
Enhanced In Vitro Cascade Catalysis of Glycerol into Pyruvate and Acetoin by Integration with Dihydroxy Acid Dehydratase from Paralcaligenes ureilyticus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
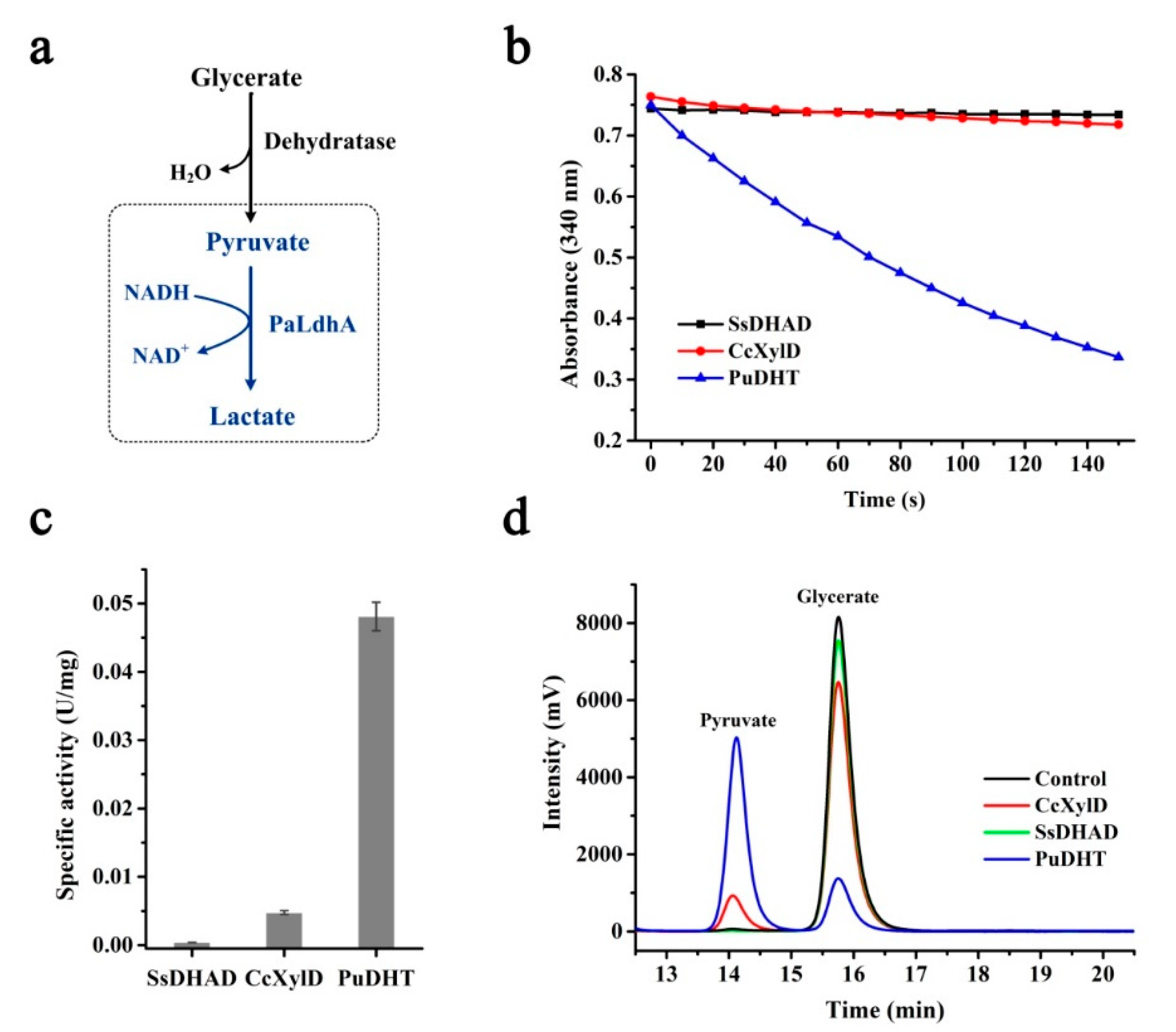
2.1. Design and Application of the Dehydratase Determination System
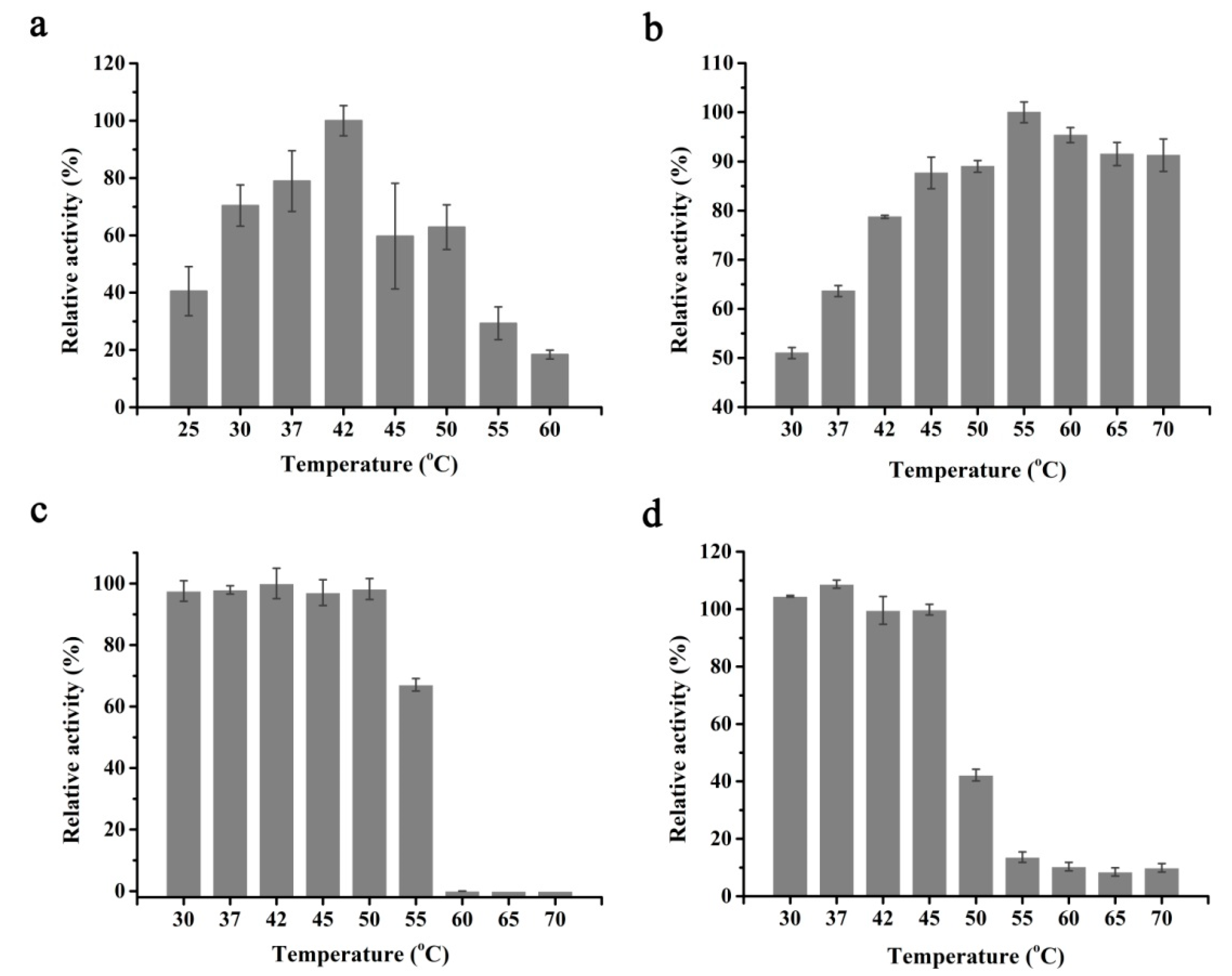
2.2. Selection of the Suitable Reaction Temperature
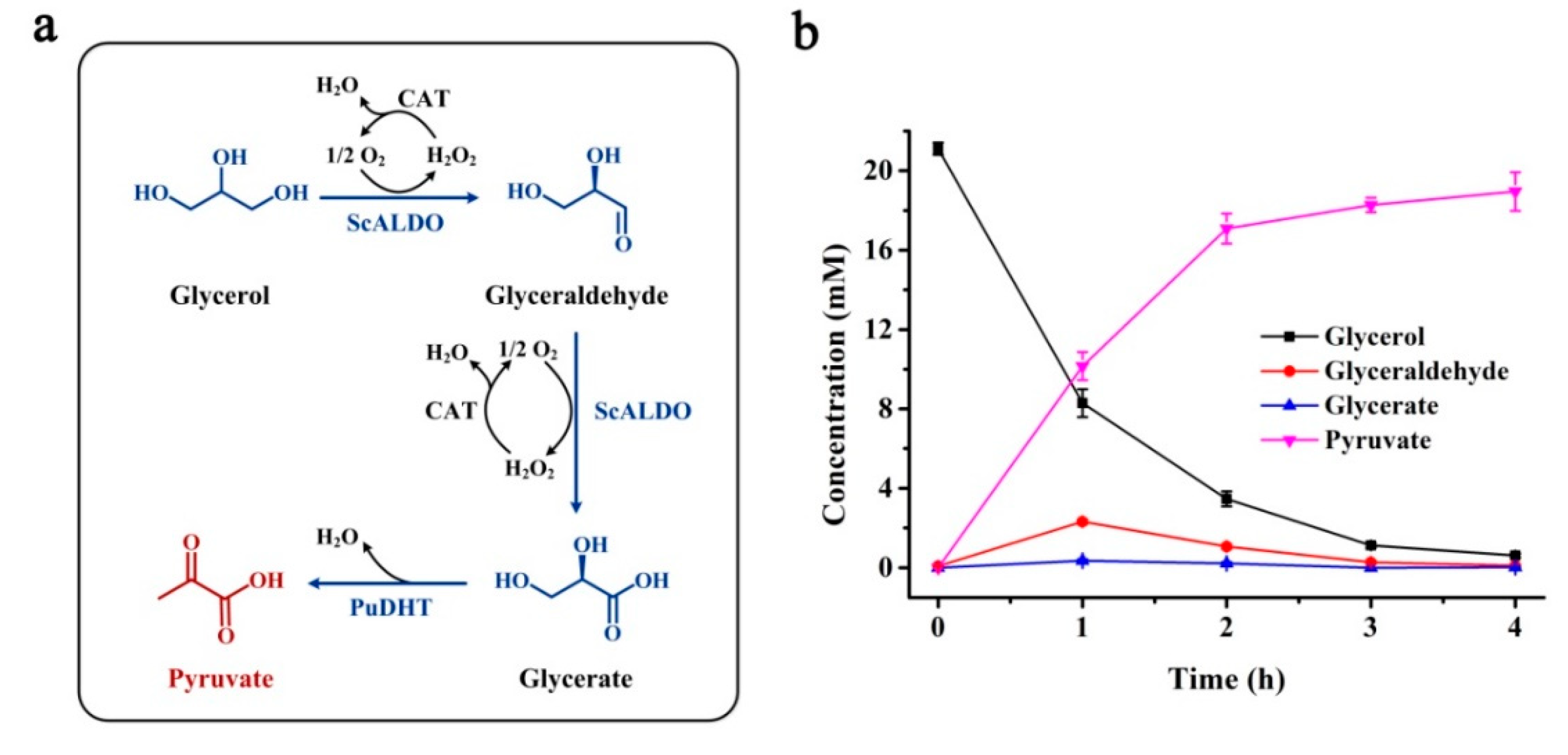
2.3. Production of Pyruvate from Glycerol by In Vitro Enzymatic Cascade with PuDHT
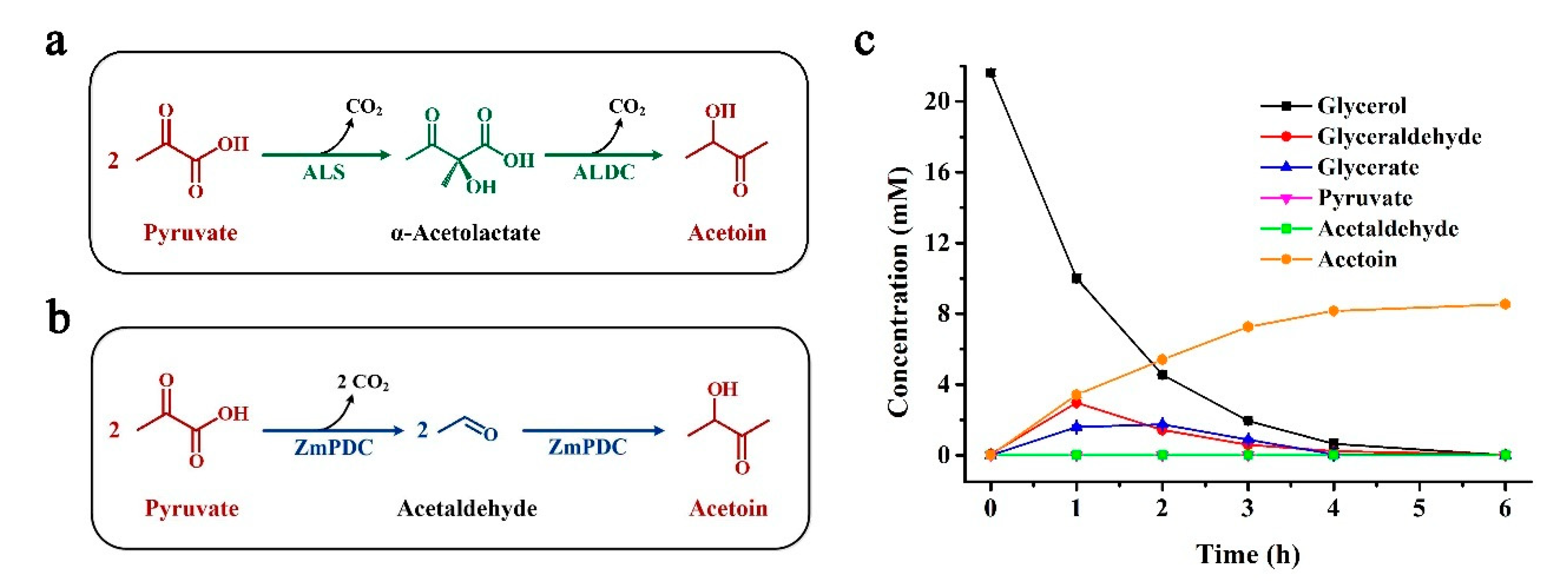
2.4. Production of Acetoin from Glycerol by In Vitro Enzymatic Cascade Coupled with ZmPDC
3. Materials and Methods
3.1. Materials
3.2. Strains and Plasmids
3.3. Expression and Purification of Enzymes
3.4. Dihydroxy Acid Dehydratase Activity Analysis
3.5. Identification of the Effects of Temperature on Activities and Stabilities of ScALDO and PuDHT
3.6. Production Pyruvate and Acetoin from Glycerol
3.7. Analytical Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Makris, D.P.; Lalas, S. Glycerol and glycerol-based deep eutectic mixtures as emerging green solvents for polyphenol extraction: The evidence so far. Molecules 2020, 25, 5842. [Google Scholar] [CrossRef]
- Rahim, S.A.N.M.; Lee, C.S.; Abnisa, F.; Aroua, M.K.; Daud, W.A.W.; Cognet, P.; Pérès, Y. A review of recent developments on kinetics parameters for glycerol electrochemical conversion—A by-product of biodiesel. Sci. Total Environ. 2020, 705, 135137. [Google Scholar] [CrossRef]
- Kumar, L.R.; Yellapu, S.K.; Tyagi, R.D.; Zhang, X. A review on variation in crude glycerol composition, bio-valorization of crude and purified glycerol as carbon source for lipid production. Bioresour. Technol. 2019, 293, 122155. [Google Scholar] [CrossRef]
- Wang, Y.; Furukawa, S.; Song, S.; He, Q.; Asakura, H.; Yan, N. Catalytic production of alanine from waste glycerol. Angew. Chem. Int. Ed. Engl. 2020, 59, 2289–2293. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.J.; Lim, E.J.; Kim, Y.; Noh, Y.; Huber, G.W.; Kim, W.B. Highly selective transformation of glycerol to dihydroxyacetone without using oxidants by a PtSb/C-catalyzed electrooxidation process. Green Chem. 2016, 18, 2877–2887. [Google Scholar] [CrossRef]
- Murakami, N.; Oba, M.; Iwamoto, M.; Tashiro, Y.; Noguchi, T.; Bonkohara, K.; Abdel-Rahman, M.A.; Zendo, T.; Shimoda, M.; Sakai, K.; et al. l-Lactic acid production from glycerol coupled with acetic acid metabolism by Enterococcus faecalis without carbon loss. J. Biosci. Bioeng. 2016, 121, 89–95. [Google Scholar] [CrossRef]
- Kaur, J.; Sarma, A.K.; Jha, M.K.; Gera, P. Valorisation of crude glycerol to value-added products: Perspectives of process technology, economics and environmental issues. Biotechnol. Rep. (Amst.) 2020, 27, e00487. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Goyal, A.; Moholkar, V.S. Microbial conversion of glycerol: Present status and future prospects. Crit. Rev. Biotechnol. 2012, 32, 235–262. [Google Scholar] [CrossRef]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.C.; Zheng, Y.G.; Shen, Y.C. Use of glycerol for producing 1,3-dihydroxyacetone by Gluconobacter oxydans in an airlift bioreactor. Bioresour. Technol. 2011, 102, 7177–7182. [Google Scholar] [CrossRef]
- Dikshit, P.K.; Moholkar, V.S. Optimization of 1,3-dihydroxyacetone production from crude glycerol by immobilized Gluconobacter oxydans MTCC 904. Bioresour. Technol. 2016, 216, 1058–1065. [Google Scholar] [CrossRef]
- Przystałowska, H.; Lipiński, D.; Słomski, R. Biotechnological conversion of glycerol from biofuels to 1,3-propanediol using Escherichia coli. Acta. Biochim. Pol. 2015, 62, 23–34. [Google Scholar] [CrossRef]
- Yu, Y.; Zhu, X.; Xu, H.; Zhang, X. Construction of an energy-conserving glycerol utilization pathways for improving anaerobic succinate production in Escherichia coli. Metab. Eng. 2019, 56, 181–189. [Google Scholar] [CrossRef]
- Ge, Y.; Li, K.; Li, L.; Gao, C.; Zhang, L.; Ma, C.; Xu, P. Contracted but effective: Production of enantiopure 2, 3-butanediol by thermophilic and GRAS Bacillus licheniformis. Green Chem. 2016, 18, 4693–4703. [Google Scholar] [CrossRef]
- Olson, D.G.; Sparling, R.; Lynd, L.R. Ethanol production by engineered thermophiles. Curr. Opin. Biotechnol. 2015, 33, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Akita, H.; Nakashima, N.; Hoshino, T. Production of d-lactate using a pyruvate-producing Escherichia coli strain. Biosci. Biotechnol. Biochem. 2017, 81, 1452–1455. [Google Scholar] [CrossRef] [Green Version]
- Cybulski, K.; Tomaszewska-Hetman, L.; Rakicka, M.; Juszczyk, P.; Rywińska, A. Production of pyruvic acid from glycerol by Yarrowia lipolytica. Folia. Microbiol. (Praha) 2019, 64, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Kamzolova, S.V.; Morgunov, I.G. Biosynthesis of pyruvic acid from glycerol-containing substrates and its regulation in the yeast Yarrowia lipolytica. Bioresour. Technol. 2018, 266, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.; Yovkova, V.; Aurich, A.; Mauersberger, S.; Barth, G. Variation of the by-product spectrum during α-ketoglutaric acid production from raw glycerol by overexpression of fumarase and pyruvate carboxylase genes in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2012, 95, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Madzak, C.; Du, G.; Zhou, J.; Chen, J. Effects of pyruvate dehydrogenase subunits overexpression on the α-ketoglutarate production in Yarrowia lipolytica WSH-Z06. Appl. Microbiol. Biotechnol. 2014, 98, 7003–7012. [Google Scholar] [CrossRef]
- Zeng, W.; Zhang, H.; Xu, S.; Fang, F.; Zhou, J. Biosynthesis of keto acids by fed-batch culture of Yarrowia lipolytica WSH-Z06. Bioresour. Technol. 2017, 243, 1037–1043. [Google Scholar] [CrossRef]
- Gao, C.; Li, Z.; Zhang, L.; Wang, C.; Li, K.; Ma, C.; Xu, P. An artificial enzymatic reaction cascade for a cell-free bio-system based on glycerol. Green Chem. 2015, 17, 804–807. [Google Scholar] [CrossRef]
- Carsten, J.M.; Schmidt, A.; Sieber, V. Characterization of recombinantly expressed dihydroxy-acid dehydratase from Sulfobus solfataricus-A key enzyme for the conversion of carbohydrates into chemicals. J. Biotechnol. 2015, 211, 31–41. [Google Scholar] [CrossRef]
- Belluzo, M.S.; Ribone, M.É.; Lagier, C.M. Assembling amperometric biosensors for clinical diagnostics. Sensors 2008, 8, 1366–1399. [Google Scholar] [CrossRef] [Green Version]
- Gilis, M.; Durliat, H.; Comtat, M. Amperometric biosensors for l-alanine and pyruvate assays in biological fluids. Anal. Chim. Acta. 1997, 355, 235–240. [Google Scholar] [CrossRef]
- Yu, Z.; Huang, H.; Reim, A.; Charles, P.D.; Northage, A.; Jackson, D.; Parry, I.; Kessler, B.M. Optimizing 2D gas chromatography mass spectrometry for robust tissue, serum and urine metabolite profiling. Talanta 2017, 165, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Grenache, D.G. Development of an automated enzymatic method to quantify pyruvate kinase in red blood cells. J. Appl. Lab. Med. 2020, 5, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, N.; Miyanaga, A.; Togawa, M.; Nakajima, M.; Taguchi, H. Diverse allosteric and catalytic functions of tetrameric d-lactate dehydrogenases from three Gram-negative bacteria. AMB Express 2014, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, N.; Miyanaga, A.; Nakajima, M.; Taguchi, H. Structural basis of sequential allosteric transitions in tetrameric d-lactate dehydrogenases from three Gram-negative bacteria. Biochemistry 2018, 57, 5388–5406. [Google Scholar] [CrossRef]
- Andberg, M.; Aro-Kärkkäinen, N.; Carlson, P.; Oja, M.; Bozonnet, S.; Toivari, M.; Hakulinen, N.; O’Donohue, M.; Penttilä, M.; Koivula, A. Characterization and mutagenesis of two novel iron-sulphur cluster pentonate dehydratases. Appl. Microbiol. Biotechnol. 2016, 100, 7549–7563. [Google Scholar] [CrossRef]
- Sutiono, S.; Teshima, M.; Beer, B.; Schenk, G.; Sieber, V. Enabling the direct enzymatic dehydration of d-glycerate to pyruvate as the key step in synthetic enzyme cascades used in the cell-free production of fine chemicals. ACS Catal. 2020, 10, 3110–3118. [Google Scholar] [CrossRef]
- Switala, J.; Loewen, P.C. Diversity of properties among catalases. Arch. Biochem. Biophys. 2002, 401, 145–154. [Google Scholar] [CrossRef]
- Fiedurek, J.; Gromada, A. Production of catalase and glucose oxidase by Aspergillus niger using unconventional oxygenation of culture. J. Appl. Microbiol. 2000, 89, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Lü, C.; Ge, Y.; Cao, M.; Guo, X.; Liu, P.; Gao, C.; Xu, P.; Ma, C. Metabolic engineering of Bacillus licheniformis for production of acetoin. Front. Bioeng. Biotechnol. 2020, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Lu, J.R. Strategies for enhancing fermentative production of acetoin: A review. Biotechnol. Adv. 2014, 32, 492–503. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, M.; Jiang, T.; Sheng, B.; Ma, C.; Xu, P.; Gao, C. Enzymatic cascades for efficient biotransformation of racemic lactate derived from corn steep water. ACS Sustainable Chem. Eng. 2017, 5, 3456–3464. [Google Scholar] [CrossRef]
- Zhu, Y.; Eiteman, M.A.; Altman, R.; Altman, E. High glycolytic flux improves pyruvate production by a metabolically engineered Escherichia coli strain. Appl. Environ. Microbiol. 2008, 74, 6649–6655. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Chen, Q.; Fan, F.; Tang, J.; Zhan, T.; Wang, H.; Zhang, X. Directed evolution of alditol oxidase for the production of optically pure d-glycerate from glycerol in the engineered Escherichia coli. J. Ind. Microbiol. Biotechnol. 2021, 48, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Zhu, J.; Xiao, D.; Xu, P.; Ma, C.; Gao, C.; Lü, C. Coculture of Gluconobacter oxydans and Escherichia coli for 3, 4-dihydroxybutyric acid production from xylose. ACS Sustainable Chem. Eng. 2021, 9, 10809–10817. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, S.; Tan, X.; Wang, Y.; Li, K.; Lü, C.; Ma, C.; Gao, C. Enhanced In Vitro Cascade Catalysis of Glycerol into Pyruvate and Acetoin by Integration with Dihydroxy Acid Dehydratase from Paralcaligenes ureilyticus. Catalysts 2021, 11, 1282. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11111282
Guo S, Tan X, Wang Y, Li K, Lü C, Ma C, Gao C. Enhanced In Vitro Cascade Catalysis of Glycerol into Pyruvate and Acetoin by Integration with Dihydroxy Acid Dehydratase from Paralcaligenes ureilyticus. Catalysts. 2021; 11(11):1282. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11111282
Chicago/Turabian StyleGuo, Shiting, Xiaoxu Tan, Yuxian Wang, Kai Li, Chuanjuan Lü, Cuiqing Ma, and Chao Gao. 2021. "Enhanced In Vitro Cascade Catalysis of Glycerol into Pyruvate and Acetoin by Integration with Dihydroxy Acid Dehydratase from Paralcaligenes ureilyticus" Catalysts 11, no. 11: 1282. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11111282