
Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Novel Esterase (ScEst) from Staphylococcus chromogenes

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
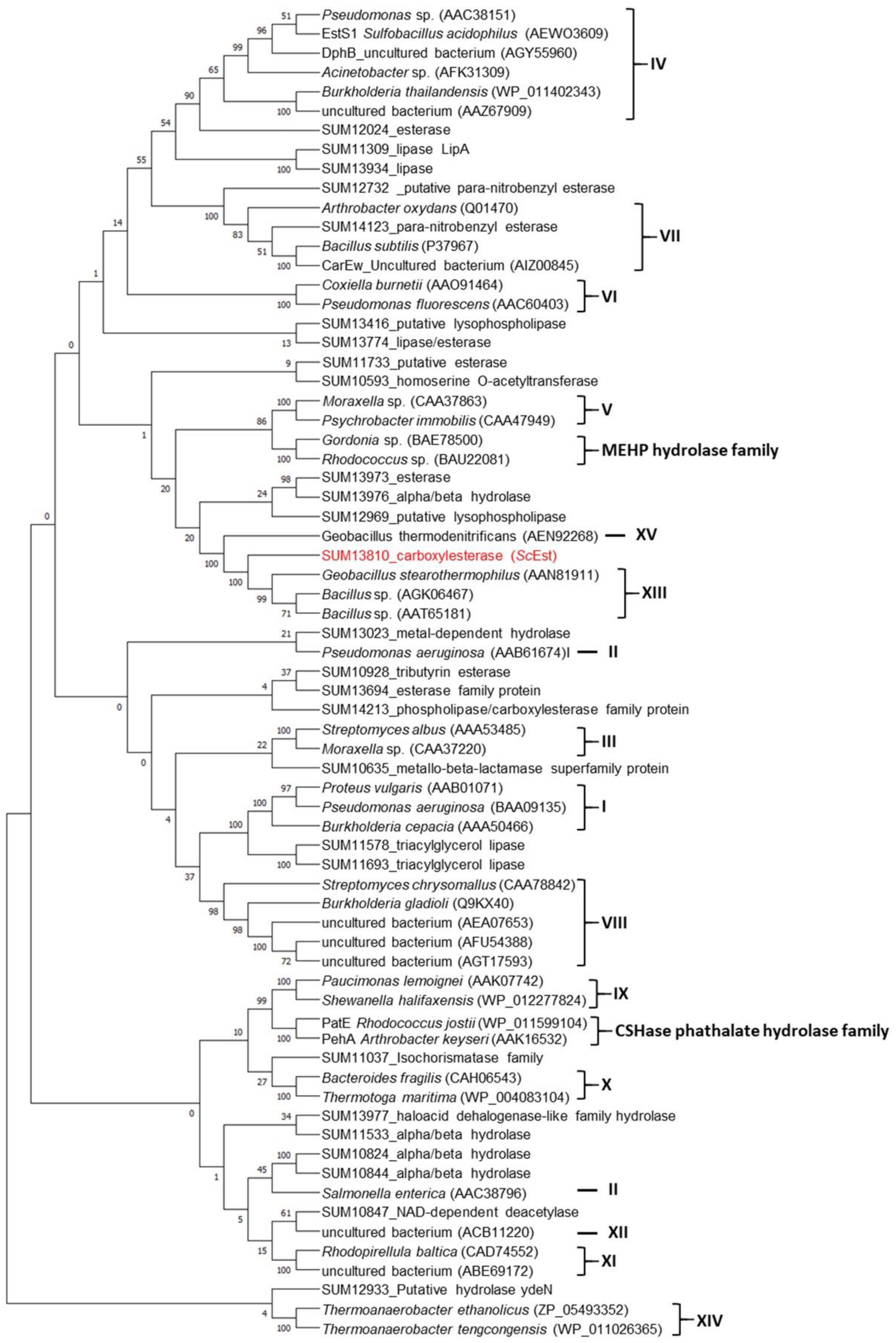
2.1. Phylogenetic Analysis
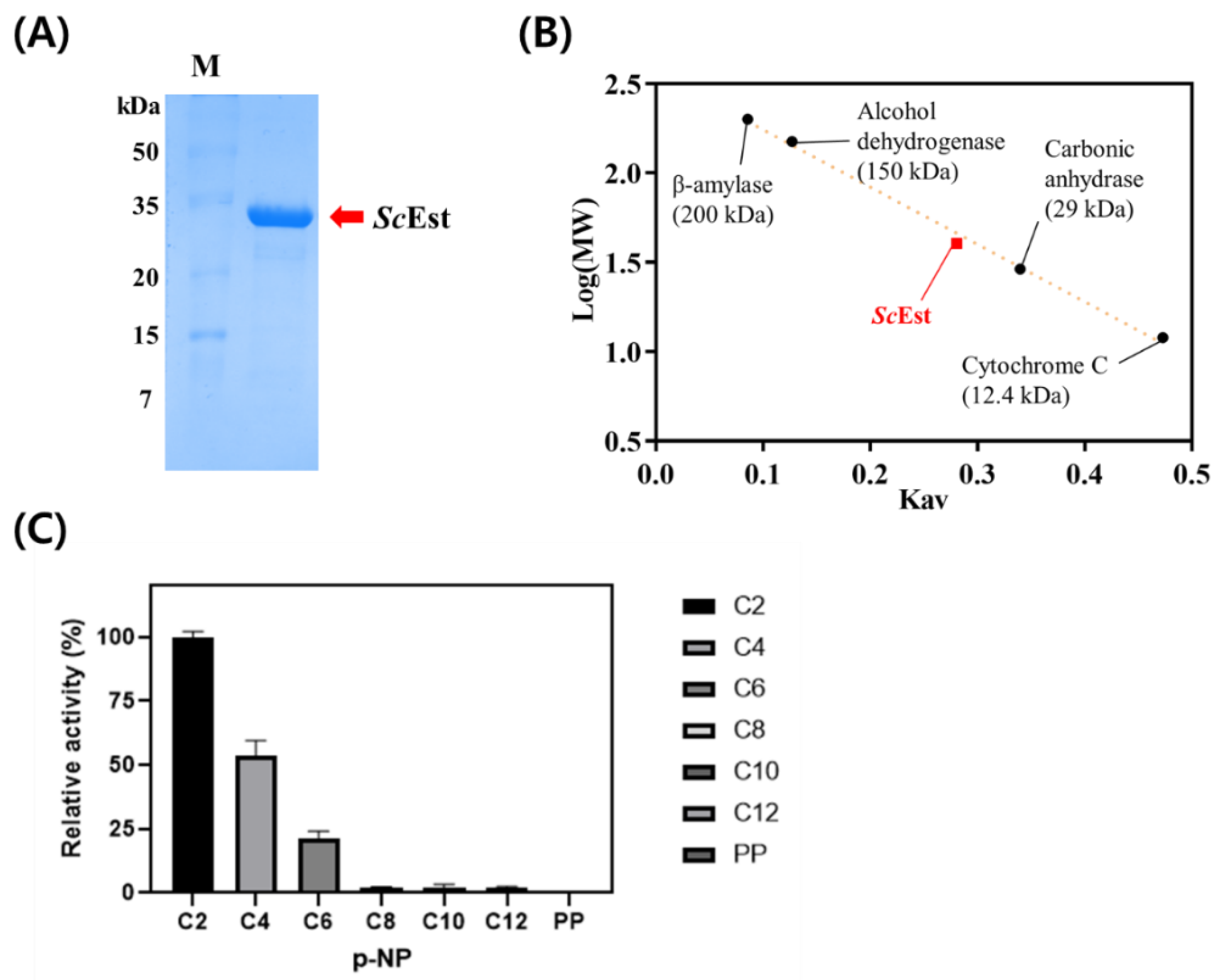
2.2. Gene Cloning, Expression, and Purification of Recombinant ScEst Protein
2.3. Enzymatic Analysis
2.4. Crystallization, Data Collection, and Structural Analysis
3. Results and Discussion
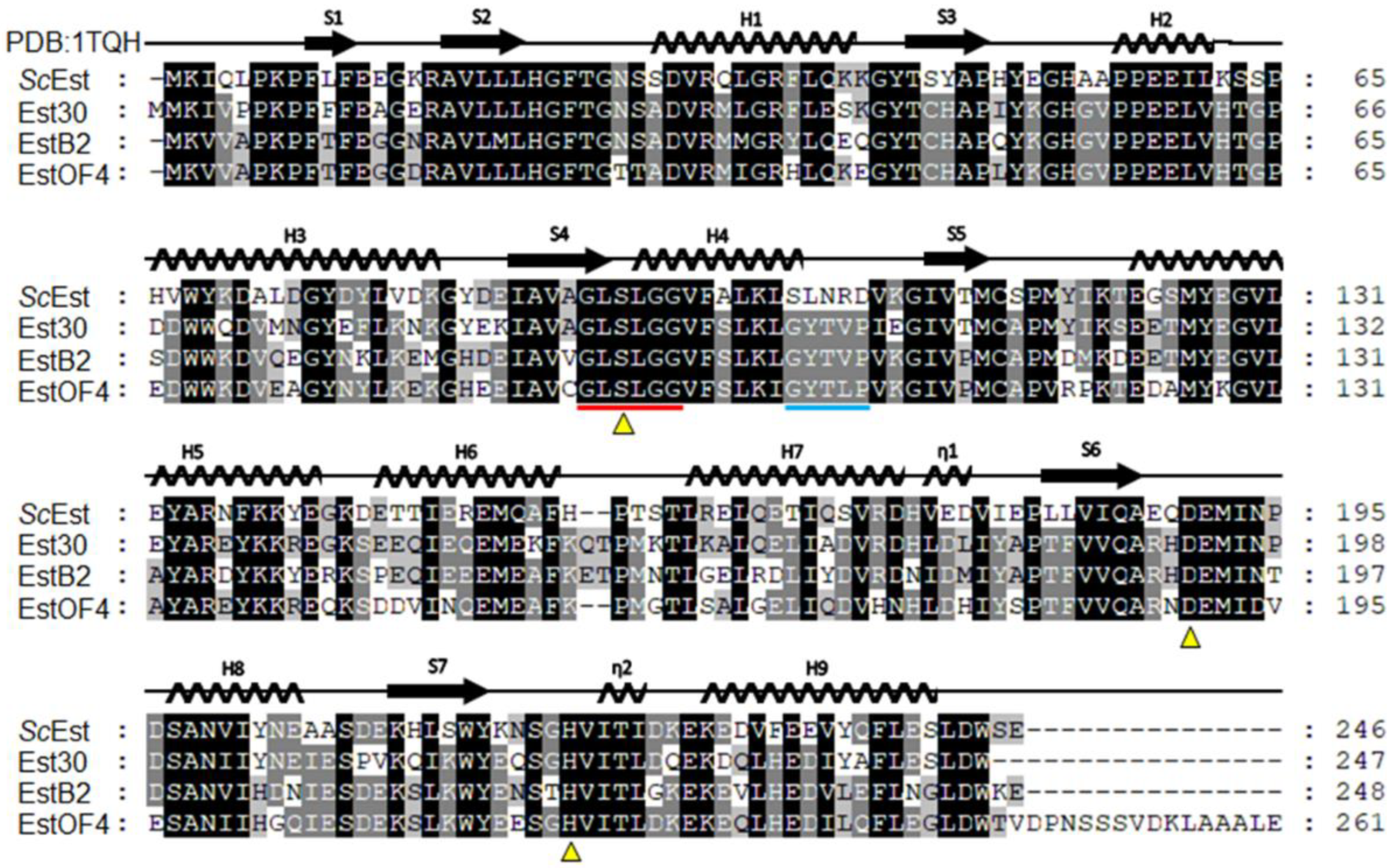
3.1. Lipolytic Enzymes of S. chromogenes NCTC10530 and Classification of ScEst
3.2. Biochemical Characterization of ScEst
3.3. X-ray Crystallographic Study of ScEst
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikaido, H. Multidrug resistance in bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra-Burriel, M.; Keys, M.; Campillo-Artero, C.; Agodi, A.; Barchitta, M.; Gikas, A.; Palos, C.; López-Casasnovas, G. Impact of multi-drug resistant bacteria on economic and clinical outcomes of healthcare-associated infections in adults: Systematic review and meta-analysis. PLoS ONE 2020, 15, e0227139. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.G.; De La Cruz Claure, M.L.; Benito-Arenas, R.; Revuelta, J.; Santana, A.G.; Bastida, A. Overcoming aminoglycoside enzymatic resistance: Design of novel antibiotics and inhibitors. Molecules 2018, 23, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Medicin. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubeh, R.; Breijyeh, B.; Karaman, Z. Antibacterial prodrugs to overcome bacterial resistance. Molecules 2020, 25, 1543. [Google Scholar] [CrossRef] [Green Version]
- Sakagami, K.; Atsumi, K.; Tamura, A.; Yoshida, T.; Nishihata, K.; Fukatsu, S.J. Antibiot. Synthesis and oral activity of ME 1207, a new orally active cephalosporin. J. Antibiot. 1990, 8, 8–11. [Google Scholar] [CrossRef] [Green Version]
- Maag, H. Prodrugs of Carboxylic Acids, Prodrugs; Springer: New York, NY, USA, 2008; pp. 703–729. [Google Scholar] [CrossRef]
- Hamada, Y. Recent progress in prodrug design strategies based on generally applicable modifications. Bioorg. Med. Chem. Lett. 2017, 27, 1627–1632. [Google Scholar] [CrossRef]
- Elayyan, S.; Karaman, D.; Mecca, G.; Scrano, L.; Bufo, S.A.; Karaman, R. Antibacterial predrugs-from 1899 till 2015. World J. Pharm. Pharm. Sci. 2015, 4, 1504–1529. [Google Scholar] [CrossRef]
- Larsen, E.M.; Johnson, R.J. Microbial esterases and ester prodrugs: An unlikely marriage for combating antibiotic resistance. Drug Dev. Res. 2019, 80, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Satoh, T.; Hosokawa, M. Structure, function and regulation of carboxylesterases. Chem. Biol. Interact. 2006, 162, 195–211. [Google Scholar] [CrossRef]
- Mikati, M.O.; Miller, J.J.; Osbourn, D.M.; Barekatain, Y.; Ghebremichael, N.; Shah, I.T.; Burnham, C.D.; Heidel, K.M.; Yan, V.C.; Muller, F.L.; et al. Antimicrobial prodrug activation by the Staphylococcal glyoxalase GloB. ACS Infect. Dis. 2020, 6, 3064–3075. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Le, L.T.H.L.; Yoo, W.; Lee, C.W.; Kim, K.K.; Lee, J.H.; Kim, T.D. Characterization, immobilization, and mutagenesis of a novel cold-active acetylesterase (EaAcE) from Exiguobacterium antarcticum B7. Int. J. Biol. Macromol. 2019, 136, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.; Ryu, B.H.; An, D.R.; Nguyen, D.D.; Yoo, W.; Kim, T.; Ngo, T.D.; Kim, H.S.; Kim, K.K.; Kim, T.D. Structural and biochemical characterization of an octameric carbohydrate acetylesterase from Sinorhizobium meliloti. FEBS Lett. 2016, 590, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Kwon, S.; Park, S.H.; Kim, B.Y.; Yoo, W.; Ryu, B.H.; Kim, H.W.; Shin, S.C.; Kim, S.; Park, H.; et al. Crystal structure and functional characterization of an esterase (EaEST) from exiguobacterium antarcticum. PLoS ONE 2017, 12, e0169540. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Dutta, A.; Pal Chowdhury, P.; Chakraborty, J.; Dutta, T.K. Characterization of a novel family VIII esterase EstM2 from soil metagenome capable of hydrolyzing estrogenic phthalates. Microb. Cell Fact. 2020, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Arpigny, J.L.; Jaeger, K.E. Bacterial lipolytic enzymes: Classification and properties. Biochem. J. 1999, 343, 177–183. [Google Scholar] [CrossRef]
- Hitch, T.C.A.; Clavel, T. A proposed update for the classification and description of bacterial lipolytic enzymes. PeerJ 2019, 7, e7249. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Fu, S.; Liu, H.; Lucia, L.A. Accurately determining esterase activity via the isosbestic point of p-nitrophenol. BioResources 2016, 11, 10099–10111. [Google Scholar] [CrossRef]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 22–25. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Y.F.; Ewis, H.E.; Abdelal, A.T.; Lu, C.D.; Harrison, R.W.; Weber, I.T. Covalent reaction intermediate revealed in crystal structure of the Geobacillus stearothermophilus carboxylesterase Est30. J. Mol. Biol. 2004, 342, 551–561. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ScEst | |
|---|---|
| Source organism | Staphylococcus chromogenes strain NCTC10530 |
| DNA source | Genomic DNA |
| Cloning vector | pVFT1S |
| Expression host | Escherichia coli BL21(DE3) |
| Amino acid sequence | MKIQLPKPFLFEEGKRAVLLLHGFTGNSSDVRQLG RFLQKKGYTSYAPHYEGHAAPPEEILKSSPHVWY KDALDGYDYLVDKGYDEIAVAGLSLGGVFALKLS LNRDVKGIVTMCSPMYIKTEGSMYEGVLEYARNF KKYEGKDETTIEREMQAFHPTSTLRELQETIQSV RDHVEDVIEPLLVIQAEQDEMINPDSANVIYNEA ASDEKHLSWYKNSGHVITIDKEKEDVFEEVYQFLESLDWSE |
| Method | Vapor Diffusion |
|---|---|
| Plate type for screening | 96-well sitting drop MRC plate (Molecular dimension, UK) |
| Composition of reservoir solution | 0.2 M Ammonium citrate tribasic (pH 7.0), 20% PEG 3350 |
| Plate type for optimization | 24-well hanging drop plate, (Molecular dimension, UK) |
| Composition of optimal solution | 0.25 M Ammonium citrate tribasic (pH 7.0), 20% PEG 3350 |
| Temperature (K) | 296 |
| Protein concentration (mg/mL) | 4.3 |
| Composition of protein solution | 20 mM Tris-HCl (pH 8.0), 200 mM NaCl |
| Volume and ratio of drop (protein: solution) | 2.0 μL, 1:1 |
| Volume of reservoir (μL) | 500 |
| Data Collection | |
|---|---|
| Wavelength (Å) | 0.9793 |
| X-ray source | PAL 7A |
| Rotation range per image (°) | 1 |
| Exposure Time (s) | 1 |
| Space group | P212121 |
| Unit-cell parameters (Å, °) | a = 50.23, b = 68.69, c = 71.15 α = 90, β = 90, γ = 90 |
| Resolution range (Å) a | 50–1.66 (1.69–1.66) |
| No. of observed reflections a | 402,244 (19,564) |
| No. of unique reflections a | 29,406 (1471) |
| Completeness (%) a | 99.3 (100) |
| Redundancy a | 13.7 (13.3) |
| Rsyma,b | 0.112 (1.202) |
| Rmeasc | 0.117 (1.250) |
| I/σa | 62.3 (4.0) |
| CC(1/2) (%) | 99.6 (82.8) |
| Wilson B factor (Å2) | 24.66 |
| Matthews coefficient | 2.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, J.; Jeon, S.; Lee, M.J.; Yoo, W.; Chang, J.; Kim, K.K.; Lee, J.H.; Do, H.; Kim, T.D. Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Novel Esterase (ScEst) from Staphylococcus chromogenes. Crystals 2022, 12, 546. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12040546
Hwang J, Jeon S, Lee MJ, Yoo W, Chang J, Kim KK, Lee JH, Do H, Kim TD. Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Novel Esterase (ScEst) from Staphylococcus chromogenes. Crystals. 2022; 12(4):546. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12040546
Chicago/Turabian StyleHwang, Jisub, Sangeun Jeon, Min Ju Lee, Wanki Yoo, Juwon Chang, Kyeong Kyu Kim, Jun Hyuck Lee, Hackwon Do, and T. Doohun Kim. 2022. "Identification, Characterization, and Preliminary X-ray Diffraction Analysis of a Novel Esterase (ScEst) from Staphylococcus chromogenes" Crystals 12, no. 4: 546. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst12040546