
Secondary Structure and X-ray Crystallographic Analysis of the Glideosome-Associated Connector (GAC) from Toxoplasma gondii

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
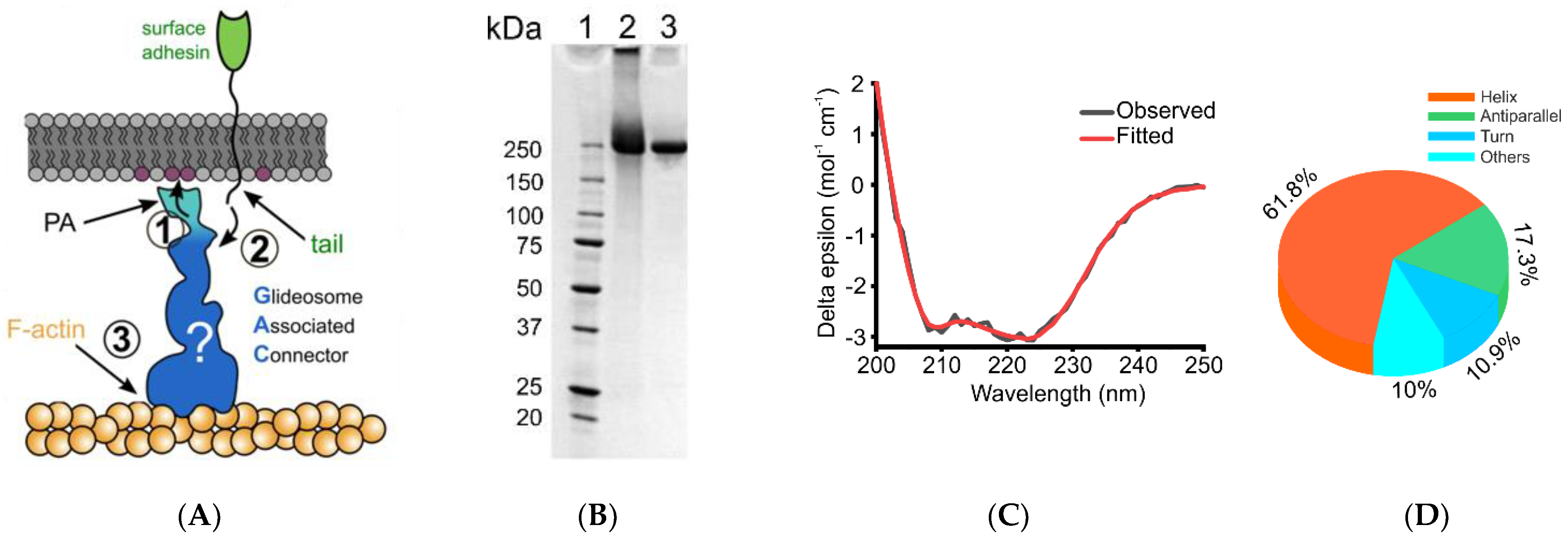
2.1. Protein Expression and Purification
2.2. TgGAC Crystallization
2.3. Circular Dichroism (CD) Spectroscopy
2.4. X-ray Data Collection and Processing
3. Results and Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meissner, M. The asexual cycle of apicomplexan parasites: New findings that raise new questions. Curr. Opin. Microbiol. 2013, 16, 421–423. [Google Scholar] [CrossRef] [Green Version]
- Heintzelman, M.B. Gliding motility in apicomplexan parasites. Semin. Cell Dev. Biol. 2015, 46, 135–142. [Google Scholar] [CrossRef]
- Frenal, K.; Dubremetz, J.F.; Lebrun, M.; Soldati-Favre, D. Gliding motility powers invasion and egress in Apicomplexa. Nat. Rev. Microbiol. 2017, 15, 645–660. [Google Scholar] [CrossRef] [Green Version]
- Boucher, L.E.; Bosch, J. The apicomplexan glideosome and adhesins—Structures and function. J. Struct. Biol. 2015, 190, 93–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowman, A.F.; Tonkin, C.J.; Tham, W.H.; Duraisingh, M.T. The Molecular Basis of Erythrocyte Invasion by Malaria Parasites. Cell Host Microbe 2017, 22, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, V.B.; Tomley, F.M. Microneme proteins in apicomplexans. Subcell. Biochem. 2008, 47, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.J.; Ramaswamy, R.; Kelsen, A.; Hamelin, D.J.; Warshaw, D.M.; Bosch, J.; Burke, J.E.; Ward, G.E.; Boulanger, M.J. Structural and mechanistic insights into the function of the unconventional class XIV myosin MyoA from Toxoplasma gondii. Proc. Natl. Acad. Sci. USA 2018, 115, E10548–E10555. [Google Scholar] [CrossRef] [Green Version]
- Jacot, D.; Tosetti, N.; Pires, I.; Stock, J.; Graindorge, A.; Hung, Y.F.; Han, H.; Tewari, R.; Kursula, I.; Soldati-Favre, D. An Apicomplexan Actin-Binding Protein Serves as a Connector and Lipid Sensor to Coordinate Motility and Invasion. Cell Host Microbe 2016, 20, 731–743. [Google Scholar] [CrossRef] [Green Version]
- Darvill, N.; Dubois, D.J.; Rouse, S.L.; Hammoudi, P.M.; Blake, T.; Benjamin, S.; Liu, B.; Soldati-Favre, D.; Matthews, S. Structural Basis of Phosphatidic Acid Sensing by APH in Apicomplexan Parasites. Structure 2018, 26, 1059–1071.e1056. [Google Scholar] [CrossRef] [Green Version]
- Bullen, H.E.; Jia, Y.; Yamaryo-Botte, Y.; Bisio, H.; Zhang, O.; Jemelin, N.K.; Marq, J.B.; Carruthers, V.; Botte, C.Y.; Soldati-Favre, D. Phosphatidic Acid-Mediated Signaling Regulates Microneme Secretion in Toxoplasma. Cell Host Microbe 2016, 19, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Whitelaw, J.A.; Latorre-Barragan, F.; Gras, S.; Pall, G.S.; Leung, J.M.; Heaslip, A.; Egarter, S.; Andenmatten, N.; Nelson, S.R.; Warshaw, D.M.; et al. Surface attachment, promoted by the actomyosin system of Toxoplasma gondii is important for efficient gliding motility and invasion. BMC Biol. 2017, 15, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Beilsten-Edmands, J.; Winter, G.; Gildea, R.; Parkhurst, J.; Waterman, D.; Evans, G. Scaling diffraction data in the DIALS software package: Algorithms and new approaches for multi-crystal scaling. Acta Crystallogr. D Struct. Biol. 2020, 76, 385–399. [Google Scholar] [CrossRef] [Green Version]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Winter, G. xia2: An expert system for macromolecular crystallography data reduction. J. Appl. Crystallogr. 2010, 43, 186–190. [Google Scholar] [CrossRef]
- Winter, G.; Waterman, D.G.; Parkhurst, J.M.; Brewster, A.S.; Gildea, R.J.; Gerstel, M.; Fuentes-Montero, L.; Vollmar, M.; Michels-Clark, T.; Young, I.D.; et al. DIALS: Implementation and evaluation of a new integration package. Acta Crystallogr. D Struct. Biol. 2018, 74, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Lobley, C.M.; Prince, S.M. Decision making in xia2. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 1260–1273. [Google Scholar] [CrossRef] [Green Version]
- Vonrhein, C.; Flensburg, C.; Keller, P.; Sharff, A.; Smart, O.; Paciorek, W.; Womack, T.; Bricogne, G. Data processing and analysis with the autoPROC toolbox. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Vonrhein, C.; Blanc, E.; Roversi, P.; Bricogne, G. Automated structure solution with autoSHARP. Methods Mol. Biol. 2007, 364, 215–230. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, B.W. Solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–497. [Google Scholar] [CrossRef]
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Improved protein structure prediction using potentials from deep learning. Nature 2020, 577, 706–710. [Google Scholar] [CrossRef]
- Källberg, M.; Wang, H.; Wang, S.; Peng, J.; Wang, Z.; Lu, H.; Xu, J. Template-based protein structure modeling using the RaptorX web server. Nat. Protoc. 2012, 7, 1511–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Crystal | Native | SeMet |
|---|---|---|
| Space group | P212121 | P212121 |
| Cell dimensions (Å) | a = 120.63, b = 123.96, c = 221.86 | a = 119.08, b = 123.60, c = 221.51 |
| Angles (°) | α = 90.00, β = 90.00, γ = 90.00 | α = 90.00, β = 90.00, γ = 90.00 |
| Resolution (Ǻ) | 82.66–2.92 (2.97–2.92) * | 110.75–2.67 (2.67–2.72) * |
| Wavelength (Ǻ) | 0.97950 | 0.97950 |
| Total reflections | 2,899,820 (140,928) | 904,519 (44214) |
| Unique observations | 72,969 (3564) | 92,922 (4581) |
| Completeness (%) | 100 (99–100) | 100 (99.7–100) |
| Multiplicity | 39.7 (39.5) | 9.7 (9.7) |
| Rpim† | 0.045 (1.486) | 0.039 (0.861) |
| <I>/σI | 13.1 (88–0.5) | 14 (47.2–0.8) |
| Molecules per asymmetric unit ‡ | 1 | 1 |
| Solvent content (%) | 56.8 | 56.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, A.; Zhang, X.; Vadas, O.; Stylianou, F.A.; Dos Santos Pacheco, N.; Rouse, S.L.; Morgan, M.L.; Soldati-Favre, D.; Matthews, S. Secondary Structure and X-ray Crystallographic Analysis of the Glideosome-Associated Connector (GAC) from Toxoplasma gondii. Crystals 2022, 12, 110. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12010110
Kumar A, Zhang X, Vadas O, Stylianou FA, Dos Santos Pacheco N, Rouse SL, Morgan ML, Soldati-Favre D, Matthews S. Secondary Structure and X-ray Crystallographic Analysis of the Glideosome-Associated Connector (GAC) from Toxoplasma gondii. Crystals. 2022; 12(1):110. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12010110
Chicago/Turabian StyleKumar, Amit, Xu Zhang, Oscar Vadas, Fisentzos A. Stylianou, Nicolas Dos Santos Pacheco, Sarah L. Rouse, Marc L. Morgan, Dominique Soldati-Favre, and Steve Matthews. 2022. "Secondary Structure and X-ray Crystallographic Analysis of the Glideosome-Associated Connector (GAC) from Toxoplasma gondii" Crystals 12, no. 1: 110. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12010110