
Chitosan/Selenium Nanoparticles Attenuate Diclofenac Sodium-Induced Testicular Toxicity in Male Rats

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Synthesis of Se/Chitosan
2.3. Experimental Animal Care
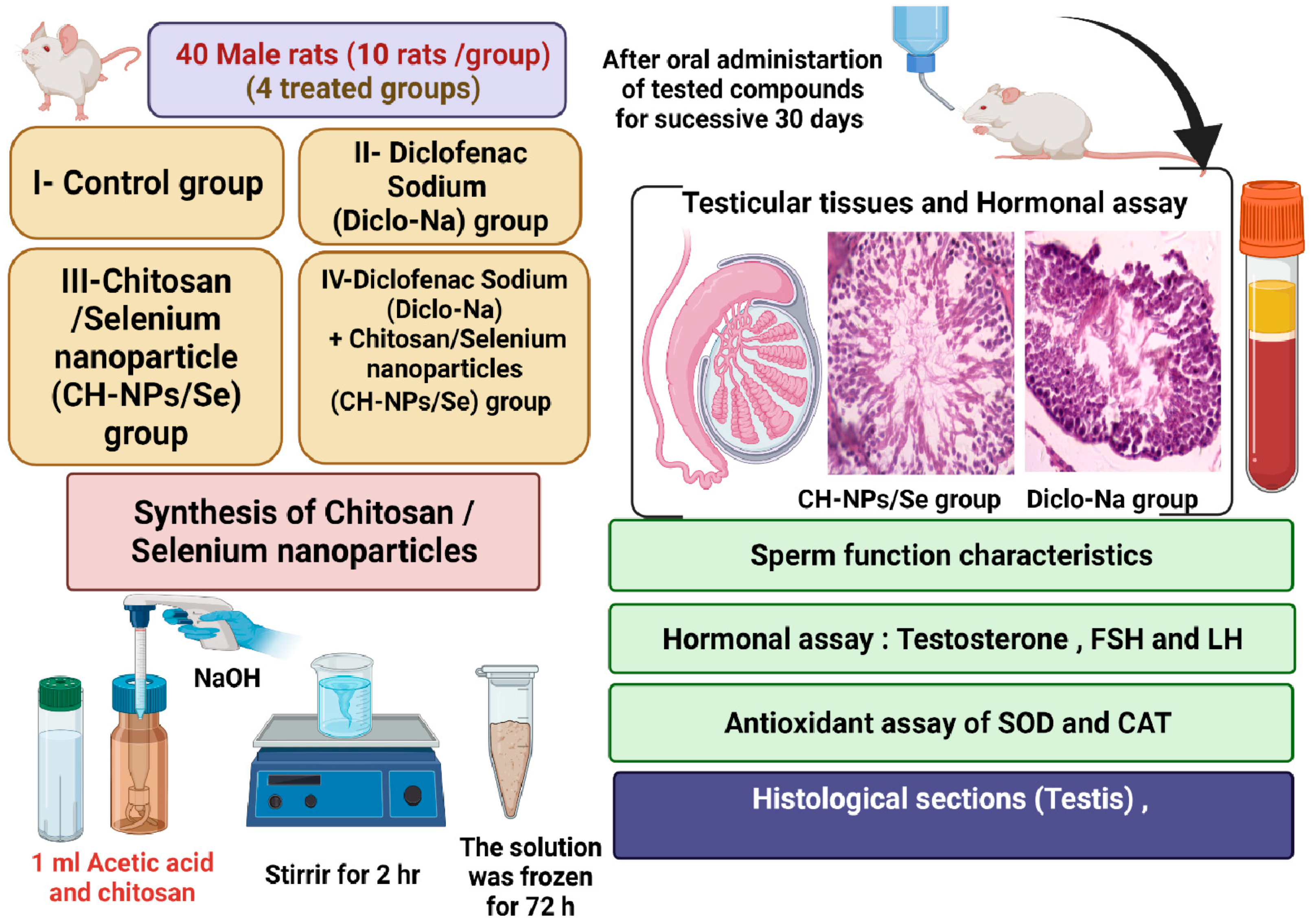
2.4. Experimental Design
2.5. Sperm Characteristics and Marker Enzymes of Testicular Function Assessment
2.6. Assessment of Testosterone Hormone
2.7. Assessment of Antioxidant and Oxidative Stress Indices
2.8. Histological Assessment
2.9. Statistical Analysis
3. Results
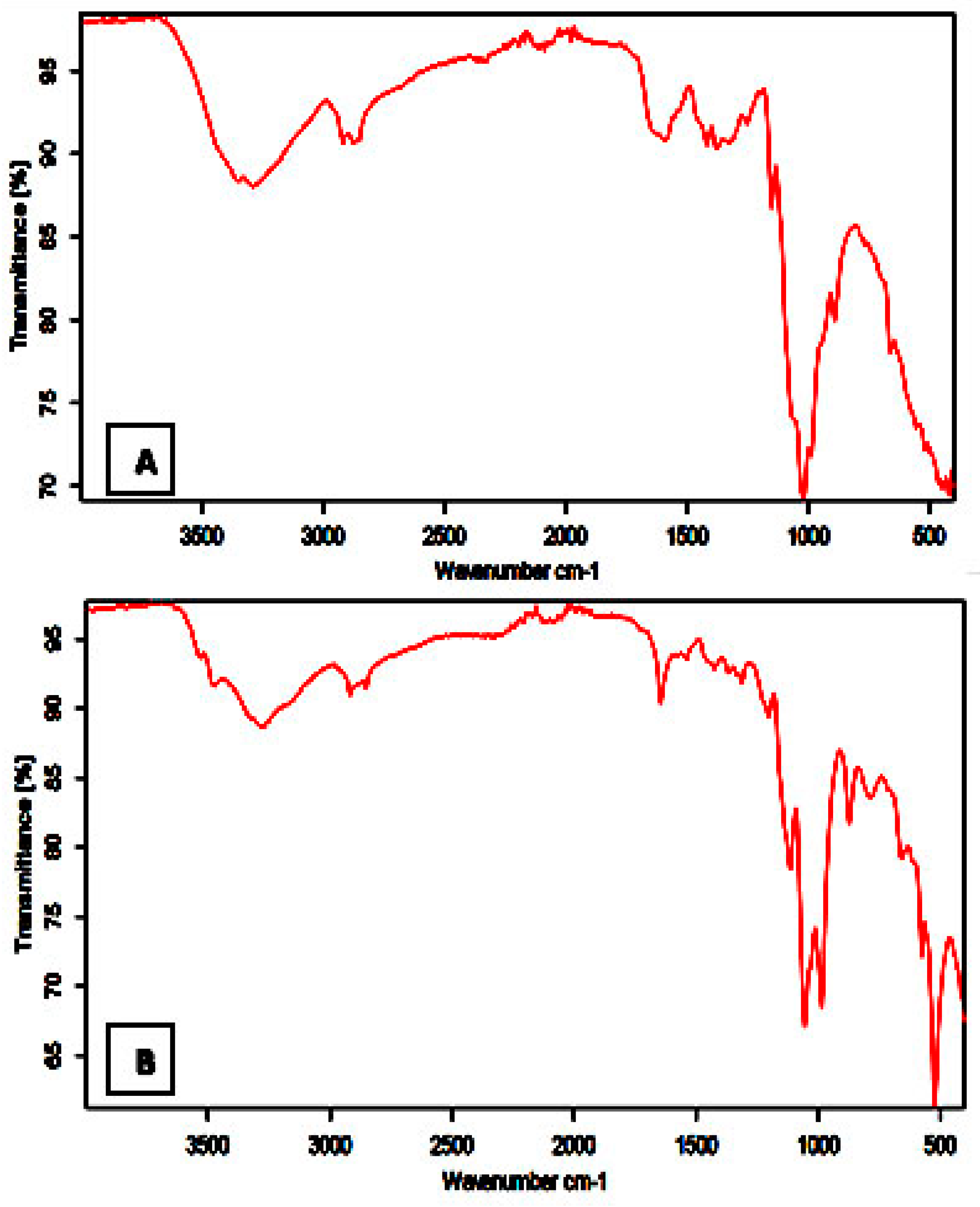
3.1. FTIR Studies
3.2. Thermal Analysis
3.3. XRD
3.4. Scanning Electron Microscopy (SEM)
3.5. Transmittance Electron Microscopy (TEM)
3.6. CH-NPs/Se Complex Improved Testosterone Hormone Levels and Sperm Characteristics in Diclo-Na Treated Rats
3.7. CH-NPs/Se Complex Alleviated Diclo-Na-Induced Oxidative Stress in the Testicular Tissue
3.8. CH-NPs/Se Complex Improved Testicular Structure in Diclo-Na-Treated Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Megharbel, S.M.; Hamza, R.Z.; Refat, M.S. Synthesis, spectroscopic and thermal studies of Mg(II), Ca(II), Sr(II) and Ba(II) diclofenac sodium complexes as anti-inflammatory drug and their protective effects on renal functions impairment and oxidative stress. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 135, 915–928. [Google Scholar] [CrossRef]
- Mousa, A.A.; Elweza, A.E.; Elbaz, H.T.; Tahoun, E.A.E.-A.; Shoghy, K.; Elsayed, I.; Hassan, E.B. Eucalyptus Globulus protects against diclofenac sodium induced hepatorenal and testicular toxicity in male rats. J. Tradit. Complement. Med. 2020, 10, 521–528. [Google Scholar] [CrossRef]
- Zhang, Y.; Geißen, S.-U.; Gal, C. Carbamazepine and diclofenac: Removal in wastewater treatment plants and occurrence in water bodies. Chemosphere 2008, 73, 1151–1161. [Google Scholar] [CrossRef]
- Ahmed, A.Y.; Gad, A.M.; El-Raouf, O.M.A. Curcumin ameliorates diclofenac sodium-induced nephrotoxicity in male albino rats. J. Biochem. Mol. Toxicol. 2017, 31, e21951. [Google Scholar] [CrossRef]
- Owumi, S.E.; Aliyu-Banjo, N.O.; Odunola, O.A. Selenium attenuates diclofenac-induced testicular and epididymal toxicity in rats. Andrologia 2020, 52, e13669. [Google Scholar] [CrossRef]
- Shintaku, K.; Hori, S.; Tsujimoto, M.; Nagata, H.; Satoh, S.; Tsukimori, K.; Nakano, H.; Fujii, T.; Taketani, Y.; Ohtani, H.; et al. Transplacental Pharmacokinetics of Diclofenac in Perfused Human Placenta. Drug Metab. Dispos. 2009, 37, 962–968. [Google Scholar] [CrossRef] [Green Version]
- Vyas, A.; Purohit, A.; Ram, H. Assessment of dose-dependent reproductive toxicity of diclofenac sodium in male rats. Drug Chem. Toxicol. 2019, 42, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Muranaka, S.; Fujita, H.; Kanno, T.; Tamai, H.; Utsumi, K. Molecular mechanism of diclofenac-induced apoptosis of promyelocytic leukemia: Dependency on reactive oxygen species, akt, bid, cytochrome and caspase pathway. Free. Radic. Biol. Med. 2004, 37, 1290–1299. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elhakeem, M.A.; Mohamed, S.R.; Aya, H.R. Chitosan nanoparticles as hepato-protective agents against alcohol and fatty diet stress in rats. J. Biochem. Int. 2017, 4, 5–10. [Google Scholar]
- Al-Baqami, N.; Hamza, R. Synergistic antioxidant capacities of vanillin and chitosan nanoparticles against reactive oxygen species, hepatotoxicity, and genotoxicity induced by aging in male Wistar rats. Hum. Exp. Toxicol. 2020, 40, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.; Al-Harbi, M.; Al-Hazaa, M. Neurological Alterations and Testicular Damages in Aging Induced by D-Galactose and Neuro and Testicular Protective Effects of Combinations of Chitosan Nanoparticles, Resveratrol and Quercetin in Male Mice. Coatings 2021, 11, 435. [Google Scholar] [CrossRef]
- Abuelzahab, H.; Hamza, R.; Montaser, M.; El-Mahdi, M.M.; Al-Harthi, W.A. Antioxidant, antiapoptotic, antigenotoxic, and hepatic ameliorative effects of L-carnitine and selenium on cadmium-induced hepatotoxicity and alterations in liver cell structure in male mice. Ecotoxicol. Environ. Saf. 2019, 173, 419–428. [Google Scholar] [CrossRef]
- Hamza, R.Z.; Al-Harbi, M.S.; El-Shenawy, N.S. Ameliorative effect of vitamin E and selenium against oxidative stress induced by sodium azide in liver, kidney, testis and heart of male mice. Biomed. Pharmacother. 2017, 91, 602–610. [Google Scholar] [CrossRef]
- Hamza, R.Z.; Diab, A.E.-A.A. Testicular protective and antioxidant effects of selenium nanoparticles on Monosodium glutamate-induced testicular structure alterations in male mice. Toxicol. Rep. 2020, 7, 254–260. [Google Scholar] [CrossRef]
- Alharthi, W.A.; Hamza, R.Z.; Elmahdi, M.M.; Abuelzahab, H.; Saleh, H. Selenium and L-Carnitine Ameliorate Reproductive Toxicity Induced by Cadmium in Male Mice. Biol. Trace Element Res. 2019, 197, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.Z.; El-Megharbel, S.M.; Altalhi, T.; Gobouri, A.A.; Alrogi, A.A. Hypolipidemic and hepatoprotective synergistic effects of selenium nanoparticles and vitamin. E against acrylamide-induced hepatic alterations in male albino mice. Appl. Organomet. Chem. 2020, 34, e5458. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Brar, S.K.; Verma, M. Green synthesis approach: Extraction of chitosan from fungus mycelia. Crit. Rev. Biotechnol. 2012, 33, 379–403. [Google Scholar] [CrossRef]
- Fernández-Llamosas, H.; Castro, L.; Blázquez, M.L.; Díaz, E.; Carmona, M. Speeding up bioproduction of selenium nanoparticles by using Vibrio natriegens as microbial factory. Sci. Rep. 2017, 7, 16046. [Google Scholar] [CrossRef] [Green Version]
- Beers, J.R.; Sizer, I.W. A Spectrophotometric Method for Measuring the Breakdown of Hydrogen Peroxide by Catalase. J. Biol. Chem. 1952, 195, 133. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of Total, Protein-Bound, and Nonprotein Sulfhydryl Groups in Tissue with Ellman’s Reagent. Anal. Biochem. 1968, 25, 192. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351. [Google Scholar] [CrossRef]
- Hayat, M.A. (Ed.) Basic Techniques for Transmission Electron Microscopy, 1st ed.; Macmillan Press: New York, NY, USA, 1986; ISBN 9780123339263. [Google Scholar]
- Dean, A.; Sullivan, K.; Soe, M. OpenEpi: Open Source Epidemiologic Statistics for Public Health. 2013. Available online: https://www.OpenEpi.com (accessed on 6 May 2013).
- Bin, J.; Xiao, Y.; Lin, Z.; Deng, Y.L.; Chen, Y.; Le, D.E.; Bin, J.; Li, M.; Liao, Y.; Liu, Y.; et al. High molecular weight chitosan derivative polymeric micelles encapsulating superparamagnetic iron oxide for tumor-targeted magnetic resonance imaging. Int. J. Nanomed. 2015, 10, 1155–1172. [Google Scholar] [CrossRef] [Green Version]
- Bujňáková, Z.; Dutková, E.; Zorkovská, A.; Baláž, M.; Kováč, J.; Kello, M.; Mojžiš, J.; Briančin, J.; Baláž, P. Mechanochemical synthesis and in vitro studies of chitosan-coated InAs/ZnS mixed nanocrystals. J. Mater. Sci. 2017, 52, 721–735. [Google Scholar] [CrossRef]
- Liu, J.; Guo, T.-F.; Shi, Y.; Yang, Y. Solvation induced morphological effects on the polymer/metal contacts. J. Appl. Phys. 2001, 89, 3668–3673. [Google Scholar] [CrossRef] [Green Version]
- Ryu, S.R.; Noda, I.; Jung, Y.M. What is the origin of positional fluctuation of spectral features: True frequency shift or relative intensity changes of two over- lapped bands? Appl. Spectrosc. 2010, 64, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, F.; Bwatanglang, I.B.; Yusof, N.A.; Abdullah, J.; Hussein, M.Z.; Alitheen, N.B.M.; Abu, N. Folic acid targeted Mn:ZnS quantum dots for theranostic applications of cancer cell imaging and therapy. Int. J. Nanomed. 2016, 11, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Lassoued, A.; Dkhil, B.; Gadri, A.; Ammar, S. Control of the shape and size of iron oxide (α-Fe2O3) nanoparticles synthesized through the chemical precipitation method. Results Phys. 2017, 7, 3007–3015. [Google Scholar] [CrossRef]
- Alagiri, M.; Hamid, S.B.A. Green synthesis of α-Fe2O3 nanoparticles for photocatalytic application. J. Mater. Sci. Mater. Electron. 2014, 25, 3572–3577. [Google Scholar] [CrossRef]
- Christen, V.; Hickmann, S.; Rechenberg, B.; Fent, K. Highly active human pharmaceuticals in aquatic systems: A concept for their identification based on their mode of action. Aquat. Toxicol. 2010, 96, 167–181. [Google Scholar] [CrossRef]
- Sofikitis, N.; Giotitsas, N.; Tsounapi, P.; Baltogiannis, D.; Giannakis, D.; Pardalidis, N. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol. 2008, 109, 323–330. [Google Scholar] [CrossRef]
- Ramaswamy, S.; Weinbauer, G.F. Endocrine control of spermatogenesis: Role of FSH and LH/testosterone. Spermatogenesis 2014, 4, e996025. [Google Scholar] [CrossRef] [PubMed]
- O’Shaughnessy, P.J. Hormonal control of germ cell development and spermatogenesis. Semin. Cell Dev. Biol. 2014, 29, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.E.; Lopez, K.H. Chapter 4—Human Reproductive Biology, 4th ed.; Elsevier Science Publishing Co. Inc.: San Diego, CA, USA, 2014. [Google Scholar]
- Liang, J.; Wang, N.; He, J.; Du, J.; Guo, Y.; Li, L.; Wu, W.; Yao, C.; Li, Z.; Kee, K. Induction of Sertoli-like cells from human fibroblasts by NR5A1 and GATA4. eLife 2019, 8, e48767. [Google Scholar] [CrossRef]
- Akhigbe, R.; Ajayi, A. Testicular toxicity following chronic codeine administration is via oxidative DNA damage and up-regulation of NO/TNF-alpha and caspase 3 activities. PLoS ONE 2020, 15, e0224052. [Google Scholar] [CrossRef] [Green Version]
- Motawi, T.M.; Sadik, N.A.; Refaat, A. Cytoprotective effects of DL-alpha-lipoic acid or squalene on cyclophosphamide-induced oxidative injury: An experimental study on rat myocardium, testicles and urinary bladder. Food Chem. Toxicol. 2010, 48, 2326–2336. [Google Scholar] [CrossRef]
- Aly, H.A.; Khafagy, R.M. Taurine reverses endosulfan-induced oxidative stress and apoptosis in adult rat testis. Food Chem. Toxicol. 2014, 64, 1–9. [Google Scholar] [CrossRef]
- Adedara, I.A.; Abolaji, A.O.; Awogbindin, I.O.; Farombi, E.O. Suppression of the brain-pituitary-testicular axis function following acute arsenic and manganese co-exposure and withdrawal in rats. J. Trace Elements Med. Biol. 2017, 39, 21–29. [Google Scholar] [CrossRef]
- AlBasher, G.; Abdel-Daim, M.M.; Almeer, R.; Ibrahim, K.; Hamza, R.Z.; Bungau, S.; Aleya, L. Synergistic antioxidant effects of resveratrol and curcumin against fipronil-triggered oxidative damage in male albino rats. Environ. Sci. Pollut. Res. 2019, 27, 6505–6514. [Google Scholar] [CrossRef]
- AdeyemiJulius, W.J.; Omoniyi, J.A.; Olayiwola, A.; Ibrahim, M.; Ogunyemi, O.; Olayaki, L.A. Elevated reproductive toxicity effects of diclofenac after withdrawal: Investigation of the therapeutic role of melatonin. Toxicol. Rep. 2019, 6, 571–577. [Google Scholar]
- Obeys, A.K.; Karim, A.; Mahood, S. Histological study of the effect of piroxicam on testes of albino mice Mus musculus. J. Univ. Anbar. Pure Sci. 2013, 7, 1–11. [Google Scholar]
- Frungieri, M.B. Cyclooxygenase and prostaglandins in somatic cell populations of the testis. Bioscientifica 2015, 149, R169–R180. [Google Scholar] [CrossRef] [Green Version]
- Sapori, P.M.H.; Chatelain, P.; Saez, J.M. In vitro interaction between Sertoli cells and steroidogenic cells. Biochem. Biophys. Res. Commun. 1986, 134, 957–962. [Google Scholar] [CrossRef]
- Papadakis, V.; Vlachopapadopoulu, W.; Van Syckle, K. Gonadal function in young patients successfully treated for Hodgkin’s disease. Med. Pediater. Oncol. 1999, 32, 366–372. [Google Scholar] [CrossRef]
- Campion, S. Male reprotoxicity and endocrine disruption. In Molecular, Clinical and Environmental Toxicology. Experientia Supplementum; Luch, A., Ed.; Springer International Publishing AG: Berlin, Germany, 2012; Volume 101, pp. 315–360. [Google Scholar]
- Mohan, D.; Sharma, S. Histopathological alterations in liver of mice exposed to different doses of diclofenac sodium. World Acad. Sci. Eng. Technol. Int. J. Anim. Vet. Sci. 2017, 11, 698–702. [Google Scholar]
- Araujo, J.M.; Fortes-Silva, R.; Pola, C.C.; Yamamoto, F.Y.; Gatlin III, D.M.; Gomes, C.L. Delivery of selenium using chitosan nanoparticles: Synthesis, characterization, and antioxidant and growth effects in Nile tilapia (Orechromis niloticus). PLoS ONE 2021, 16, e0251786. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Weight Loss 5% | Weight Loss 10% | Weight Loss 20% | Residue % | Temperature of 1st Derivative Weight Loss |
|---|---|---|---|---|---|
| Chitosan | 47.00 °C | 71.98 °C | 298.00 °C | 21.08% | 310.00 °C |
| Se/Chitosan | 65.00 °C | 93.00 °C | 277.00 °C | 25.95% | 300.00 °C |
| Title | Control | Diclo-Na | CH-NPs/Se | Diclo-Na + CH-NPs/Se |
|---|---|---|---|---|
| LH (ng/mL) | 0.15 ± 0.01 | 0.10 ± 0.04 * | 0.16 ± 0.03 | 0.13 ± 0.03 ** |
| FSH (ng/mL) | 0.80 ± 0.04 | 0.72 ± 0.01 | 0.82 ± 0.01 | 0.78 ± 0.01 |
| Testosterone (ng/mL) | 2.35 ± 0.02 | 1.02 ± 0.08 * | 2.34 ± 0.01 | 2.06 ± 0.05 ** |
| Title | Control | Diclo-Na | CH-NPs/Se | Diclo-Na + CH-NPs/Se |
|---|---|---|---|---|
| Count | 98 ± 7.01 | 81.65 ± 6.05 * | 99.28 ± 9.06 | 90.85 ± 5.33 ** |
| Viability | 96.11 ± 4.26 | 90.11 ± 5.43 | 96.90 ± 3.55 | 93.44 ± 4.23 |
| Motility | 91.00 ± 6.44 | 68.98 ± 4.28 * | 91.15 ± 8.08 | 84.87 ± 5.13 ** |
| Abnormality | 4.68 ± 0.85 | 8.94 ± 1.06 | 4.79 ± 0.87 | 5.74 ± 0.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Megharbel, S.M.; Al-Salmi, F.A.; Al-Harthi, S.; Alsolami, K.; Hamza, R.Z. Chitosan/Selenium Nanoparticles Attenuate Diclofenac Sodium-Induced Testicular Toxicity in Male Rats. Crystals 2021, 11, 1477. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11121477
El-Megharbel SM, Al-Salmi FA, Al-Harthi S, Alsolami K, Hamza RZ. Chitosan/Selenium Nanoparticles Attenuate Diclofenac Sodium-Induced Testicular Toxicity in Male Rats. Crystals. 2021; 11(12):1477. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11121477
Chicago/Turabian StyleEl-Megharbel, Samy M., Fawziah A. Al-Salmi, Sarah Al-Harthi, Khadeejah Alsolami, and Reham Z. Hamza. 2021. "Chitosan/Selenium Nanoparticles Attenuate Diclofenac Sodium-Induced Testicular Toxicity in Male Rats" Crystals 11, no. 12: 1477. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11121477