
Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Single Stranded DNA Binding Protein (LjSSB) from Psychrophilic Lacinutrix jangbogonensis PAMC 27137

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
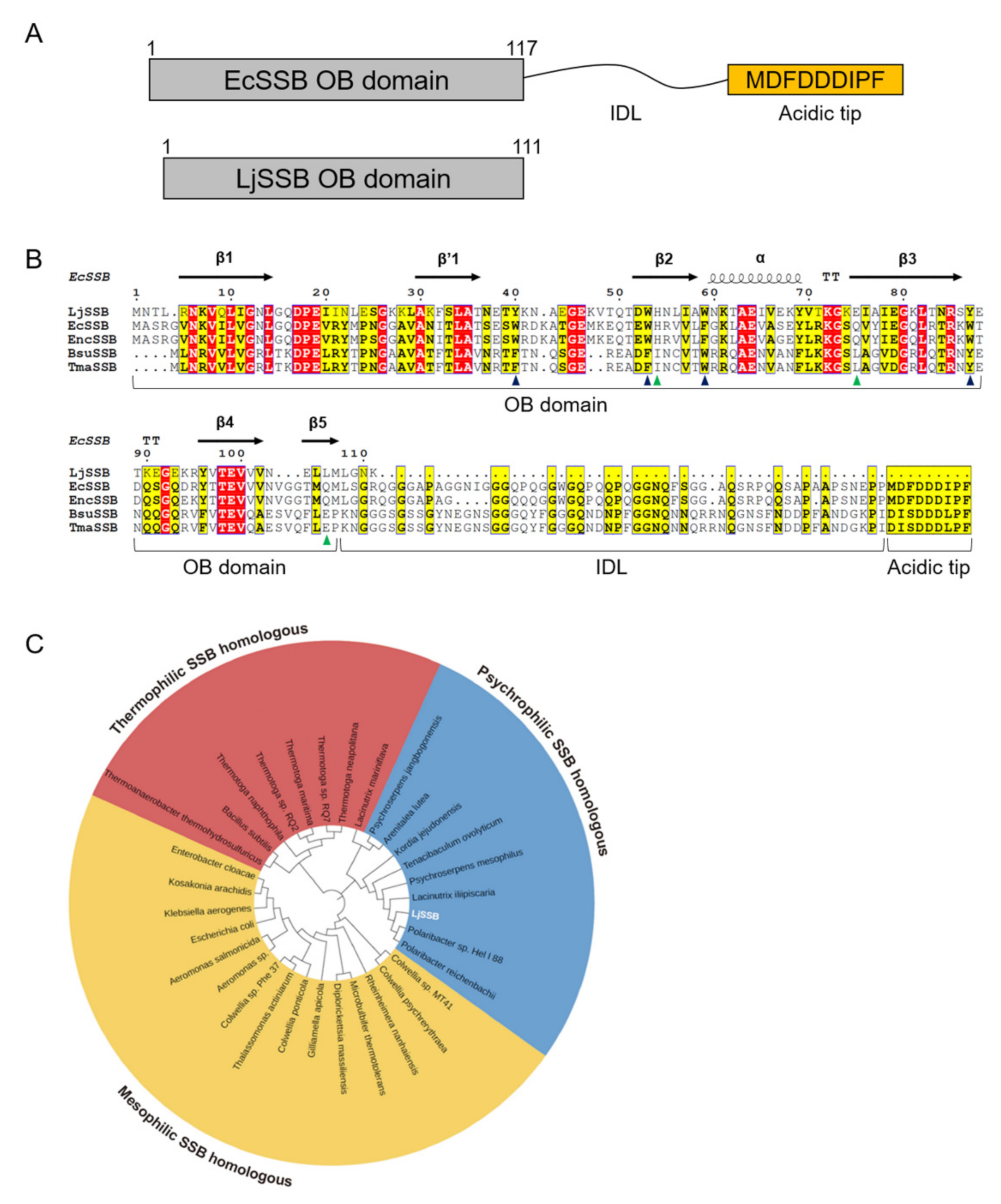
2.1. Sequence Analysis for LjSSB
2.2. Cloning, Expression, and Purification of the LjSSB from L. jangbogonensis
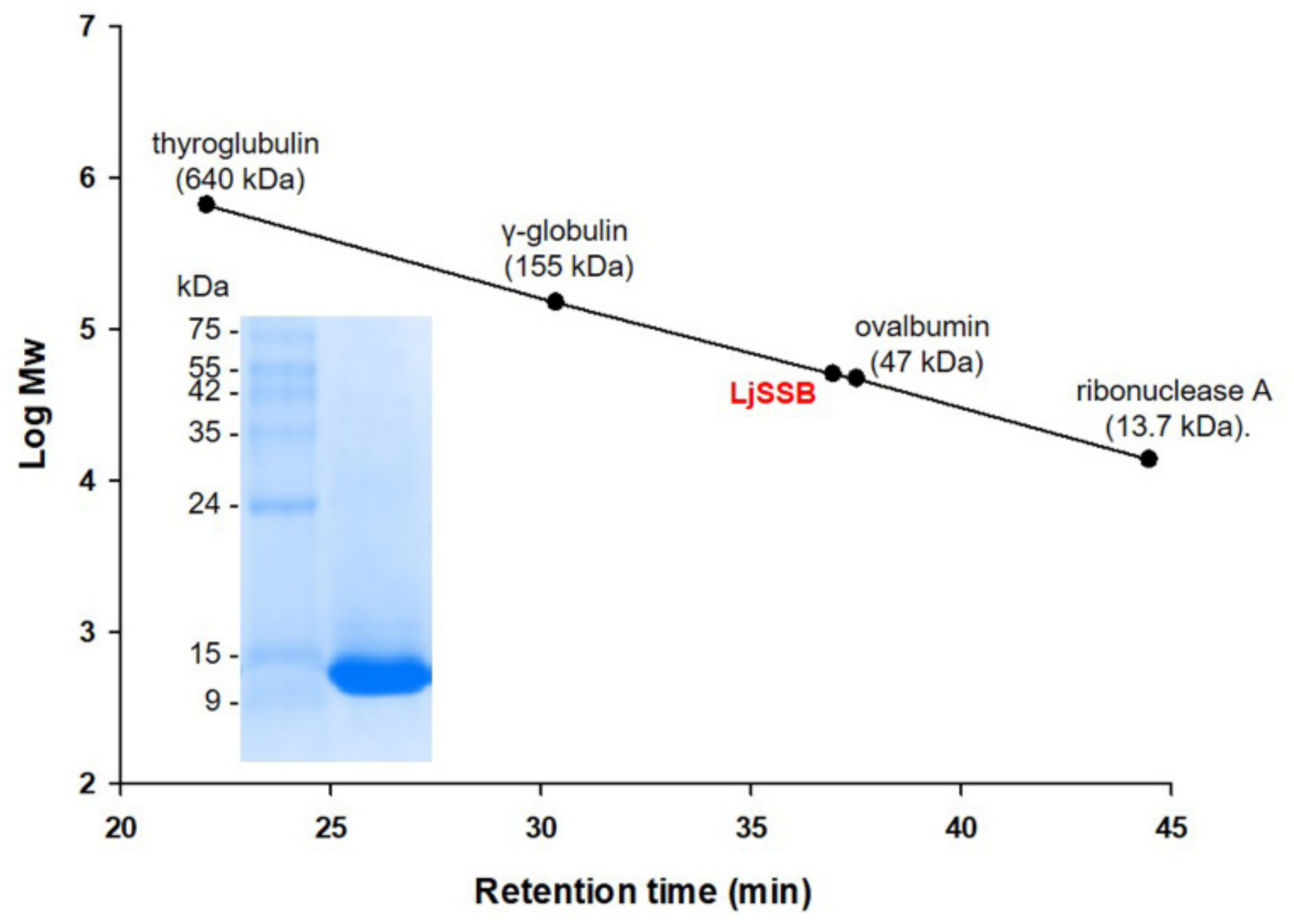
2.3. Estimation of Oligomerization State
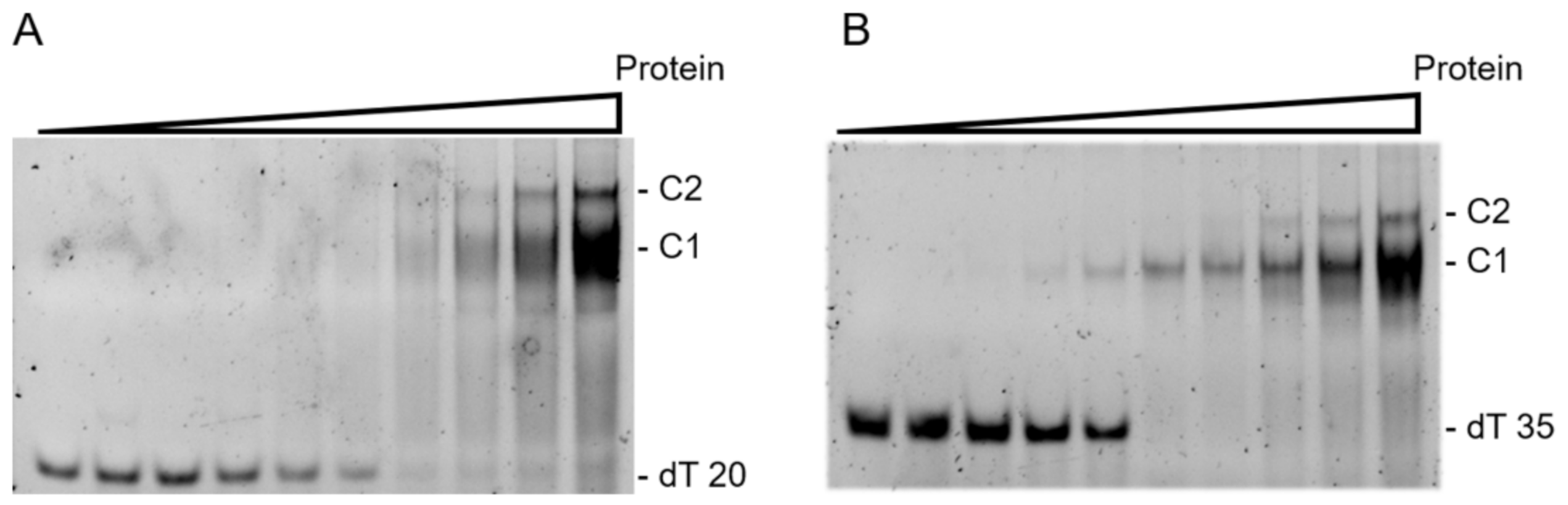
2.4. Enzymatic Analysis Using Gel Electrophoresis Mobility Shift Assay (EMSA)
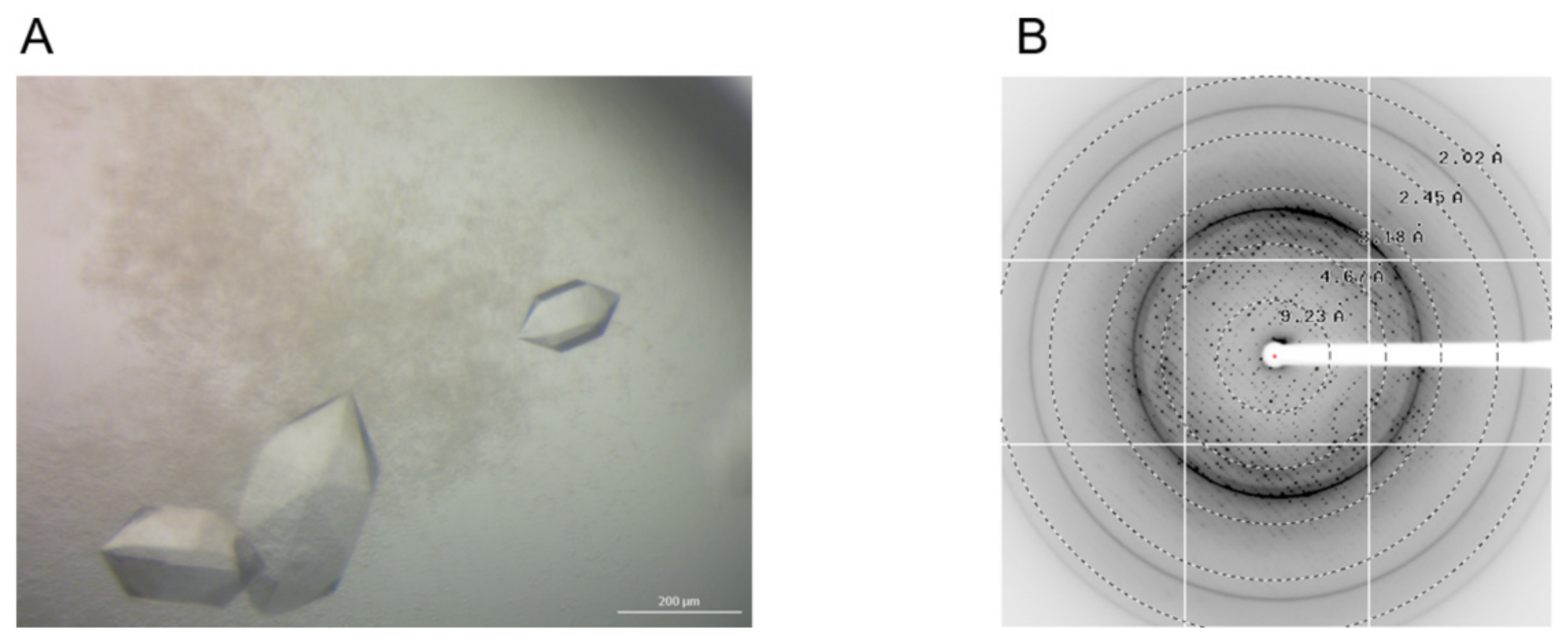
2.5. Crystallization, Data Collection, and Refinement for LjSSB
3. Results and Discussion
3.1. Sequence Analysis for LjSSB

3.2. Oligomerization Status for LjSSB
3.3. Binding Properties for LjSSB with ssDNA
3.4. Crystallization and X-ray Structure Analysis of LjSSB
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marceau, A.H. Functions of single-strand DNA-binding proteins in DNA replication, recombination, and repair. In Methods in Molecular Biology; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2012; Volume 922, pp. 1–21. [Google Scholar]
- Lohman, T.M.; Ferrari, M.E. Escherichia coli single-stranded DNA-binding protein: Multiple DNA-binding modes and cooperativities. Annu. Rev. Biochem. 1994, 63, 527–570. [Google Scholar] [CrossRef]
- Mishra, G.; Levy, Y. Molecular determinants of the interactions between proteins and ssDNA. Proc. Natl. Acad. Sci. USA 2015, 112, 5033–5038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestryakov, P.E.; Lavrik, O.I. Mechanisms of single-stranded DNA-binding protein functioning in cellular DNA metabolism. Biochemistry (Moscow) 2008, 73, 1388–1404. [Google Scholar] [CrossRef] [PubMed]
- Theobald, D.L.; Mitton-Fry, R.M.; Wuttke, D.S. Nucleic acid recognition by OB-fold proteins. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 115–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, R.L.; Zou, L. Oligonucleotide/oligosaccharide-binding fold proteins: A growing family of genome guardians. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antony, E.; Lohman, T.M. Dynamics of E. coli single stranded DNA binding (SSB) protein-DNA complexes. Semin. Cell Dev. Biol. 2019, 86, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinn, M.K.; Kozlov, A.G.; Nguyen, B.; Bujalowski, W.M.; Lohman, T.M. Are the intrinsically disordered linkers involved in SSB binding to accessory proteins? Nucleic Acids Res. 2019, 47, 8581–8594. [Google Scholar] [CrossRef]
- Costes, A.; Lecointe, F.; McGovern, S.; Quevillon-Cheruel, S.; Polard, P. The C-terminal domain of the bacterial SSB protein acts as a DNA maintenance hub at active chromosome replication forks. PLoS Genet. 2010, 6, e1001238. [Google Scholar] [CrossRef] [Green Version]
- Bianco, P.R.; Pottinger, S.; Tan, H.Y.; Nguyenduc, T.; Rex, K.; Varshney, U. The IDL of E. coli SSB links ssDNA and protein binding by mediating protein–protein interactions. Protein Sci. 2017, 26, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Huang, J. Replication protein A and more: Single-stranded DNA-binding proteins in eukaryotic cells. Acta Biochim. Biophys. Sin. 2016, 48, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Raghunathan, S.; Kozlov, A.G.; Lohman, T.M.; Waksman, G. Structure of the DNA binding domain of E. coli SSB bound to ssDNA. Nat. Struct. Biol. 2000, 7, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Pavletich, N.P. Structure and conformational change of a replication protein A heterotrimer bound to ssDNA. Genes Dev. 2012, 26, 2337–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, N.W.; Bolderson, E.; Cubeddu, L.; O’Byrne, K.J.; Richard, D.J. Human single-stranded DNA binding proteins are essential for maintaining genomic stability. BMC Mol. Biol. 2013, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapley, R. Enhancing PCR amplification and sequencing using DNA-binding proteins. Mol. Biotechnol. 1994, 2, 295–298. [Google Scholar] [CrossRef]
- Dąbrowski, S.; Kur, J. Cloning, overexpression, and purification of the recombinant his-tagged SSB protein of Escherichia coli and use in polymerase chain reaction amplification. Protein Expr. Purif. 1999, 16, 96–102. [Google Scholar] [CrossRef]
- Perales, C.; Cava, F.; Meijer, W.J.J.; Berenguer, J. Enhancement of DNA, cDNA synthesis and fidelity at high temperatures by a dimeric single-stranded DNA-binding protein. Nucleic Acids Res. 2003, 31, 6473–6480. [Google Scholar] [CrossRef] [Green Version]
- Ducani, C.; Bernardinelli, G.; Högberg, B. Rolling circle replication requires single-stranded DNA binding protein to avoid termination and production of double-stranded DNA. Nucleic Acids Res. 2014, 42, 10596–10604. [Google Scholar] [CrossRef] [Green Version]
- Cavicchioli, R.; Charlton, T.; Ertan, H.; Omar, S.M.; Siddiqui, K.S.; Williams, T.J. Biotechnological uses of enzymes from psychrophiles. Microb. Biotechnol. 2011, 4, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Shin, S.C.; Baek, K.; Hwang, C.Y.; Hong, S.G.; Chun, J.; Lee, H.K. Draft genome sequence of the psychrophilic bacterium Lacinutrix jangbogonensis PAMC 27137T. Mar. Genom. 2015, 23, 31–32. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using clustalW and clustalX. In Current Protocol in Bioinformatics; Baxevanis, A.D., Pearson, W.R., Stein, L.D., Storomo, G.D., Yates, J.R., III, Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2002; pp. 2–3. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shereda, R.D.; Kozlov, A.G.; Lohman, T.M.; Cox, M.M.; Keck, J.L. SSB as an organizer/mobilizer of genome maintenance complexes. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 289–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Shamoo, Y.; Friedman, A.M.; Parsons, M.R.; Konigsberg, W.H.; Steitz, T.A. Crystal structure of a replication fork single-stranded DNA binding protein (T4 gp32) complexed to DNA. Nature 1995, 376, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.A.; Eggington, J.M.; Killoran, M.P.; Misic, A.M.; Cox, M.M.; Keck, J.L. Crystal structure of the Deinococcus radiodurans single-stranded DNA-binding protein suggests a mechanism for coping with DNA damage. Proc. Natl. Acad. Sci. USA 2004, 101, 8575. [Google Scholar] [CrossRef] [Green Version]
- Wold, M.S. Replication protein A: A heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu. Rev. Biochem. 1997, 66, 61–92. [Google Scholar] [CrossRef]
- Norais, C.A.; Chitteni-Pattu, S.; Wood, E.A.; Inman, R.B.; Cox, M.M. DdrB protein, an alternative Deinococcus radiodurans SSB induced by ionizing radiation. J. Biol. Chem. 2009, 284, 21402–21411. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.H.; Lee, Y.L.; Huang, C.Y. Characterization of a single-stranded DNA binding protein from Salmonella enterica serovar Typhimurium LT2. Protein J. 2011, 30, 102–108. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Chen, I.C.; Huang, C.-Y. Characterization of an SSB–dT25 complex: Structural insights into the S-shaped ssDNA binding conformation. RSC Adv. 2019, 9, 40388–40396. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.G.; Cox, M.M.; Lohman, T.M. Regulation of single-stranded DNA binding by the C termini of Escherichia coli single-stranded DNA-binding (SSB) protein. J. Biol. Chem. 2010, 285, 17246–17252. [Google Scholar] [CrossRef] [Green Version]
- Su, X.-C.; Wang, Y.; Yagi, H.; Shishmarev, D.; Mason, C.E.; Smith, P.J.; Vandevenne, M.; Dixon, N.E.; Otting, G. Bound or free: Interaction of the C-terminal domain of Escherichia coli single-stranded DNA-binding protein (SSB) with the tetrameric core of SSB. Biochemistry 2014, 53, 1925–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Vagin, A.A.; Steiner, R.A.; Lebedev, A.A.; Potterton, L.; McNicholas, S.; Long, F.; Murshudov, G.N. REFMAC5 dictionary: Organization of prior chemical knowledge and guidelines for its use. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2184–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix. refine. Acta Cryst. D Biol. Cryst. 2012, 68, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Cryst. D Biol. Cryst. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | |
|---|---|
| Wavelength (Å) | 0.9796 |
| X-ray source | PAL 5C |
| Rotation range per image (°) | 1 |
| Exposure Time (s) | 0.5 |
| Space group | C 2 2 2 |
| Unit-cell parameters (Å, °) | a = 106.58, b = 231.14, c = 66.14 |
| Resolution range (Å) a | 50.00–2.60 (2.70–2.60) |
| No. of observed reflections | 71,319 |
| No. of unique reflections a | 37,560 (3753) |
| Completeness (%) a | 99.4 (98.7) |
| Redundancy a | 7.1 (7.6) |
| Rsym a,b | 0.138 (0.771) |
| Rmeas a,b | 0.144 (0.800) |
| I/σ a | 14.23 (4.67) |
| CC(1/2) c (%) | 99.9 (94.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, W.; Son, J.; Park, A.; Jin, H.; Shin, S.C.; Lee, J.H.; Kim, T.D.; Kim, H.-W. Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Single Stranded DNA Binding Protein (LjSSB) from Psychrophilic Lacinutrix jangbogonensis PAMC 27137. Crystals 2022, 12, 538. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12040538
Choi W, Son J, Park A, Jin H, Shin SC, Lee JH, Kim TD, Kim H-W. Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Single Stranded DNA Binding Protein (LjSSB) from Psychrophilic Lacinutrix jangbogonensis PAMC 27137. Crystals. 2022; 12(4):538. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12040538
Chicago/Turabian StyleChoi, Woong, Jonghyeon Son, Aekyung Park, Hongshi Jin, Seung Chul Shin, Jun Hyuck Lee, T. Doohun Kim, and Han-Woo Kim. 2022. "Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Single Stranded DNA Binding Protein (LjSSB) from Psychrophilic Lacinutrix jangbogonensis PAMC 27137" Crystals 12, no. 4: 538. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12040538