
Interplay of Nanoparticle Properties during Endocytosis


1
State Key Laboratory of Nonlinear Mechanics and Beijing Key Laboratory of Engineered Construction and Mechanobiology, Institute of Mechanics, Chinese Academy of Sciences, Beijing 100190, China
2
School of Engineering Science, University of Chinese Academy of Sciences, Beijing 100049, China
*
Authors to whom correspondence should be addressed.
Crystals 2021, 11(7), 728; https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11070728
Submission received: 30 May 2021
/
Revised: 11 June 2021
/
Accepted: 21 June 2021
/
Published: 24 June 2021
(This article belongs to the Special Issue Selected Papers from the 2nd International Online Conference on Crystals)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Nanoparticles (NPs) have been widely applied as drug carriers in drug delivery, due to their unique physical and structural properties. To achieve the drug delivery purpose, receptor-mediated endocytosis is a primary explored mechanism to internalize NPs into tumor cells. During the endocytosis process, properties of NPs, including size, shape, and surface functionality, play an important role in determining the final drug delivery efficacy. Many of these NP properties have been extensively explored individually. However, the multiple NP properties naturally interplay with each other in the endocytosis process to determine the internalization efficiency together. Therefore, it is significantly important to understand the interplay of different NP properties to improve the NP’s final delivery efficacy. In this review, we focus on the interplay of NPs properties on the endocytosis process to summarize the relevant experimental observations and physical mechanisms. Particularly, three different aspects are discussed in detail, including the interplay between size and shape; size and elasticity; shape and elasticity. We have summarized the most recent works and highlighted that building up systematic understandings for the complex interplay between NP properties can greatly help a better design of NP platforms for drug delivery.
1. Introduction
Nanoparticles (NPs) have been widely used to deliver drugs by virtue of their unique physical and structural properties [1,2,3,4,5]. NPs as drug delivery vehicles allow an efficient drug accumulation and precise payload dispensing at the targeted sites, and thus, hold great promise to improve therapeutic index [6,7,8]. For example, utilization of NPs decorated with tLyp1 peptide is shown to enhance tumor inhibition and regression in vivo, elevating the survival ratio to 40% after 60 days in a melanoma mouse model [8,9]. Generally, drug-loaded NPs can translocate from their site of deposition to the disease sites by the blood circulation system after intravenous injection [10,11]. When NPs reach the site of action, tumor cells can take up these engineered NPs and unload drug molecules at the desired intracellular compartment. The cellular uptake of NPs can significantly elevate the drug concentration and act as an intracellular reservoir for long-term drug release [12,13]. However, there are many biological barriers during the drug delivery process to achieve such a purpose. One significant challenge for the further development of NP-based therapies is the low level of cellular uptake and drug delivery [14,15]. To this end, a detailed understanding of the NP cellular uptake mechanisms is crucial for efficient therapeutic applications.
Receptor-mediated endocytosis is a primary route exploited in drug delivery to deliver the NPs and drug molecules into tumor cells [16,17]. In general, receptor-ligand complexes form when mobile receptors on the cell membrane diffuse to bind ligands on the NP’s surface. Binding between receptors and ligands drives engulfing to overcome resistance to membrane deformation and changes in configuration entropy of receptors, thus contributing to membrane wrapping and cellular uptake [18,19,20,21,22,23]. Depending on the type of involved proteins, the receptor-mediated endocytosis can be classified as clathrin-mediated endocytosis, caveolae-mediated endocytosis, and calthrin/caveolae-independent endocytosis [16,24,25,26]. In addition to the receptor-mediated endocytosis, NPs can also be internalized by cells via pinocytosis and phagocytosis, depending on the cargo type, internalization route, and scission mechanism [26]. Pinocytosis is commonly involved in the internalization of fluids and molecules from the external environment of a cell by small vesicles called pinosomes, and is ubiquitous to almost any eukaryotic cell [27]. Phagocytosis is an actin-dependent process by which the specialized cells, such as macrophages, monocytes, dendritic cells, and neutrophils, engulf large particulate matter with the formation of vesicles called phagosomes [28]. In contrast to receptor-mediated endocytosis, both pinocytosis and phagocytosis are non-selective modes of taking up foreign materials [26].
Accumulating evidence indicates that NP uptake through receptor-mediated endocytosis depends strongly on their physical and chemical characteristics, including the size, shape, stiffness, and surface functionality [17,29,30,31,32]. For instance, it has been well recognized that there is an optimal size of NP during the endocytosis process, at which the cellular uptake reaches a maximum in a cell type-dependent manner [33,34,35]. Chithrani et al. investigated the size-dependent uptake of gold NPs by Hela cells, showing that the NPs with a diameter of 50 nm were taken up into cells more effectively than those with diameters of 14, 30, 74, and 100 nm [36]. Win and Feng found that the 100 nm polystyrene NPs showed the most efficient cellular uptake compared with other sizes (50, 200, 500, and 1000 nm) [37]. Gao et al. developed a mathematical framework to uncover the mechanism underlying the size effect. They found that the optimal particle diameter is determined by the competition between receptor diffusion kinetics and thermodynamic driving force. For NPs larger than the optimal size, receptors need to diffuse longer distances, leading to increased wrapping time. For NPs smaller than the optimal size, it needs to overcome more elastic bending energy of membrane, resulting in the decreased driving force for wrapping, hence decreased efficiency of cellular uptake [38]. Apart from size, the effects of the other properties of NP (such as shape, stiffness, and surface functionality) on the cellular uptake have also been extensively investigated, and discussed in detail in many excellent reviews [29,39,40,41].
It has been indicated that NPs uptake by cells through endocytosis is actually determined by an intricate interplay between physicochemical particle properties. For instance, the cellular uptake of NPs depends strongly on the collaborative effects of shape and size [42,43]. Studying the contribution made by the interplay of NPs properties to the cellular uptake can not only deepen our understandings of the endocytosis and underlying mechanisms, but also provide guidelines and strategies for the NP design to improve the drug delivery efficacy. Here, we review representative works regarding the interplay of NP properties on the endocytosis process mainly from three aspects: (1) Size and shape, (2) size and elasticity, (3) shape and elasticity.
2. Size and Shape
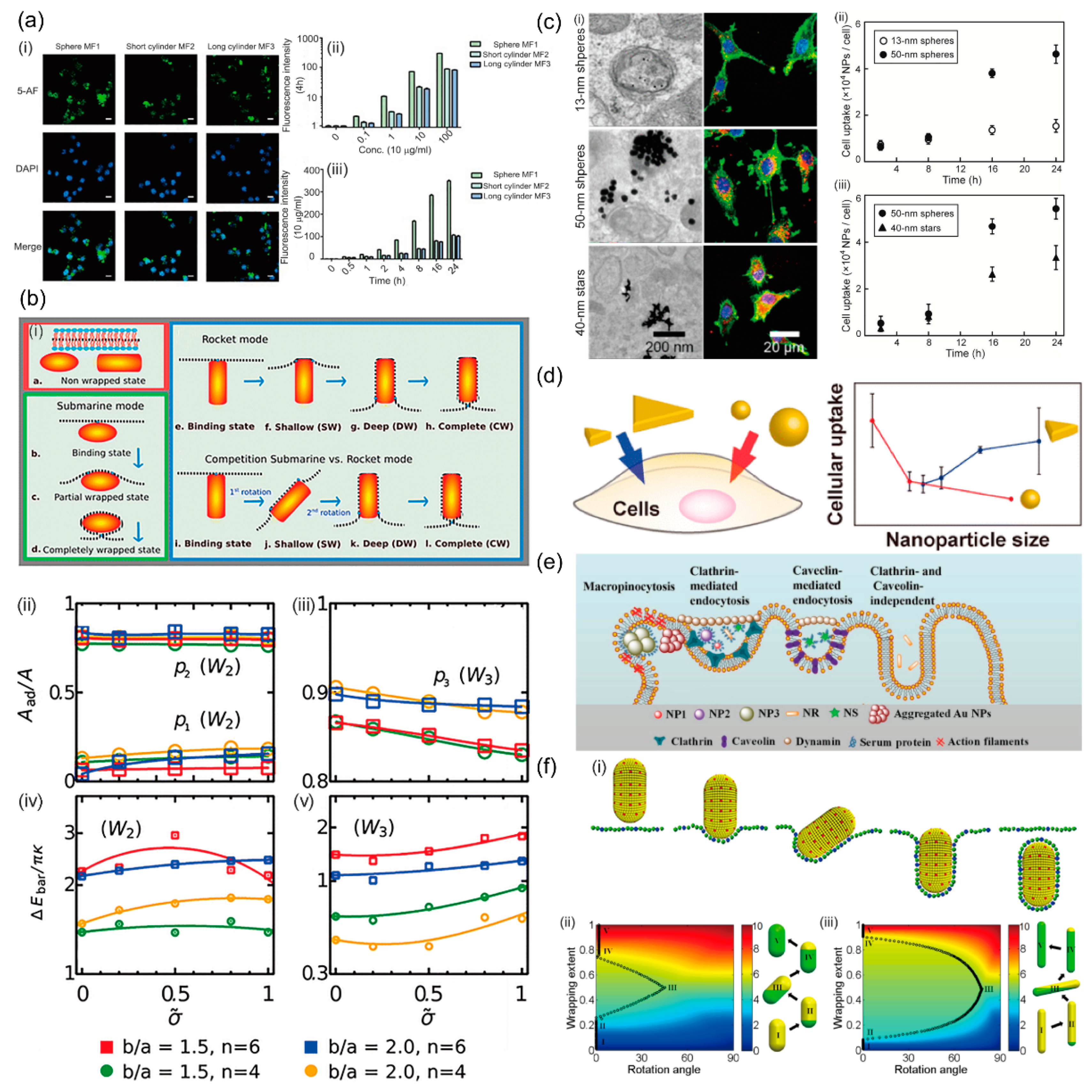
As mentioned above, size-dependent cellular uptake of NPs has been extensively investigated in various cell lines because particle size is known as a key determinant of the cellular uptake pathways; see reviews by the authors of [16,44,45] for more detail. Additionally, NPs can be of various shapes, such as spheres, triangles, stars, rods, prolate and oblate spheroids, which have been demonstrated to have a significant effect on cellular uptake of NPs [39,46,47,48]. Li et al. showed that the spherical NPs were internalized to a much greater extent than the cylindrical NPs using RAW 264.7 (Figure 1a). This difference can be attributed to their different endocytosis pathways: Uptake of spherical NPs was based on clathrin- and caveolin-mediated endocytosis, while cylindrical NPs mainly depended on clathrin-mediated endocytosis [49]. Xie et al. studied the efficiency of cellular uptake of the gold NPs with different shapes by RAW 264.7, which was found to rank as stars < rods < triangles. Further studies revealed that the various endocytosis pathways adopted by NPs are responsible for their shape-dependent uptakes [50]. Using dynamic molecular modeling combined with free energy calculations, Dasgupta et al. showed that NPs’ entry modes of membrane wrapping strongly depend on the NP shape. Their work suggested that an increased NP aspect ratio is unfavorable for complete membrane wrapping (Figure 1b). Especially, the NP enters the cell via a submarine mode for high aspect ratios and round tips and via a rocket mode for small aspect ratios and flat tips. They further showed that for cube-like and rod-like particles, the energy barriers for transitions during the wrapping process increase with the sharpness of the edges [51]. A question naturally arises, that is—how the NPs are internalized by cells if simultaneously considering the size and shape.
Yue et al. studied the effect of the structural features of NP size and shape on the ability of siRNA-conjugated gold NPs to enter U87 cells. Experimental results indicated that 50-nm spheres, and 40-nm stars have much higher cellular uptake efficiency compared to 13-nm spheres (Figure 1c). They also showed that the siRNA delivery level after 24 h of 50-nm spheres is 50-fold and 1.6-fold higher than that of 13-nm spheres and 40-nm stars, respectively, suggesting that larger NPs have higher potential as nanocarriers for the siRNA delivery. They suggested that the more efficient uptake of spheres over that of stars is caused by the differences in the interactions between oligonucleotides and scavenger receptors [43]. A more direct study to explore the interplay of size and shape of NPs was performed by Nambara et al. They investigated the uptake of triangular NPs with a side length of 46, 55, 72, and 94 nm and spherical NPs with a diameter of 22, 39, and 66 nm by RAW264.7 and HeLa cells. They found a reverse size-dependence of the cellular uptake of triangular and spherical gold NPs: Cellular uptake increases with the side length of triangular NPs, but decreases with the diameter of spherical NPs (Figure 1d). Particularly, the triangular NPs with 72 nm in side length show 20-fold more efficient uptake compared to spherical NPs with a similar surface area. They proposed that the edges and vertices of triangular NPs with high local curvature are responsible for this reverse size-dependence [42]. Furthermore, Ding et al. found that the endocytosis mechanism of gold NPs with different sizes and shapes is totally different (Figure 1e). The uptake of spherical particles with diameters of 15 and 45 nm, and rod-shaped particles is a clathrin-mediated endocytosis pathway. The internalization of star-shaped particles depends on both the clathrin-mediated and caveolin-mediated endocytosis pathways. By contrast, the uptake of spherical particles with a diameter of 80 nm is mainly through macropinocytosis, due to the bigger size [52]. There are also many computational studies exploring the interplay of NP size and shape on the endocytosis process. Using coarse-grained molecular dynamics, Huang et al. studied the receptor-mediated endocytosis of NPs, which is demonstrated to be size-dependent and shape sensitive. For spherical NPs, there is an optimal size at which cellular uptake maximizes. For the uptake of an upright spherocylindrical NP endocytosis proceeds through a laying-down-then-standing up sequence (Figure 1f). According to free energy analyses, they showed that NP size and shape affect the endocytosis in different ways: NP size primarily determines whether endocytosis can complete, while NP shape dictates the endocytic pathway and the angle of entry [53]. These results highlight the importance of the interplay between NP size and shape on their interactions with cells, and contribute to a complete understanding of cellular uptake and the design of NP platform.
3. Size and Elasticity
The mechanical property of NPs is another important factor in determining the endocytosis process, as proved by extensive experimental research [54,55,56]. However, there are often seemly controversial results regarding the effect of NP elasticity. On the one hand, Sun et al. synthesized core-shell poly (lactic-co-glycolic acid) (PLGA)-lipid NPs of varying rigidities with the same chemical composition, size, and surface properties and showed that cellular uptake of rigid NPs is more efficient than soft ones in HeLa cells (Figure 2a(i,ii)) [57]. Hui et al. studied the effect of elasticity on cancer cell uptake by synthesizing silica nanocapsules (SNCs) with a wide range of elasticity (Young’s modulus ranging from 560 kPa to 1.18 GPa) [58]. They reported a four-time higher uptake of the stiff folate-poly (ethylene glycol) (FA-PEG)-modified SNCs than their soft counterparts. Using transmission electron microscopy, they revealed that soft SNCs are deformed during endocytosis, which reduces the endocytosis rate (Figure 2a(iii)) [58].
On the other hand, Liu et al. synthesized four types of poly (2-hydroxyethyl methacrylate) (HEMA) hydrogel particles with the different compressive modulus (15–156 kPa), but similar size and surface property by an emulsion-precipitation. Their results revealed that softer NPs are internalized by HepG2 cells at a faster rate and larger amount than the stiffer NPs (Figure 2b(i)). They proposed that this difference results from the different cellular uptake mechanisms: The softer particles are internalized by cells mainly through micropinocytosis, while the stiffer particles enter cells via caveolae- and clathrin-mediated endocytosis, as well as macropinocytosis pathways [59]. Similarly, Guo et al. investigated in vitro and in vivo cellular uptake of nanolipogels (NLGs) [60]. The elasticity of NLGs is regulated by an alginate core to obtain different Young’s moduli ranging from 45 ± 9 to 19,000 ± 5 kPa. They reported that uptake of softer NLGs is significantly greater than their rigid counterparts by Neoplastic and non-neoplastic cells (Figure 2b(ii)). They also suggested that different endocytic pathways are the main reason for the different efficiencies observed in soft and rigid NPs [60].
These seemingly controversial results might be caused by the different mechanical properties of the NP itself. Particularly, soft NPs can be characterized into two different categories based on their materials: (1) Soft NPs that preserve their volume during their cellular uptake process. It is represented by lipid-based NPs [15]. The amphiphilic lipid membrane surfaces are impermeable for the water molecules inside. Therefore, the volume of lipid-based NPs is usually considered as constant during the membrane wrapping process; (2) Soft NPs that can easily change their volume. Hydrogel-based capsule NPs are representative of the second type NPs [61,62]. The mechanical properties of these NPs are maintained by the outside layer of hydrogel materials, which is permeable for the water molecules inside. Therefore, the volume of these soft capsule NPs can be easily changed during the membrane wrapping process. The difference in a volume change of NPs can significantly influence the contact area between NPs and cell membrane during the endocytosis, which further affects their endocytosis efficiency and their size effect during the cellular uptakes.
In the simulation work, Shen et al. applied the coarse-grained molecular dynamics simulations to investigate the membrane wrapping of soft NPs. In simulations, soft NPs have different bending constants and constant volume and area to regulate mechanical properties of lipid-based NPs. They first compared the membrane wrapping of soft NPs with rigid NPs. It is found that the membrane wrapping process of soft NPs is slower than rigid NPs. Further energy and kinetic analysis revealed that wrapping soft NPs needs to overcome a higher energy barrier, which requires recruiting and bind more receptors. Additionally, soft NPs and rigid NPs have a similar contact edge length with the lipid membrane, due to the controlled volume and area. The similar contact edge length induces similar receptor recruiting speeds for soft and rigid NPs. Therefore, rigid NPs are more efficient than soft NPs. They further investigated the receptor-mediated endocytosis of elastic spherical nanoparticles (NPs) with different radii (R = 25 to 100 nm) [63]. Their results showed that the minimum fully wrapped size was shifted from 27.5 nm for rigid spherical NPs to 30 nm for soft spherical, due to the increased energy barrier in soft NPs and similar contact length (Figure 2c). They further demonstrated that all soft spherical NPs are less efficient to be fully wrapped. Moreover, the wrapping efficiency difference between soft and rigid spherical NPs increased with NP sizes. Their analysis further revealed that the energy barrier of soft NPs increases as the NP size increases. However, the energy barrier of rigid NPs keeps as constant. This computational work explained the aforementioned experimental work for lipid-based NPs [64,65].
The trend of the wrapping efficiency between soft and stiff NPs in the simulations by Shen et al. seems to conflict with the theory study carried by Yi and Gao [66]. In their theoretical model, soft NPs can easily change their volume to mimic the soft elastic capsule. Due to the easy volume change, they showed that a soft NP has a larger contact area between cell membrane compared to the rigid counterparts. This large contact area can induce higher receptor recruiting speeds, which offsets the required receptors caused by the increased energy barrier. Therefore, their study indicated that the wrapping of soft NPs is kinetically faster than that of the stiff counterparts (Figure 2d). They also showed that as the particles get softer, the minimum particle size required for full internalization increases [67]. This simulation work is consistent with experimental observations on hydrogel nanoparticles [59], polymer microcapsules [68,69], in which soft NPs can be more easily deformed into a flattened configuration to enhance cell-particle interaction during the membrane wrapping process.
4. Shape and Elasticity
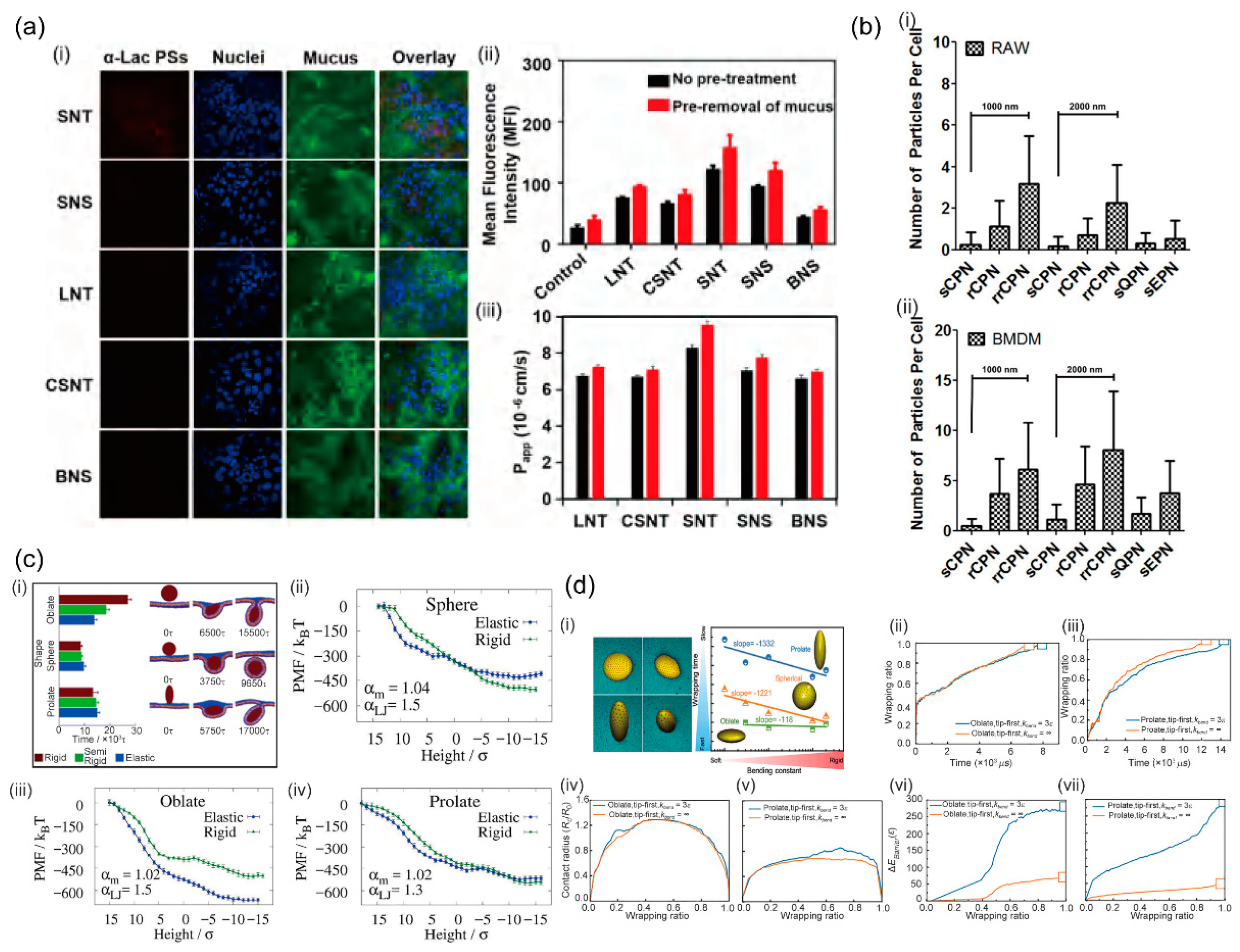
It is quite common that elastic NPs have different shapes in experiments. Thus, NP shape and elasticity may be closely intertwined in affecting the endocytosis of NPs. Based on the self-assembly of amphiphilic α-lactalbumin (α-lac) peptides from partial enzymolysis and cross-linking, Bao et al. prepared peptosomes (PSs) with various shapes and rigidities [70]. The combined experimental results and the molecular simulations demonstrated that short nanotube exhibits the highest cellular uptake and transmembrane permeability. Particularly compared with the other counterparts, including long nanotubes, big nanospheres, small nanos pheres, and cross-linked short nanotubes (Figure 3a). More importantly, their results indicated that tubular and flexible nanocarriers penetrate more efficiently through mucus than the spherical and stiffer ones, which may be attributed to their more flexible structure and the easier conformational change [70]. Palomba et al. investigated discoidal polymeric nanoconstructs (DPNs) with varying geometrical and mechanical properties to resist cellular uptake by professional phagocytic cells, including RAW 264.7 and primary cells [71]. Using confocal fluorescent microscopy (FM) and flow cytometry (FC) analysis to quantify cellular uptake of DPN, they revealed that soft circular nanoconstructs (sCPN) with a size of 1000 nm (185 ± 35 kPa) and 2000 nm (56 ± 6 kPa) are both poorly internalized compared with spherical polymeric nanoconstructs (SPNs) and ellipsoidal nanoconstructs (EPNs). While the uptake level increases by two to four times as the increase of the rigidity of rigid circular nanoconstructs (rCPNs) and rigid-rigid circular nanoconstructs (rrCPNs) (Figure 3b). Upon a critical analysis, Palomba et al. identified three different internalization regimens based on the bending stiffness ratios between particles and cells. When the particle bending stiffness is lower than cells, it facilitates internalization; while a slightly higher bending stiffness than cells is detrimental to internalization; however, when the bending stiffness is much higher than cells, it favors internalization again [71].
Utilizing a two-dimensional triangulation method, Chen et al. developed a coarse-grained model for elastically deformable NP with tunable shape and mechanical properties, wherein the interactions between NPs and membrane are described by the surface area-dependent adhesion energy with no explicit consideration of the specific types of ligands and receptors [72]. Their simulation results indicated that the effects of elasticity on the cellular uptake of deformable NPs strongly depend on their shape: The increase in softness slows down the uptakes rate of spherical and prolate NPs, but speeds up that of oblate NPs (Figure 3c). Further analyses of endocytosis mechanism and free energy calculations revealed that, due to the flexibility and the morphology deformation of soft NPs during the reorientation step, the greatly decreased curvature of oblate edge, i.e., the less membrane bending energy required for wrapping NPs, leads to a higher uptake rate compared with the stiff NPs [72]. Shen et al. investigated the receptor-mediated membrane wrapping process of elastic NPs with different shapes based on coarse-grained molecular dynamics simulations to explore the interplay between geometry and elasticity in cellular uptake of NPs [63]. In contrast to the model developed by Chen et al., the receptor diffusion kinetics has been incorporated. With comparable ligand and receptor densities as experimental values in the simulations, their results revealed three major points regarding the membrane wrappings of elastic NPs with different shapes: (1) The membrane wrapping efficiency of NPs with the same bending constant is ranked as oblates > spheres > prolates. This shape-dependent uptake is because oblate NPs have the largest contact edge lengths, followed by spherical and prolate NPs; (2) For each geometry, the wrapping time of NPs decreases with the increase in their bending constant, due to the smaller energy barrier of more rigid NPs; (3) the oblate ellipsoid is found to be the least sensitive geometry to the bending constant change, since these soft oblate NPs with significantly larger contact edge length accelerate the formation speed of receptor-ligand complexes to overcome the increased energy barrier (Figure 3d). Their results clearly showed that the membrane wrapping efficiency of NPs during receptor-mediated endocytosis depends both on receptor diffusion kinetics and thermodynamic driving force [63].
Contributions made by the interplay between NP properties to the biological performance of NPs has attracted increasing interest. In fact, besides the three combinations discussed here, there also exists interplay among other various properties of NP, which has been demonstrated to be of crucial significance for the NP uptake during the endocytosis process. For example, using confocal laser scanning microscopy, Bhattacharjee et al. investigated how the size and surface charge of NPs affect their cellular uptake and found that smaller NPs (45 nm) with a positive charge are more easily internalized by NR8383 and Caco-2 cells than the bigger NPs (90 nm) with a positive or negative charge. Furthermore, their results indicated that the NPs with different sizes and charges are taken up through different endocytic pathways, determined by the specific interactions with cell membrane-bound receptors, including clathrin, caveolin, and mannose [73]. Jiang et al. quantified the cellular internalization of different sized-NPs featuring neutral, anionic, and cationic headgroups. They showed that the cellular internalization of neutral, anionic NPs decreased with increasing their size, whereas the opposite is true for cationic NPs. They proposed that this contrasting behavior is ascribed to the surface-dictated shift in uptake pathways [74]. In addition to the interplay between NP size and surface charge, other properties of NPs, such as size and surface chemistry, shell thickness, and size, have also been demonstrated to interact in an interrelated fashion to modulate cellular uptake [75,76]. Recent studies focus on the more complicated multiparameter system where distinct properties ranging from size, shape, stiffness, surface chemistry, and composition are simultaneously investigated to establish the optimal feature of NP for efficient cellular uptake [77,78].
5. Summary and Perspective
Engineering NPs to deliver drug molecules has attracted extensive attention in the past decades, due to their great potential to improve cancer therapy, vaccination, and treatment of genetic disorders. One significant obstacle for developing effective NP-based drug therapies is their limited ability to cross the plasma membranes of cells. Endocytosis is a primary exploited route in NP cellular uptake for drug delivery purposes. There is a growing appreciation that a comprehensive understanding of the endocytosis mechanisms responsible for the cellular uptake will play a central role in nanomedicine. It has been well established that the cellular uptake of the NPs via endocytosis depends strongly on the physicochemical characteristics of NPs such as size, shape, stiffness, which have been investigated individually. In practice, it is the properties of NP that interact in an interrelated fashion to regulate their cellular uptake together. This review summarizes the advances regarding the interplay of NP properties on the cellular uptake during endocytosis mainly from three aspects: (1) Size and shape, (2) size and elasticity, (3) shape and elasticity. For case (1), the efficiency of cellular uptake is determined together by both size and shape. The difference in uptake efficiency is proposed to lie in the receptor-ligand binding and endocytic pathways. For case (2), it indicates that softer NPs can be internalized at a faster or slower rate than the stiffer NPs, depending on the volume constraint, and this wrapping efficiency difference increased with NP size. For case (3), the collaborative effects of shape and elasticity on the uptake efficiency are complicated, some of which are proposed to be a result of competition between the receptor diffusion kinetics and driving force provided by the receptor-ligand complexes. These results highlight the importance of interplay between properties of NPs and shed light on the better NP design in drug delivery.
To develop more efficient intracellular NP-dependent drug-delivery nanosystems, further studies on the properties of NPs based on more complicated multiparameter systems are needed. Accumulating evidence indicates that the cellular uptake of NPs is both property- and cell type-dependent, and there is a difference in the uptake of NPs between cell types, e.g., cancer vs. normal cells, phagocytic vs. nonphagocytic cells, and monocytes vs. macrophages [79,80,81,82,83]. The underlying mechanisms accounting for this cell type-dependent manner are manifold. For example, cancer cells can express different numbers and types of receptors on their surfaces compared with normal cells, thus affecting cellular uptake of NPs by regulating the available binding sites of cargos. In addition, the membrane composition, metabolic activity, cell size, and endocytic pathway can also affect the cellular uptake of NPs [79,80]. There is still much work to be done to understand the cell type-dependent uptake of NPs with various properties. Coupled with innovations in materials science, the results of further studies will continue to contribute to the rational design of clinically useful NPs and the realization of transition from a promising field to the medical application for the NP-based drug-delivery.
Author Contributions
Writing—original draft preparation, review and editing, X.W. L.L. and F.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the NSFC Basic Science Center Program for “Multiscale Problems in Nonlinear Mechanics” (11988102), the Youth Innovation Promotion Association CAS, the National Natural Science Foundation of China (11902327 and 11972041), and the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB22040102).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Jong, W.H.; Borm, P.J. Drug delivery and nanoparticles:applications and hazards. Int. J. Nanomed. 2008, 3, 133–149. [Google Scholar] [CrossRef] [Green Version]
- Ashfaq, U.A.; Riaz, M.; Yasmeen, E.; Yousaf, M.Z. Recent advances in nanoparticle-based targeted drug-delivery systems against cancer and role of tumor microenvironment. Crit. Rev. Ther. Drug Carr. Syst. 2017, 34, 317–353. [Google Scholar] [CrossRef]
- Allen, T.M.; Cullis, P.R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Deliv. Rev. 2013, 65, 36–48. [Google Scholar] [CrossRef]
- Kumari, P.; Ghosh, B.; Biswas, S. Nanocarriers for cancer-targeted drug delivery. J. Drug Target. 2016, 24, 179–191. [Google Scholar] [CrossRef]
- Erdoğar, N.; Akkın, S.; Bilensoy, E. Nanocapsules for drug delivery: An updated review of the last decade. Recent Pat. Drug Deliv. Formul. 2018, 12, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Z.; Langer, R.; Farokhzad, O.C. Nanoparticle delivery of cancer drugs. Annu. Rev. Med. 2012, 63, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Mishra, D.; Hubenak, J.R.; Mathur, A.B. Nanoparticle systems as tools to improve drug delivery and therapeutic efficacy. J. Biomed. Mater. Res. A 2013, 101, 3646–3660. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Ou, W.; Thapa, R.K.; Jiang, L.; Soe, Z.C.; Gautam, M.; Chang, J.H.; Jeong, J.H.; Ku, S.K.; Choi, H.G.; Yong, C.S.; et al. Regulatory T cell-targeted hybrid nanoparticles combined with immuno-checkpoint blockage for cancer immunotherapy. J. Control. Release 2018, 281, 84–96. [Google Scholar] [CrossRef]
- Torchilin, V. Tumor delivery of macromolecular drugs based on the EPR effect. Adv. Drug Deliv. Rev. 2011, 63, 131–135. [Google Scholar] [CrossRef]
- Hui, Y.; Yi, X.; Hou, F.; Wibowo, D.; Zhang, F.; Zhao, D.; Gao, H.; Zhao, C.X. Role of nanoparticle mechanical properties in cancer drug delivery. ACS Nano 2019, 13, 7410–7424. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.M.; Hunter, A.C.; Andresen, T.L. Factors controlling nanoparticle pharmacokinetics: An integrated analysis and perspective. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 481–503. [Google Scholar] [CrossRef] [PubMed]
- Muhamad, N.; Plengsuriyakarn, T.; Na-Bangchang, K. Application of active targeting nanoparticle delivery system for chemotherapeutic drugs and traditional/herbal medicines in cancer therapy: A systematic review. Int. J. Nanomed. 2018, 13, 3921–3935. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.; Tavares, A.J.; Dai, Q.; Ohta, S.; Audet, J.; Dvorak, H.F.; Chan, W.C.W. Analysis of nanoparticle delivery to tumours. Nat. Rev. Mater. 2016, 1, 16014. [Google Scholar] [CrossRef]
- Shen, Z.; Nieh, M.P.; Li, Y. Decorating nanoparticle surface for targeted drug delivery: Opportunities and challenges. Polymers 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Gao, H.; Bao, G. Physical principles of nanoparticle cellular endocytosis. ACS Nano 2015, 9, 8655–8671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Becton, M.; Zhang, L.; Wang, X. Mechanism of coupling nanoparticle stiffness with shape for endocytosis: From rodlike penetration to wormlike wriggling. J. Phys. Chem. B 2020, 124, 11145–11156. [Google Scholar] [CrossRef]
- Dasgupta, S.; Auth, T.; Gompper, G. Wrapping of ellipsoidal nano-particles by fluid membranes. Soft Matter 2013, 9, 5473–5482. [Google Scholar] [CrossRef] [Green Version]
- Bahrami, A.H.; Raatz, M.; Agudo-Canalejo, J.; Michel, R.; Curtis, E.M.; Hall, C.K.; Gradzielski, M.; Lipowsky, R.; Weikl, T.R. Wrapping of nanoparticles by membranes. Adv. Colloid Interface Sci. 2014, 208, 214–224. [Google Scholar] [CrossRef]
- Li, L.; Hu, J.; Shi, X.; Różycki, B.; Song, F. Interplay between cooperativity of intercellular receptor-ligand binding and coalescence of nanoscale lipid clusters in adhering membranes. Soft Matter 2021, 17, 1912–1920. [Google Scholar] [CrossRef]
- Li, L.; Hu, J.; Różycki, B.; Song, F. Intercellular Receptor-Ligand Binding and Thermal Fluctuations Facilitate Receptor Aggregation in Adhering Membranes. Nano Lett. 2020, 20, 722–728. [Google Scholar] [CrossRef]
- Hemmerle, A.; Malaquin, L.; Charitat, T.; Lecuyer, S.; Fragneto, G.; Daillant, J. Controlling interactions in supported bilayers from weak electrostatic repulsion to high osmotic pressure. Proc. Natl. Acad. Sci. USA 2012, 109, 19938–19942. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Hu, J.; Li, L.; Song, F. Binding constant of membrane-anchored receptors and ligands that induce membrane curvatures. Soft Matter 2019, 15, 3507–3514. [Google Scholar] [CrossRef] [Green Version]
- Ziello, J.E.; Huang, Y.; Jovin, I.S. Cellular endocytosis and gene delivery. Mol. Med. 2010, 16, 222–229. [Google Scholar] [CrossRef]
- Oh, N.; Park, J.H. Endocytosis and exocytosis of nanoparticles in mammalian cells. Int. J. Nanomed. 2014, 9 (Suppl. 1), 51–63. [Google Scholar]
- Khan, I.; Steeg, P.S. Endocytosis: A pivotal pathway for regulating metastasis. Br. J. Cancer 2021, 124, 66–75. [Google Scholar] [CrossRef]
- Canton, I.; Battaglia, G. Endocytosis at the nanoscale. Chem. Soc. Rev. 2012, 41, 2718–2739. [Google Scholar] [CrossRef] [PubMed]
- Sousa de Almeida, M.; Susnik, E.; Drasler, B.; Taladriz-Blanco, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Understanding nanoparticle endocytosis to improve targeting strategies in nanomedicine. Chem. Soc. Rev. 2021, 50, 5397–5434. [Google Scholar] [CrossRef] [PubMed]
- Albanese, A.; Tang, P.S.; Chan, W.C. The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annu. Rev. Biomed. Eng. 2012, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Ma, H.; Liu, J.; Huo, S.; Kumar, A.; Wei, T.; Zhang, X.; Jin, S.; Gan, Y.; Wang, P.C.; et al. Size-dependent localization and penetration of ultrasmall gold nanoparticles in cancer cells, multicellular spheroids, and tumors in vivo. ACS Nano 2012, 6, 4483–4493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anselmo, A.C.; Zhang, M.; Kumar, S.; Vogus, D.R.; Menegatti, S.; Helgeson, M.E.; Mitragotri, S. Elasticity of nanoparticles influences their blood circulation, phagocytosis, endocytosis, and targeting. ACS Nano 2015, 9, 3169–3177. [Google Scholar] [CrossRef]
- Yang, K.; Ma, Y.Q. Computer simulation of the translocation of nanoparticles with different shapes across a lipid bilayer. Nat. Nanotechnol. 2010, 5, 579–583. [Google Scholar] [CrossRef]
- Aoyama, Y.; Kanamori, T.; Nakai, T.; Sasaki, T.; Horiuchi, S.; Sando, S.; Niidome, T. Artificial viruses and their application to gene delivery. Size-controlled gene coating with glycocluster nanoparticles. J. Am. Chem. Soc. 2003, 125, 3455–3457. [Google Scholar] [CrossRef]
- Nakai, T.; Kanamori, T.; Sando, S.; Aoyama, Y. Remarkably size-regulated cell invasion by artificial viruses. Saccharide-dependent self-aggregation of glycoviruses and its consequences in glycoviral gene delivery. J. Am. Chem. Soc. 2003, 125, 8465–8475. [Google Scholar] [CrossRef]
- Osaki, F.; Kanamori, T.; Sando, S.; Sera, T.; Aoyama, Y. A quantum dot conjugated sugar ball and its cellular uptake. On the size effects of endocytosis in the subviral region. J. Am. Chem. Soc. 2004, 126, 6520–6521. [Google Scholar] [CrossRef]
- Chithrani, B.D.; Ghazani, A.A.; Chan, W.C. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006, 6, 662–668. [Google Scholar] [CrossRef]
- Win, K.Y.; Feng, S.S. Effects of particle size and surface coating on cellular uptake of polymeric nanoparticles for oral delivery of anticancer drugs. Biomaterials 2005, 26, 2713–2722. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Shi, W.; Freund, L.B. Mechanics of receptor-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2005, 102, 9469–9474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnear, C.; Moore, T.L.; Rodriguez-Lorenzo, L.; Rothen-Rutishauser, B.; Petri-Fink, A. Form Follows Function: Nanoparticle Shape and Its Implications for Nanomedicine. Chem. Rev. 2017, 117, 11476–11521. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Mitragotri, S. Impact of particle elasticity on particle-based drug delivery systems. Adv. Drug Deliv. Rev. 2017, 108, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, R.; Xu, Z.P. Nanoparticle-based nanomedicines to promote cancer immunotherapy: Recent advances and future directions. Small 2019, 15, e1900262. [Google Scholar] [CrossRef] [PubMed]
- Nambara, K.; Niikura, K.; Mitomo, H.; Ninomiya, T.; Takeuchi, C.; Wei, J.; Matsuo, Y.; Ijiro, K. Reverse size dependences of the cellular uptake of triangular and spherical gold nanoparticles. Langmuir 2016, 32, 12559–12567. [Google Scholar] [CrossRef]
- Yue, J.; Feliciano, T.J.; Li, W.; Lee, A.; Odom, T.W. Gold nanoparticle size and shape effects on cellular uptake and intracellular distribution of siRNA nanoconstructs. Bioconjug. Chem. 2017, 28, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Hoshyar, N.; Gray, S.; Han, H.; Bao, G. The effect of nanoparticle size on in vivo pharmacokinetics and cellular interaction. Nanomedicine 2016, 11, 673–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosquera, J.; García, I.; Liz-Marzán, L.M. Cellular uptake of nanoparticles versus small molecules: A matter of size. Acc. Chem. Res. 2018, 51, 2305–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Kröger, M.; Liu, W.K. Shape effect in cellular uptake of PEGylated nanoparticles: Comparison between sphere, rod, cube and disk. Nanoscale 2015, 7, 16631–16646. [Google Scholar] [CrossRef]
- Tree-Udom, T.; Seemork, J.; Shigyou, K.; Hamada, T.; Sangphech, N.; Palaga, T.; Insin, N.; Pan-In, P.; Wanichwecharungruang, S. Shape effect on particle-lipid bilayer membrane association, cellular uptake, and cytotoxicity. ACS Appl. Mater. Interfaces 2015, 7, 23993–24000. [Google Scholar] [CrossRef]
- Chen, L.; Xiao, S.; Zhu, H.; Wang, L.; Liang, H. Shape-dependent internalization kinetics of nanoparticles by membranes. Soft Matter 2016, 12, 2632–2641. [Google Scholar] [CrossRef]
- Li, Z.; Sun, L.; Zhang, Y.; Dove, A.P.; O’Reilly, R.K.; Chen, G. Shape effect of Glyco-Nanoparticles on macrophage cellular uptake and immune response. ACS Macro Lett. 2016, 5, 1059–1064. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Liao, J.; Shao, X.; Li, Q.; Lin, Y. The effect of shape on cellular uptake of gold nanoparticles in the forms of stars, sods, and triangles. Sci. Rep. 2017, 7, 3827. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Auth, T.; Gompper, G. Shape and orientation matter for the cellular uptake of nonspherical particles. Nano Lett. 2014, 14, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yao, C.; Yin, X.; Li, C.; Huang, Y.; Wu, M.; Wang, B.; Guo, X.; Wang, Y.; Wu, M. Size, shape, and protein corona determine cellular uptake and removal mechanisms of gold nanoparticles. Small 2018, 14, e1801451. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, Y.; Yuan, H.; Gao, H.; Zhang, S. Role of nanoparticle geometry in endocytosis: Laying down to stand up. Nano Lett. 2013, 13, 4546–4550. [Google Scholar] [CrossRef] [PubMed]
- Marwah, M.; Perrie, Y.; Badhan, R.K.S.; Lowry, D. Intracellular uptake of EGCG-loaded deformable controlled release liposomes for skin cancer. J. Liposome Res. 2020, 30, 136–149. [Google Scholar] [CrossRef]
- Zheng, Y.; Xing, L.; Chen, L.; Zhou, R.; Wu, J.; Zhu, X.; Li, L.; Xiang, Y.; Wu, R.; Zhang, L.; et al. Tailored elasticity combined with biomimetic surface promotes nanoparticle transcytosis to overcome mucosal epithelial barrier. Biomaterials 2020, 262, 120323. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, C.T.; Li, W.F.; Cheng, J.G.; Tian, X.Q.; Zhao, Y.Z.; Li, X.; Lv, H.F.; Li, X.K. Physical characterization and cellular uptake of propylene glycol liposomes in vitro. Drug Dev. Ind. Pharm. 2012, 38, 365–371. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, L.; Wang, J.; Feng, Q.; Liu, D.; Yin, Q.; Xu, D.; Wei, Y.; Ding, B.; Shi, X.; et al. Tunable rigidity of (polymeric core)-(lipid shell) nanoparticles for regulated cellular uptake. Adv. Mater. 2015, 27, 1402–1407. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Yi, X.; Wibowo, D.; Yang, G.; Middelberg, A.P.J.; Gao, H.; Zhao, C.X. Nanoparticle elasticity regulates phagocytosis and cancer cell uptake. Sci. Adv. 2020, 6, eaaz4316. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Zhou, X.; Mao, Z.; Yu, D.; Gao, C. Uptake of hydrogel particles with different stiffness and its influence on HepG2 cell functions. Soft Matter 2012, 8, 9235–9245. [Google Scholar] [CrossRef]
- Guo, P.; Liu, D.; Subramanyam, K.; Wang, B.; Yang, J.; Huang, J.; Auguste, D.T.; Moses, M.A. Nanoparticle elasticity directs tumor uptake. Nat. Commun. 2018, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Hou, K.; Ren, Q.; Chen, G.; Wei, P.; Zhu, M. Nanoparticle-polymer synergies in nanocomposite hydrogels: From design to application. Macromol. Rapid Commun. 2018, 39, e1800337. [Google Scholar] [CrossRef]
- Elkhoury, K.; Russell, C.S.; Sanchez-Gonzalez, L.; Mostafavi, A.; Williams, T.J.; Kahn, C.; Peppas, N.A.; Arab-Tehrany, E.; Tamayol, A. Soft-nanoparticle functionalization of natural hydrogels for tissue engineering applications. Adv. Healthc. Mater 2019, 8, e1900506. [Google Scholar] [CrossRef]
- Shen, Z.; Ye, H.; Yi, X.; Li, Y. Membrane wrapping efficiency of elastic nanoparticles during endocytosis: Size and shape matter. ACS Nano 2019, 13, 215–228. [Google Scholar] [CrossRef]
- Takechi-Haraya, Y.; Goda, Y.; Sakai-Kato, K. Control of liposomal penetration into three-dimensional multicellular tumor spheroids by modulating liposomal membrane rigidity. Mol. Pharm. 2017, 14, 2158–2165. [Google Scholar] [CrossRef]
- Takechi-Haraya, Y.; Sakai-Kato, K.; Abe, Y.; Kawanishi, T.; Okuda, H.; Goda, Y. Atomic force microscopic analysis of the effect of lipid composition on liposome membrane rigidity. Langmuir 2016, 32, 6074–6082. [Google Scholar] [CrossRef]
- Yi, X.; Gao, H. Kinetics of receptor-mediated endocytosis of elastic nanoparticles. Nanoscale 2017, 9, 454–463. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Burnouf, P.A.; Su, Y.C.; Chen, B.M.; Chuang, K.H.; Lee, C.W.; Wei, P.K.; Cheng, T.L.; Roffler, S.R. Engineering chimeric receptors to investigate the size- and rigidity-dependent tnteraction of PEGylated nanoparticles with cells. ACS Nano 2016, 10, 648–662. [Google Scholar] [CrossRef]
- Hartmann, R.; Weidenbach, M.; Neubauer, M.; Fery, A.; Parak, W.J. Stiffness-dependent in vitro uptake and lysosomal acidification of colloidal particles. Angew. Chem. Int. Ed. Engl. 2015, 54, 1365–1368. [Google Scholar] [CrossRef]
- Sun, H.; Wong, E.H.H.; Yan, Y.; Cui, J.; Dai, Q.; Guo, J.; Qiao, G.G.; Caruso, F. The role of capsule stiffness on cellular processing. Chem. Sci. 2015, 6, 3505–3514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, C.; Liu, B.; Li, B.; Chai, J.; Zhang, L.; Jiao, L.; Li, D.; Yu, Z.; Ren, F.; Shi, X.; et al. Enhanced transport of shape and rigidity-tuned α-Lactalbumin nanotubes across intestinal mucus and cellular barriers. Nano Lett. 2020, 20, 1352–1361. [Google Scholar] [CrossRef]
- Palomba, R.; Palange, A.L.; Rizzuti, I.F.; Ferreira, M.; Cervadoro, A.; Barbato, M.G.; Canale, C.; Decuzzi, P. Modulating phagocytic cell sequestration by tailoring nanoconstruct softness. ACS Nano 2018, 12, 1433–1444. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Zhang, Y.; Chen, T.; Xiao, S.; Liang, H. Morphological and mechanical determinants of cellular uptake of deformable nanoparticles. Nanoscale 2018, 10, 11969–11979. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Ershov, D.; Fytianos, K.; van der Gucht, J.; Alink, G.M.; Rietjens, I.M.; Marcelis, A.T.; Zuilhof, H. Cytotoxicity and cellular uptake of tri-block copolymer nanoparticles with different size and surface characteristics. Part. Fibre Toxicol. 2012, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Huo, S.; Mizuhara, T.; Das, R.; Lee, Y.W.; Hou, S.; Moyano, D.F.; Duncan, B.; Liang, X.J.; Rotello, V.M. The interplay of size and surface functionality on the cellular uptake of sub-10 nm gold nanoparticles. ACS Nano 2015, 9, 9986–9993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walkey, C.D.; Olsen, J.B.; Guo, H.; Emili, A.; Chan, W.C. Nanoparticle size and surface chemistry determine serum protein adsorption and macrophage uptake. J. Am. Chem. Soc. 2012, 134, 2139–2147. [Google Scholar] [CrossRef] [PubMed]
- Panja, P.; Jana, N.R. Lipid-raft-mediated direct cytosolic delivery of polymer-coated soft nanoparticles. J. Phys. Chem. B 2020, 124, 5323–5333. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Wang, S.; Yan, X.; Zhou, H.; Zhan, J.; Liu, S.; Sharma, V.K.; Jiang, G.; Zhu, H.; Yan, B. Regulation of cell uptake and cytotoxicity by nanoparticle core under the controlled shape, size, and surface chemistries. ACS Nano 2020, 14, 289–302. [Google Scholar] [CrossRef]
- Brown, T.D.; Habibi, N.; Wu, D.; Lahann, J.; Mitragotri, S. Effect of nanoparticle composition, size, shape, and stiffness on penetration across the blood-brain barrier. ACS Biomater. Sci. Eng. 2020, 6, 4916–4928. [Google Scholar] [CrossRef]
- Kettler, K.; Veltman, K.; van de Meent, D.; van Wezel, A.; Hendriks, A.J. Cellular uptake of nanoparticles as determined by particle properties, experimental conditions, and cell type. Environ. Toxicol. Chem. 2014, 33, 481–492. [Google Scholar] [CrossRef]
- Xia, Q.; Huang, J.; Feng, Q.; Chen, X.; Liu, X.; Li, X.; Zhang, T.; Xiao, S.; Li, H.; Zhong, Z.; et al. Size-and cell type-dependent cellular uptake, cytotoxicity and in vivo distribution of gold nanoparticles. Int. J. Nanomed. 2019, 14, 6957–6970. [Google Scholar] [CrossRef] [Green Version]
- Blechinger, J.; Bauer, A.T.; Torrano, A.A.; Gorzelanny, C.; Bräuchle, C.; Schneider, S.W. Uptake kinetics and nanotoxicity of silica nanoparticles are cell type dependent. Small 2013, 9, 3970–3980. [Google Scholar] [CrossRef]
- Donahue, N.D.; Acar, H.; Wilhelm, S. Concepts of nanoparticle cellular uptake, intracellular trafficking, and kinetics in nanomedicine. Adv. Drug Deliv. Rev. 2019, 143, 68–96. [Google Scholar] [CrossRef] [PubMed]
- Salatin, S.; Yari Khosroushahi, A. Overviews on the cellular uptake mechanism of polysaccharide colloidal nanoparticles. J. Cell Mol. Med. 2017, 21, 1668–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The interplay of NP size and shape on endocytosis. (a) (i) Confocal fluorescence microscopy images showing the uptake of various gold NPs by RAW 264.7 macrophages after 4 h of incubation. (ii) Dose dependence and (iii) time dependence of the binding of gold NPs with macrophages [49]. (b) (i) Submarine and rocket modes of entry for uptake of NPs with different shapes. (ii,iii) Wrapping fractions for stable states for the discontinuous wrapping transitions W2 (transition from shallow- to the deep-wrapped state) and W3 (transition from the deep-wrapped to the complete-wrapped state), respectively. (iv) and (v) correspond to the energy barriers for both transitions, respectively [51]. (c) (i) TEM images and confocal fluorescence images of U87 cells treated with NPs with different sizes and shapes. (ii) Size- and (iii) shape-dependent cellular uptake of siRNA-conjugated gold NPs [43]. (d) Reverse size-dependent effect of triangular and spherical NPs on cellular uptake [42]. (e) Schematic illustration of the endocytosis mechanism of gold NPs with different sizes and shapes [52]. (f) (i) Schematics of the laying-down-to-standing-up process. (ii,iii) The endocytic pathways for spherocylindrical NPs with an aspect ratio ρ = 2 and ρ = 5.5, respectively [53].
Figure 1.
The interplay of NP size and shape on endocytosis. (a) (i) Confocal fluorescence microscopy images showing the uptake of various gold NPs by RAW 264.7 macrophages after 4 h of incubation. (ii) Dose dependence and (iii) time dependence of the binding of gold NPs with macrophages [49]. (b) (i) Submarine and rocket modes of entry for uptake of NPs with different shapes. (ii,iii) Wrapping fractions for stable states for the discontinuous wrapping transitions W2 (transition from shallow- to the deep-wrapped state) and W3 (transition from the deep-wrapped to the complete-wrapped state), respectively. (iv) and (v) correspond to the energy barriers for both transitions, respectively [51]. (c) (i) TEM images and confocal fluorescence images of U87 cells treated with NPs with different sizes and shapes. (ii) Size- and (iii) shape-dependent cellular uptake of siRNA-conjugated gold NPs [43]. (d) Reverse size-dependent effect of triangular and spherical NPs on cellular uptake [42]. (e) Schematic illustration of the endocytosis mechanism of gold NPs with different sizes and shapes [52]. (f) (i) Schematics of the laying-down-to-standing-up process. (ii,iii) The endocytic pathways for spherocylindrical NPs with an aspect ratio ρ = 2 and ρ = 5.5, respectively [53].

Figure 2.
The interplay of NP size and elasticity on endocytosis. (a) (i) Confocal fluorescent images of HeLa cells incubated with poly(lactic-co-glycolic acid)-lipid (P-L) NPs and poly(lactic-co-glycolic acid)-water-lipid (P-W-L) NPs. (ii) The effect of hypothermia (4 °C) treatment and endocytic inhibitors (CPZ, EIPA, and Genistein) on the cellular uptake of P–L and P–W–L NPs [57]. (iii) Four-, 12-, and 24-h cellular uptake of FA-PEG–modified silica NPs in RAW264.7. All values are means ± SD (n = 3, with * p < 0.05, ** p < 0.01, and # p < 0.001; N.S., not significant) [58]. (b) (i) 24-h cellular uptake of hydrogel NPs by HepG2 cells as a function of particle concentration. Asterisk indicates significant difference at p < 0.05 level [59]. (ii) Elasticity dependence of cellular uptake of nanoliposomes by MDA-MB-231 cells [60]. (c) (i) Snapshots showing the membrane wrapping of soft spherical NPs. (ii) Wrapping ratio evolution for rigid and soft spherical NPs with the same radii. (iii) Wrapping time for rigid and soft spherical NPs of different radii. (iv,v) The NP elastic energy change and membrane energy change during the wrapping process, respectively [63]. (d) (i) Schematics of the receptor-mediated endocytosis of spherical soft and stiff NPs. (ii) The wrapping time as a function of the particle size for soft and stiff NPs. (iii,iv) The variation of elastic energy change and contact radius during endocytosis for different particle-membrane stiffness ratios κp/κm, respectively [66].
Figure 2.
The interplay of NP size and elasticity on endocytosis. (a) (i) Confocal fluorescent images of HeLa cells incubated with poly(lactic-co-glycolic acid)-lipid (P-L) NPs and poly(lactic-co-glycolic acid)-water-lipid (P-W-L) NPs. (ii) The effect of hypothermia (4 °C) treatment and endocytic inhibitors (CPZ, EIPA, and Genistein) on the cellular uptake of P–L and P–W–L NPs [57]. (iii) Four-, 12-, and 24-h cellular uptake of FA-PEG–modified silica NPs in RAW264.7. All values are means ± SD (n = 3, with * p < 0.05, ** p < 0.01, and # p < 0.001; N.S., not significant) [58]. (b) (i) 24-h cellular uptake of hydrogel NPs by HepG2 cells as a function of particle concentration. Asterisk indicates significant difference at p < 0.05 level [59]. (ii) Elasticity dependence of cellular uptake of nanoliposomes by MDA-MB-231 cells [60]. (c) (i) Snapshots showing the membrane wrapping of soft spherical NPs. (ii) Wrapping ratio evolution for rigid and soft spherical NPs with the same radii. (iii) Wrapping time for rigid and soft spherical NPs of different radii. (iv,v) The NP elastic energy change and membrane energy change during the wrapping process, respectively [63]. (d) (i) Schematics of the receptor-mediated endocytosis of spherical soft and stiff NPs. (ii) The wrapping time as a function of the particle size for soft and stiff NPs. (iii,iv) The variation of elastic energy change and contact radius during endocytosis for different particle-membrane stiffness ratios κp/κm, respectively [66].

Figure 3.
The interplay of NP shape and elasticity on endocytosis. (a) Different cellular uptake behavior of peptosomes with different shapes and rigidity as indicated by (i) confocal laser scanning microscope images, (ii) mean fluorescence intensity (MFI), and (iii) apparent permeability coefficients (Papp) [70]. (b) Number of particles internalized by (i) RAW cells and (ii) BMDM cells for discoidal polymeric NPs with different geometries and mechanical properties [71]. (c) (i) Endocytic time for oblate, spheric, and prolate NPs with varied stiffness; (ii–iv) Profile of potential of mean force (PMF) for elastic and rigid NPs with different shapes [72]. (d) (i) Wrapping time as a function of bending constant for oblate, prolate, and spherical NPs. (ii–vii) Comparisons of the (ii,iii) wrapping ratio, (iv,v) contact radius, and (vi,vii) energy barrier for rigid and soft oblate and prolate NPs [63].
Figure 3.
The interplay of NP shape and elasticity on endocytosis. (a) Different cellular uptake behavior of peptosomes with different shapes and rigidity as indicated by (i) confocal laser scanning microscope images, (ii) mean fluorescence intensity (MFI), and (iii) apparent permeability coefficients (Papp) [70]. (b) Number of particles internalized by (i) RAW cells and (ii) BMDM cells for discoidal polymeric NPs with different geometries and mechanical properties [71]. (c) (i) Endocytic time for oblate, spheric, and prolate NPs with varied stiffness; (ii–iv) Profile of potential of mean force (PMF) for elastic and rigid NPs with different shapes [72]. (d) (i) Wrapping time as a function of bending constant for oblate, prolate, and spherical NPs. (ii–vii) Comparisons of the (ii,iii) wrapping ratio, (iv,v) contact radius, and (vi,vii) energy barrier for rigid and soft oblate and prolate NPs [63].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, X.; Li, L.; Li, L.; Song, F.; Song, F. Interplay of Nanoparticle Properties during Endocytosis. Crystals 2021, 11, 728. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11070728
AMA Style
Wang X, Li L, Li L, Song F, Song F. Interplay of Nanoparticle Properties during Endocytosis. Crystals. 2021; 11(7):728. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11070728
Chicago/Turabian StyleWang, Xiaohuan, Long Li, Long Li, Fan Song, and Fan Song. 2021. "Interplay of Nanoparticle Properties during Endocytosis" Crystals 11, no. 7: 728. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst11070728
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.