
A Thermophilic GH5 Endoglucanase from Aspergillus fumigatus and Its Synergistic Hydrolysis of Mannan-Containing Polysaccharides

Abstract
:1. Introduction
2. Results
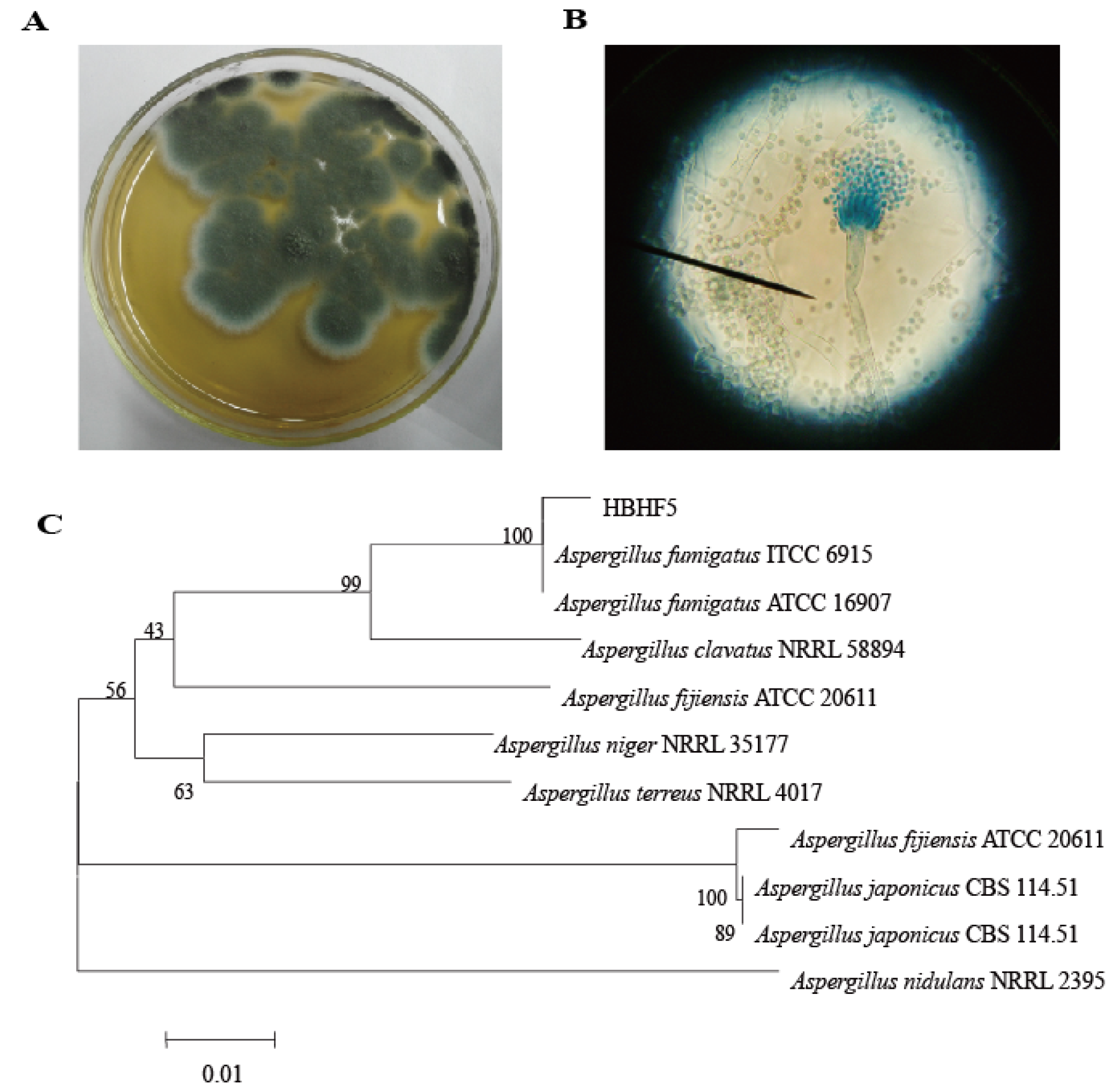
2.1. Isolation and Identification of the Thermophilic Fungi
2.2. Transcriptome Analysis of A. fumigatus HBHF
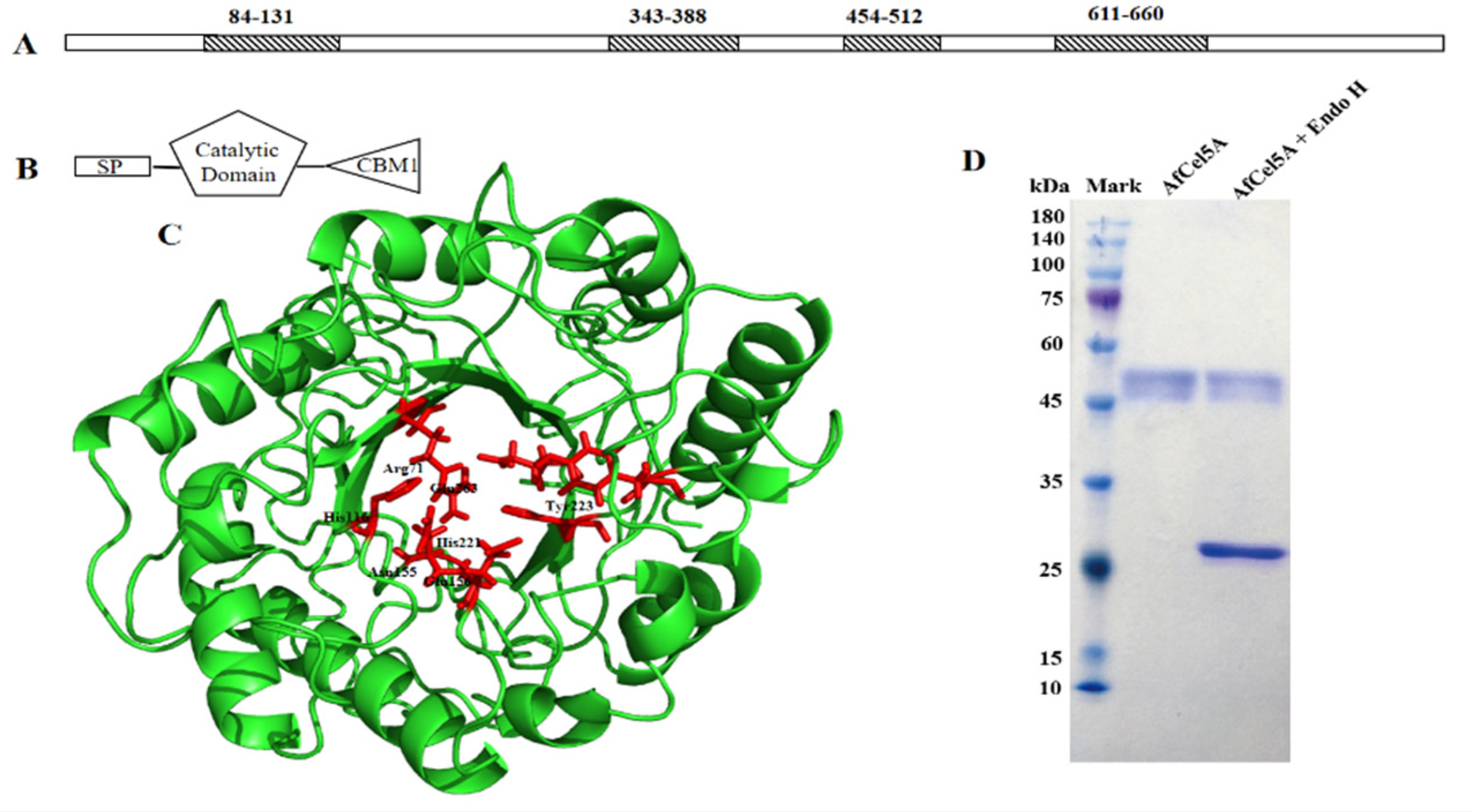
2.3. Sequence Analysis, Expression and Purification of AfCel5A
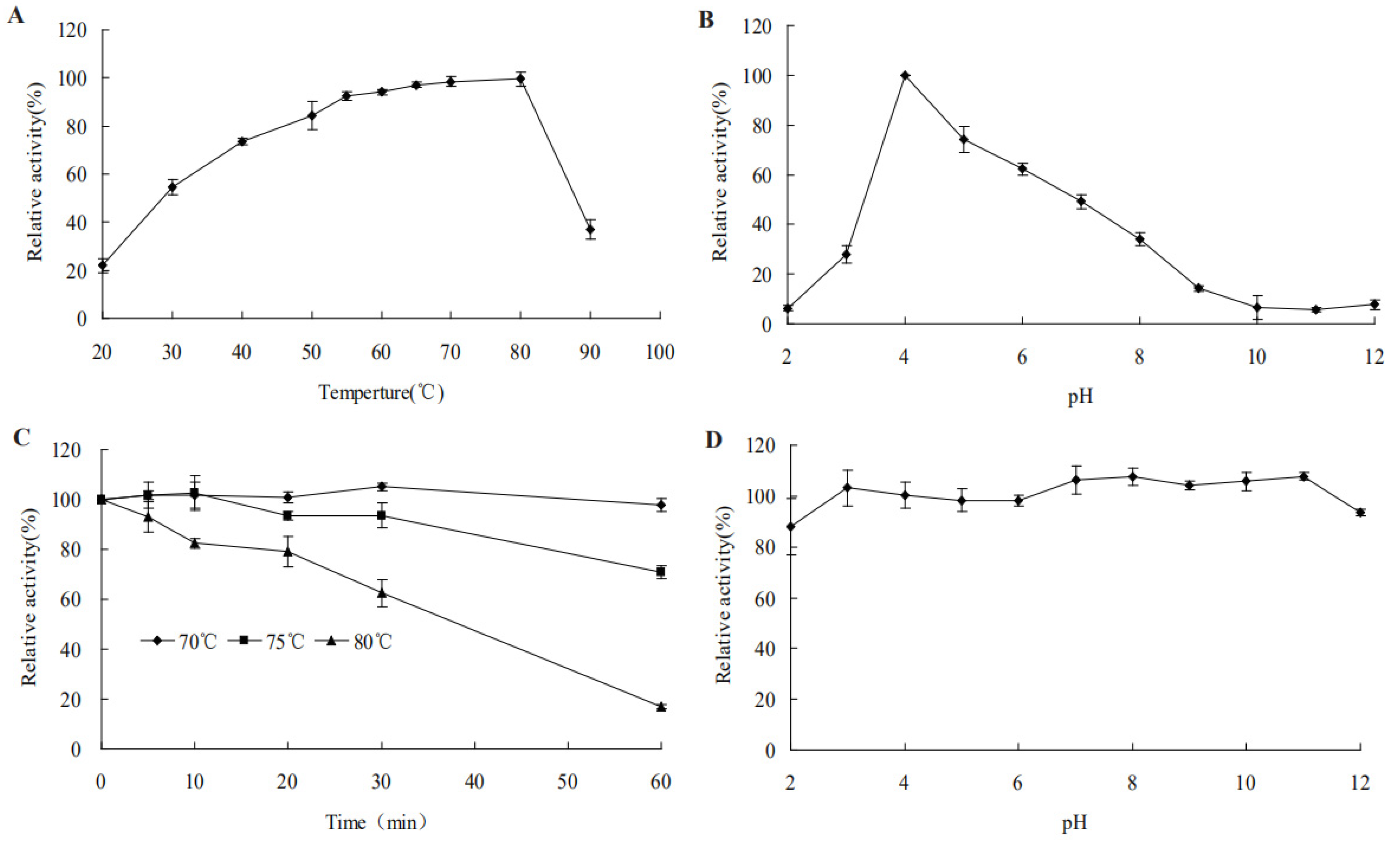
2.4. Characteristics of Recombinant AfCel5A
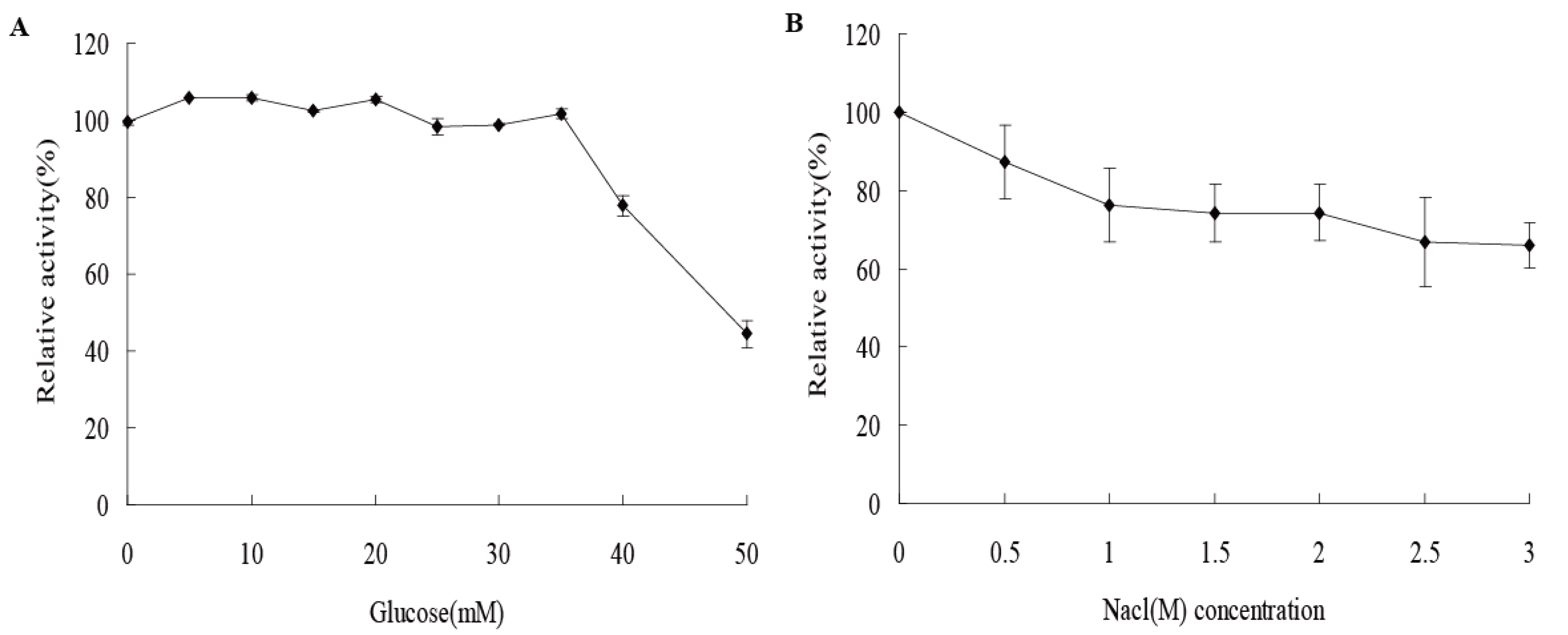
2.5. Activity of AfCel5A in a High Concentration of NaCl and Glucose
2.6. Synergistic Action of AfCel5A and ManBK on Mannan Degradation
3. Discussion
4. Materials and Methods
4.1. Strain and Culture Medium
4.2. Microorganism Isolation and Identification
4.3. RNA Isolation and Sequencing Analysis
4.4. Cloning and Bioinformatic Analysis of AfCel5A Gene
4.5. Heterologous Expression and Purification of AfCel5
4.6. Determination of Enzymatic Activity
4.7. Biochemical Characterization
4.8. Effect of NaCl and Glucose on Activity
4.9. Synergistic Hydrolysis of Mannans by AfCel5 and ManBK
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanderson, K. Lignocellulose: A chewy problem. Nature 2011, 474, S12–S14. [Google Scholar] [CrossRef]
- Arevalo-Gallegos, A.; Ahmad, Z.; Asgher, M.; Parra-Saldivar, R.; Iqbal, H.M.N. Lignocellulose: A sustainable material to produce value-added products with a zero waste approach—A review. Int. J. Biol. Macromol. 2017, 99, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Zhao, Y.; Pan, S.; Wang, X.; Dong, C. A comparative life cycle analysis of wheat straw utilization modes in China. Energy 2020, 194, 116914. [Google Scholar] [CrossRef]
- Liqiang, C.; Noerpel, M.R.; Scheckel, K.G.; Ippolito, J.A. Wheat straw biochar reduces environmental cadmium bioavailability. Environ. Int. 2019, 126, 69–75. [Google Scholar] [CrossRef]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef]
- Oke, M.A.; Annuar, M.S.M.; Simarani, K. Enhanced endoglucanase production by Bacillus aerius on mixed lignocellulosic substrates. BioResources 2016, 11, 5854–5869. [Google Scholar] [CrossRef] [Green Version]
- Akram, F.; ul Haq, I.; Imran, W.; Mukhtar, H. Insight perspectives of thermostable endoglucanases for bioethanol production: A review. Renew. Energ. 2018, 122, 225–238. [Google Scholar] [CrossRef]
- Siqueira, J.G.W.; Rodrigues, C.; de Souza Vandenberghe, L.P.; Woiciechowski, A.L.; Soccol, C.R. Current advances in on-site cellulase production and application on lignocellulosic biomass conversion to biofuels: A review. Biomass Bioenerg. 2020, 132, 105419. [Google Scholar] [CrossRef]
- Ahmed, A.; Bibi, A. Fungal cellulase; production and applications: Minireview. Int. J. Health Life Sci. 2018, 4, 19–36. [Google Scholar] [CrossRef] [Green Version]
- Behera, B.C.; Sethi, B.K.; Mishra, R.R.; Dutta, S.K.; Thatoi, H.N. Microbial cellulases–Diversity & biotechnology with reference to mangrove environment: A review. J. Genet. Eng. Biotechnol. 2017, 15, 197–210. [Google Scholar] [CrossRef]
- Maleki, M.; Shahraki, M.F.; Kavousi, K.; Ariaeenejad, S.; Salekdeh, G.H. A novel thermostable cellulase cocktail enhances lignocellulosic bioconversion and biorefining in a broad range of pH. Int. J. Biol. Macromol. 2020, 154, 349–360. [Google Scholar] [CrossRef]
- Atalah, J.; Cáceres-Moreno, P.; Espina, G.; Blamey, J.M. Thermophiles and the applications of their enzymes as new biocatalysts. Bioresour. Technol. 2019, 280, 478–488. [Google Scholar] [CrossRef]
- Sahoo, K.; Sahoo, R.K.; Gaur, M.; Subudhi, E. Cellulolytic thermophilic microorganisms in white biotechnology: A review. Folia Microbiol. 2020, 65, 25–43. [Google Scholar] [CrossRef]
- Mohammad, B.T.; Al Daghistani, H.I.; Jaouani, A.; Abdel-Latif, S.; Kennes, C. Isolation and characterization of thermophilic bacteria from Jordanian hot springs: Bacillus licheniformis and Thermomonas hydrothermalis isolates as potential producers of thermostable enzymes. Int. J. Microbiol. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Liu, B.; Zhang, X. Characterization of a recombinant thermostable xylanase from deep-sea thermophilic Geobacillus sp. MT-1 in East Pacific. Appl. Microbiol. Biotechnol. 2006, 72, 1210–1216. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, H.; Shi, P.; Luo, H.; Wang, Y.; Yang, P.; Yao, B. A family 5 β-mannanase from the thermophilic fungus Thielavia arenaria XZ7 with typical thermophilic enzyme features. Appl. Microbiol. Biotechnol. 2013, 97, 8121–8128. [Google Scholar] [CrossRef]
- Zheng, X.W.; Tabrizi, M.R.; Nout, M.R.; Han, B.Z. Daqu—A traditional Chinese liquor fermentation starter. J. Inst. Brew. 2011, 117, 82–90. [Google Scholar] [CrossRef]
- Yao, S.; Liu, Y.; Zhang, M.; Zhang, X.; Li, H.; Zhao, T.; Xin, C.; Xu, L.; Zhang, B.; Cheng, C. Thermoactinomyces daqus sp. nov., a thermophilic bacterium isolated from high-temperature Daqu. Int. J. Syst. Evol. Microbiol. 2014, 64, 206–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Yi, Z.; Fang, Y.; Jin, Y.; Xiao, Y.; Zhao, D.; Huo, H.; He, H.; Sun, Q.; Zhao, H. Uncovering key residues responsible for the thermostability of a thermophilic 1, 3 (4)-β-d-glucanase from Nong flavor Daqu by rational design. Enzym. Microb. Tech. 2020, 142, 109672. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, G.; Arya, S.K. Mannans: An overview of properties and application in food products. Int. J. Biol. Macromol. 2018, 119, 79–95. [Google Scholar] [CrossRef]
- Moreira, L.R.S. An overview of mannan structure and mannan-degrading enzyme systems. Appl. Microbiol. Biotechnol. 2008, 79, 165–178. [Google Scholar] [CrossRef]
- Malgas, S.; van Dyk, J.S.; Pletschke, B.I. A review of the enzymatic hydrolysis of mannans and synergistic interactions between β-mannanase, β-mannosidase and α-galactosidase. World J. Microb. Biotechnol. 2015, 31, 1167–1175. [Google Scholar] [CrossRef]
- Jeon, S.D.; Yu, K.O.; Kim, S.W.; Han, S.O. A celluloytic complex from Clostridium cellulovorans consisting of mannanase B and endoglucanase E has synergistic effects on galactomannan degradation. Appl. Microbiol. Biotechnol. 2011, 90, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Várnai, A.; Huikko, L.; Pere, J.; Siika-Aho, M.; Viikari, L. Synergistic action of xylanase and mannanase improves the total hydrolysis of softwood. Bioresour. Technol. 2011, 102, 9096–9104. [Google Scholar] [CrossRef] [PubMed]
- De Gouvêa, P.F.; Bernardi, A.V.; Gerolamo, L.E.; de Souza Santos, E.; Riaño-Pachón, D.M.; Uyemura, S.A.; Dinamarco, T.M. Transcriptome and secretome analysis of Aspergillus fumigatus in the presence of sugarcane bagasse. BMC Genom. 2018, 19, 1–18. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, R.; Yang, X.; Zhang, Z.; Song, S.; Miao, Y.; Shen, Q. Characterization of a thermostable β-glucosidase from Aspergillus fumigatus Z5, and its functional expression in Pichia pastoris X33. Microb. Cell Fact. 2012, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yilmazer, C.; Germec, M.; Turhan, I. Solid-state fermentation for the production of a recombinant β-mannanase from Aspergillus fumigatus expressed in Aspergillus sojae grown on renewable resources. J. Food Process. Preserv. 2020, e14584. [Google Scholar] [CrossRef]
- Zehra, M.; Syed, M.N.; Sohail, M. Banana peels: A promising substrate for the coproduction of pectinase and xylanase from Aspergillus fumigatus MS16. Pol. J. Microbiol. 2020, 69, 19. [Google Scholar] [CrossRef] [Green Version]
- Dave, B.R.; Sudhir, A.P.; Subramanian, R.B. Purification and properties of an endoglucanase from Thermoascus aurantiacus. Biotechnol. Rep. 2015, 6, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Zheng, F.; Wang, Y.; Su, X.; Bai, Y.; Yao, B.; Huang, H.; Luo, H. Characterization of two thermophilic cellulases from Talaromyces leycettanus JCM12802 and their synergistic action on cellulose hydrolysis. PLoS ONE 2019, 14, e0224803. [Google Scholar] [CrossRef] [PubMed]
- Bruins, M.E.; Janssen, A.E.; Boom, R.M. Thermozymes and their applications. Appl. Miochem. Biotech. 2001, 90, 155–186. [Google Scholar] [CrossRef]
- Vianna Bernardi, A.; Kimie Yonamine, D.; Akira Uyemura, S.; Magnani Dinamarco, T. A thermostable Aspergillus fumigatus GH7 endoglucanase over-expressed in Pichia pastoris stimulates lignocellulosic biomass hydrolysis. Int. J. Mol. Sci. 2019, 20, 2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Jana, A.; Paul, T.; Halder, S.K.; Ghosh, K.; Maity, C.; Mohapatra, P.K.D.; Pati, B.R.R.; Mondal, K.C. Thermodynamics and kinetic properties of halostable endoglucanase from Aspergillus fumigatus ABK9. J. Basic Microbiol. 2014, 54, S142–S151. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, R.; Yang, X.; Xu, Y.; Tang, Z.; Tian, W.; Shen, Q. Expression, purification and characterization of two thermostable endoglucanases cloned from a lignocellulosic decomposing fungi Aspergillus fumigatus Z5 isolated from compost. Protein. Expres. Purif. 2011, 79, 176–186. [Google Scholar] [CrossRef]
- Meera, B.; Vanitha, M.C.; Ramani, G.; Manjula, A.; Gunasekaran, P. Cloning and Expression of an Intronless Gene Encoding Endoglucanase from Aspergillus fumigatus MKU1. Indian J Biotechnol. 2011, 10, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Rungrattanakasin, B.; Premjet, S.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. Cloning and expression of an endoglucanase gene from the thermotolerant fungus Aspergillus fumigatus DBiNU-1 in Kluyveromyces lactis. Braz. J. Microbiol. 2018, 49, 647–655. [Google Scholar] [CrossRef]
- Wang, K.; Luo, H.; Bai, Y.; Shi, P.; Huang, H.; Xue, X.; Yao, B. A thermophilic endo-1, 4-β-glucanase from Talaromyces emersonii CBS394. 64 with broad substrate specificity and great application potentials. Appl. Microbiol. Biotechnol. 2014, 98, 7051–7060. [Google Scholar] [CrossRef] [PubMed]
- Malgas, S.; van Dyk, S.J.; Pletschke, B.I. β-Mannanase (Man26A) and α-galactosidase (Aga27A) synergism–a key factor for the hydrolysis of galactomannan substrates. Enzym. Microb. Technol. 2015, 70, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Picout, D.R.; Ross-Murphy, S.B.; Jumel, K.; Harding, S.E. Pressure cell assisted solution characterization of polysaccharides. 2. Locust bean gum and tara gum. Biomacromolecules 2002, 3, 761–767. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, F.A.; Fiorentino, G.; Bartolucci, S.; Contursi, P.; Limauro, D. A thermophilic enzymatic cocktail for galactomannans degradation. Enzym. Microb. Technol. 2018, 111, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Pan, L.; He, Z.; Liu, W.; Zheng, D.; Zhang, Z.; Wang, B. A novel thermophilic β-mannanase with broad-range pH stability from Lichtheimia ramosa and its synergistic effect with α-galactosidase on hydrolyzing palm kernel meal. Process. Biochem. 2020, 88, 51–59. [Google Scholar] [CrossRef]
- Yang, L.; Shi, G.; Tao, Y.; Lai, C.; Li, X.; Zhou, M.; Yong, Q. The Increase of Incomplete Degradation Products of Galactomannan Production by Synergetic Hydrolysis of β-Mannanase and α-Galactosidase. Appl. Biochem. Biotech. 2021, 193, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Lu, Z.M.; Zhang, X.J.; Wang, S.T.; Ao, L.; Shen, C.H.; Shi, J.S.; Xu, Z.H. Bio-heat is a key environmental driver shaping the microbial community of medium-temperature Daqu. Appl. Environ. Microb. 2017, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAZy Family | Gene Number | Gene ID |
|---|---|---|
| CE1 | 1 | AFUA_5G09860 |
| CE4 | 5 | AFUA_4G12330, AFUA_1G10540, AFUA_2G05900, AFUA_3G00520, AFUA_4G12120 |
| CE8 | 3 | AFUA_3G07650, AFUA_8G06880, AFUA_3G07650 |
| CE9 | 1 | AFUA_8G04100 |
| CE12 | 1 | AFUA_1G03890 |
| PL1 | 2 | AFUA_2G00760, AFUA_7G06400 |
| GH1 | 2 | AFUA_2G05580, AFUA_3G06060 |
| GH5 | 2 | AFUA_6G08840, AFUA_7G01320 |
| GH13 | 6 | AFUA_1G15150, AFUA_2G13460, AFUA_3G07380, AFUA_4G10130, AFUA_5G10540, AFUA_2G00710 |
| GH15 | 3 | AFUA_2G00690, AFUA_3G00610, AFUA_4G10140 |
| GH18 | 12 | AFUA_1G02800, AFUA_3G07110, AFUA_3G07160, AFUA_3G11280, AFUA_5G01400, AFUA_5G03760, AFUA_5G03850, AFUA_7G05140, AFUA_8G01410 AFUA_5G03960, AFUA_5G06840, AFUA_8G00700, |
| GH20 | 4 | AFUA_2G00640, AFUA_8G05020, AFUA_3G11780, AFUA_8G04060 |
| GH28 | 6 | AFUA_1G17220, AFUA_4G13920, AFUA_1G17220, AFUA_8G01970, AFUA_8G02630, AFUA_8G06890 |
| GH30 | 13 | AFUA_1G05770, AFUA_1G14710, AFUA_1G16400, AFUA_1G17410, AFUA_5G07080, AFUA_5G07190, AFUA_6G03570, AFUA_6G08700, AFUA_6G12010, AFUA_7G06140, AFUA_3G00230, AFUA_5G07080, AFUA_8G07120 |
| GH32 | 3 | AFUA_2G01240, AFUA_5G00480, AFUA_6G05000 |
| GH33 | 1 | AFUA_4G13800 |
| GH35 | 7 | AFUA_1G14170, AFUA_3G00380, AFUA_5G14090, AFUA_5G14550, AFUA_6G06660, AFUA_4G00390, AFUA_1G16700 |
| GH36 | 5 | AFUA_1G01200, AFUA_5G02130, AFUA_5G13830, AFUA_8G01100, AFUA_8G01130 |
| GH54 | 2 | AFUA_1G16920, AFUA_3G02090 |
| GH55 | 6 | AFUA_1G03600, AFUA_1G14450, AFUA_6G13270, AFUA_1G11460, AFUA_3G07520, AFUA_6G09250 |
| GH62 | 2 | AFUA_1G09900, AFUA_2G15160 |
| GH63 | 5 | AFUA_1G16250, AFUA_4G10150, AFUA_8G07070, AFUA_5G03500, AFUA_6G04210 |
| GH65 | 1 | AFUA_4G13530 |
| GH74 | 8 | AFUA_2G09520, AFUA_3G03870, AFUA_5G01830, AFUA_6G01800, AFUA_6G07480, AFUA_6G11600, AFUA_7G01540, AFUA_7G06740 |
| GH75 | 4 | AFUA_3G14980, AFUA_4G01290, AFUA_6G00500, AFUA_8G00930 |
| GH92 | 5 | AFUA_1G14560, AFUA_4G10070, AFUA_6G06790, AFUA_6G12360, AFUA_3G08200 |
| GT2 | 10 | AFUA_3G10400, AFUA_5G08210, AFUA_1G12600, AFUA_2G01870, AFUA_2G13430, AFUA_2G13440, AFUA_3G14420, AFUA_4G04180, AFUA_5G00760, AFUA_8G05630 |
| GT4 | 3 | AFUA_4G13400, AFUA_5G13210, AFUA_6G06940 |
| GT5 | 1 | AFUA_5G02480 |
| GT15 | 1 | AFUA_1G06890 |
| GT20 | 6 | AFUA_6G12950, AFUA_2G04010, AFUA_3G05650, AFUA_3G07370AFUA_7G03940, AFUA_2G04020, |
| GT21 | 1 | AFUA_5G09550 |
| GT22 | 1 | AFUA_1G13870 |
| GT33 | 1 | AFUA_6G14180 |
| GT35 | 1 | AFUA_1G12920 |
| GT39 | 3 | AFUA_1G07690, AFUA_3G06450, AFUA_8G04500 |
| GT48 | 1 | AFUA_6G12400 |
| GT58 | 1 | AFUA_5G11990 |
| GT59 | 1 | AFUA_2G11080 |
| GT66 | 1 | AFUA_8G04430 |
| GT90 | 16 | AFUA_2G02360, AFUA_5G14780, AFUA_6G14480, AFUA_2G01450, AFUA_2G14910, AFUA_2G15910, AFUA_4G10750, AFUA_5G13090, AFUA_6G04450, AFUA_6G14040, AFUA_7G01300, AFUA_5G10760, AFUA_4G09130, AFUA_4G11280, AFUA_5G06050, AFUA_1G01380 |
| Metal Ions and Chemicals | Relative Activity (%) a | |
|---|---|---|
| - | 1 mM | 5 mM |
| None | 100.0 ± 1.5 | 100.0 ± 0.7 |
| Ag+ | 56.9 ± 2.7 | 13.1 ± 3.3 |
| Mn2+ | 133.4 ± 2.6 | 13.9 ± 6.9 |
| Zn2+ | 96.7 ± 1.3 | 56.3 ± 5.2 |
| Al3+ | 90.6 ± 2.3 | 79.3 ± 1.3 |
| Mg2+ | 94.8 ± 1.4 | 89.1 ± 1.4 |
| Fe3+ | 84.6 ± 1.4 | 91.6 ± 1.8 |
| Li+ | 84.75 ± 1.4 | 95.7 ± 4.1 |
| Ca2+ | 80.1 ± 1.5 | 87.2 ± 2.9 |
| K+ | 89.5 ± 3.6 | 102.7 ± 1.2 |
| Acetone | 77.5 ± 1.8 | 70.3 ± 0.6 |
| Triton X-100 | 75.5 ± 3.0 | 58.8 ± 4.6 |
| SDS | 20.9 ± 8.9 | 0.5 ± 3.4 |
| CTAB | 82.9 ± 7.5 | 65.9 ± 4.1 |
| EDTA | 90.4 ± 3.6 | 84.3 ± 6.6 |
| Urea | 97.3 ± 4.3 | 96.8 ± 3.5 |
| Enzyme Added | Reducing Sugar Content (μ mol) | Synergistic Effect (DS) | ||
|---|---|---|---|---|
| Mannan | First Enzyme | Second Enzyme | ||
| AfCel5 | None | 0.02 | - | |
| ManBK | None | 0.30 | - | |
| LBG | AfCel5 | ManBK | 0.55 | 1.57 |
| ManBK | AfCel5 | 0.54 | 1.53 | |
| AfCel5 + ManBK | None | 0.49 | 1.39 | |
| AfCel5 | None | 0.44 | - | |
| ManBK | None | 0.45 | - | |
| Guar gum | AfCel5 | ManBK | 1.06 | 1.19 |
| ManBK | AfCel5 | 1.06 | 1.19 | |
| AfCel5 + ManBK | None | 1.07 | 1.20 | |
| AfCel5 | None | 0.05 | - | |
| ManBK | None | 0.29 | - | |
| KGM | AfCel5 | ManBK | 0.59 | 1.74 |
| ManBK | AfCel5 | 0.63 | 1.61 | |
| AfCel5 + ManBK | None | 0.41 | 1.19 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, X.; Lu, H.; Zhang, L.; Meng, X. A Thermophilic GH5 Endoglucanase from Aspergillus fumigatus and Its Synergistic Hydrolysis of Mannan-Containing Polysaccharides. Catalysts 2021, 11, 862. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11070862
Gu X, Lu H, Zhang L, Meng X. A Thermophilic GH5 Endoglucanase from Aspergillus fumigatus and Its Synergistic Hydrolysis of Mannan-Containing Polysaccharides. Catalysts. 2021; 11(7):862. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11070862
Chicago/Turabian StyleGu, Xinxi, Haiqiang Lu, Lijuan Zhang, and Xiangchen Meng. 2021. "A Thermophilic GH5 Endoglucanase from Aspergillus fumigatus and Its Synergistic Hydrolysis of Mannan-Containing Polysaccharides" Catalysts 11, no. 7: 862. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11070862