and verbenone as favored by-product in the bi-enzymatic cascade system; the hypothetical mechanism of the cascade process is also indicated.
Scheme 2.
Proposed biocatalytic route of the (+)-α-pinene biotransformation leading to verbenol as principal product and verbenone as favored by-product in the bi-enzymatic cascade system; the hypothetical mechanism of the cascade process is also indicated.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Kinetic/catalytic characterization of 2-1B peroxidase, M120 laccase, and their mixture.
| Enzyme(s) | Km (μM) | kcat (s−1) | kcat/Km (s−1 mM−1) |
|---|---|---|---|
| 2-1B peroxidase | 23 ± 5 | 0.122 ± 0.05 | 5.30 ± 0.92 |
| M120 laccase | 580 ± 73 | 0.526 ± 0.07 | 0.91 ± 0.09 |
| 2-1B peroxidase in the presence of M120 laccase | 65 ± 3 | 0.991± 0.04 | 15.25 ± 5.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gheorghita, G. R.; Sora, C.; Ion, S.; Parvulescu, V. I.; Tudorache, M. Cascade Biocatalysis Designed for the Allylic Oxidation of α-Pinene. Catalysts 2021, 11, 134. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11010134
AMA Style
Gheorghita GR, Sora C, Ion S, Parvulescu VI, Tudorache M. Cascade Biocatalysis Designed for the Allylic Oxidation of α-Pinene. Catalysts. 2021; 11(1):134. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11010134
Chicago/Turabian StyleGheorghita, Giulia R., Cristina Sora, Sabina Ion, Vasile I. Parvulescu, and Madalina Tudorache. 2021. "Cascade Biocatalysis Designed for the Allylic Oxidation of α-Pinene" Catalysts 11, no. 1: 134. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11010134
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.