1. Introduction
Grafting has become an effective practice in the production of high-value solanaceous and cucurbitaceous vegetables to help overcome biotic and abiotic stresses and improve crop productivity [
1,
2,
3]. Although grafting also has been used as a tool in plant physiology studies of Arabidopsis (
Arabidopsis thaliana L.) [
4], for accelerating the breeding work of common beans (
Phaseolus vulgaris L.) [
5], and for combating Verticillium wilt of globe artichoke (
Cynara cardunculus L. subsp.
Scolymus) [
6], grafting in other vegetable species beyond Solanaceae and Cucurbitaceae is generally not practiced commercially. Interestingly, some attempts have been made to explore the feasibility of grafting vegetable plants in
Brassicaceae. Oda et al. [
7] tested inter-varietal, inter-specific, and inter-generic grafting among cabbage (
Brassica oleracea L. var.
capitata), kale (
Brassica oleracea var.
sabellica), kohlrabi (
Brassica oleracea var.
gongylodes), Chinese cabbage (
Brassica rapa L. subsp.
pekinensis), turnip (
Brassica rapa subsp.
rapa), Japanese mustard (Takana) (
Brassica juncea L. var.
integrifolia), and Japanese radish (
Raphanus sativus L. var.
longipinnatus) and obtained successful grafts. Particularly, an adhesive and hardener system was developed for making grafts between Chinese cabbage (scion) and turnip (rootstock) [
8]. Recently, Chen et al. [
9] evaluated the survival rate of cabbage grafted onto Chinese kale (
B. oleracea Alboglabra group) rootstocks and assessed the feasibility of using grafting to improve cabbage head quality.
The effort of cruciferous vegetable grafting has not only broadened the potential use of vegetable grafting as a management tool, but also presents the possibility of creating a novel vegetable product with added value. In the case of grafted Chinese cabbage/turnip plants, the above-ground portion of Chinese cabbage—a leafy vegetable, and the below-ground portion of turnip—a root vegetable, can be harvested from the same plant. This type of rootstock–scion combination holds promise for space saving in small-scale intensive cultivation systems. Moreover, the grafted Chinese cabbage/turnip vegetable may possess added economic value with minimal waste, since many consumers may prefer not to eat turnip leaves and could be drawn to the novelty of this new product. However, the Chinese cabbage and turnip grafting study of Oda and Nakajima [
8] only reported a 50% graft survival rate and observed restricted development of the Chinese cabbage head.
In this proof of concept study, we grafted the pac choi (
B. rapa var.
chinensis) scion onto the daikon radish (
R. sativus var.
longipinnatus) rootstock to generate a vegetable plant that produced a pac choi leafy vegetable above-ground and an edible daikon radish root below-ground. Pac choi and daikon radish are among specialty vegetables increasingly grown for local markets in the U.S. Although edible, daikon radish leaves are often discarded at consumption. Recent genetic studies supported the feasibility of making successful inter-generic grafts between
B. rapa and
R. sativus. Yang et al. [
10] sequenced the chloroplast noncoding region and found that
R. sativus was closely related to
B. rapa/oleracea and proposed that
Raphanus was derived from hybridization between
B. rapa/oleracea and
B. nigra, the two evolutionary lineages in the genus Brassica. Furthermore, the reciprocal hybridization between
R. sativus and
B. rapa has been proven viable [
11]. Vigorous growth was also observed for most of the successful primary hybrids between
B. rapa and
R. sativus [
12]. On the other hand, according to Tonosaki et al. [
13], when hybridized with
R. sativus, only one particular breeding line of
B. rapa (‘Shogoin-kabu’) successfully produced hybrid seeds, whereas most other lines failed due to embryo breakdown.
By grafting the pac choi scion onto the daikon radish rootstock, the objectives of this pilot experiment were to examine the feasibility of developing successful grafts for harvesting both pac choi leaves and daikon radish taproot from the same plant, and to compare the growth and development of grafted plants with self-grafted and non-grafted pac choi and daikon radish plants.
2. Materials and Methods
Two experiments were carried out in this study. The first experiment was a pilot study to test the feasibility of grafting pac choi onto daikon radish. The second experiment was intended to provide a better understanding of above-ground growth and below-ground development of this unique scion-rootstock system over an extended post-grafting period of plant establishment. In both experiments, ‘Bora King’ (BK), a daikon radish with purple taproots (Johnny’s Selected Seeds, Winslow, ME, USA) was used as the rootstock, while ‘Mei Qing Choi’ (MQ) pac choi (Johnny’s Selected Seeds) was used as the scion. They were selected based on our preliminary study in which these two cultivars were found to be compatible for grafting and have similar hypocotyl diameters.
2.1. Setup of the Pilot Experiment
Pac choi and daikon radish were seeded on 7 and 13 November 2016, respectively. The pac choi was seeded 6 d earlier than the daikon radish in order to match the stem diameter of the seedlings at grafting, as the daikon radish germinated and emerged much quicker than the pac choi based on a preliminary seeding test. All the seeds were sown in 72-cell Speedling trays (Speedling Inc., Ruskin, FL, USA) and filled with Fafard-2 potting mix (Sun Gro Horticulture, Agawam, MA, USA) containing a mixture of peat moss, perlite, vermiculite, and dolomite lime. Plants were grown in a greenhouse at the University of Florida campus (Gainesville, FL, USA). Water-soluble fertilizer 20N-8.7P-16.7K (Jack’s Classic; Jr Peters Inc., Allentown, PA, USA) was applied on 17 and 28 November at a nitrogen (N) concentration of 200 mg L−1.
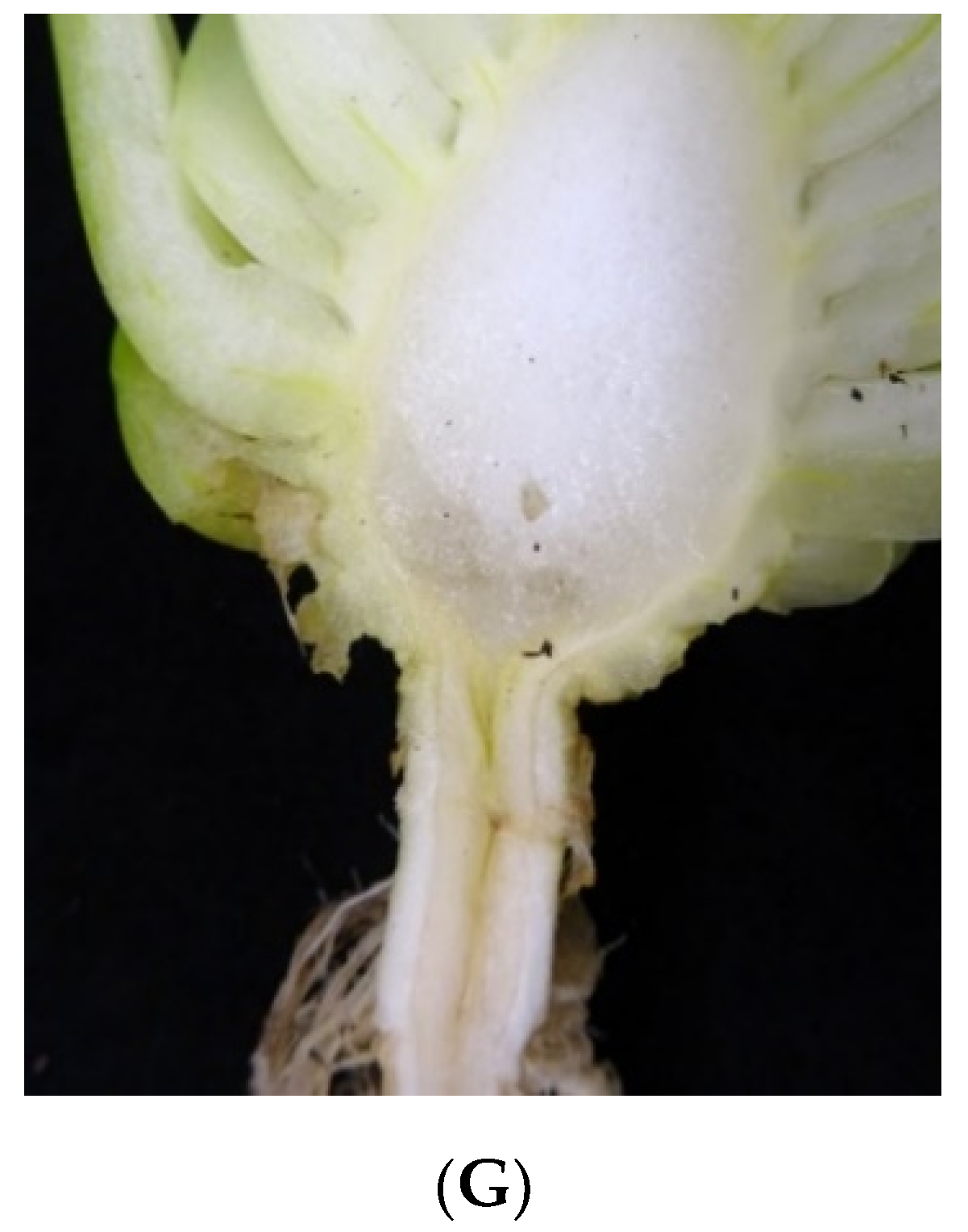
Plants were grafted on 30 November 2016 (0 d after grafting (DAG)) using the splice grafting method [
1]. Twenty-four plants were grafted using seedlings with the most consistent growth. With the purpose of ensuring consistent grafting quality, only a small number of plants were grafted in this pilot experiment after earlier attempts at practicing the grafting technique. The daikon radish seedlings were severed using a double edge razor blade at approximately a 45-degree angle below cotyledons to remove the shoots, with pac choi scions cut at the hypocotyl with the same angle just above the soil surface. The cut surfaces of the pac choi scion and the daikon radish seedlings with shoot removal were conjoined using a 1.5 mm silicone grafting clip (Johnny’s Selected Seeds). Grafted plants were then placed in a healing chamber constructed by wrapping a metal shelving unit with thin plastic film (Uline Econo-Wrapper (0.02 mm), Uline corporation, Pleasant Prairie, WI, USA) in a temperature-controlled room with air temperature set at 23 °C and relative humidity (RH) at 99%. Light was provided by two, 54-watt T5 fluorescent lights (Philips Lighting Company, Somerset, NJ, USA) at a photosynthetic photon flux density (PPFD) of 56 μmol m
−2 s
−1 at seedling canopy level for 12 h each day. An additional plastic tray with a wet sheet of germination paper was placed inside the healing chamber to help maintain humidity.
From 5 DAG, the healing chamber was gradually cut open and the ambient RH setting was reduced to 60%. At 8 DAG, the plastic film was completely removed, and grafts remained exposed in the temperature-controlled room until 13 DAG. Water was applied to plants by filling the bottom of the tray for absorption. All the grafted plants were transferred to a greenhouse at 13 DAG, and graft survival rate was determined by counting the number of live and dead plants; only plants with turgid leaves were counted as living. At harvest, the number of surviving grafted plants was counted again for calculation of the final graft survival rate, as some plants severely declined following transplanting into pots.
At 16 DAG, surviving grafted plants were transplanted into 11.36 L black plastic pots (1200C; Hummert International, Earth City, MO, USA) filled with Fafard-2 soilless mix (Sun Gro Horticulture) for continued monitoring of the survival of the grafted plants. In addition, five plants of non-grafted ‘Mei Qing Choi’ and ‘Bora King’ were potted as controls. All the plants were placed on the greenhouse bench following a completely randomized design. Organic fertilizer MicroSTART60 3N-0.9P-2.5K (Perdue AgriRecycle, LLC., Seaford, DE, USA) was applied to each pot at the rate of 80 g/pot. Drip irrigation was used by placing one 1.89 L h−1 emitter (Woodpecker pressure compensating junior dripper; Netafim USA, Fresno, CA, USA) in each pot; plants were watered once a day for 3 min. Irrigation increased to twice per day for 2 min each time starting at 56 DAG. Insecticidal soap (Safer Brand; Woodstream Corporation, Lancaster, PA, USA) was sprayed at 56 DAG and lacewing larvae (Chrysoperla rufilabris (Neuroptera: Chrysopidae); Rincon-Vitova Insectaries, Ventura, CA, USA) were released at 64 DAG for aphid control. The average day and night temperatures of the greenhouse during the plant growth were 22.8 °C and 16.5 °C, respectively.
2.2. Setup of the Follow-Up Experiment
A follow-up experiment was conducted in 2019 to further explore the above-ground growth and below-ground taproot development in grafted pac choi–daikon radish plants. ‘Mei Qing Choi’ (MQ) pac choi was grafted onto ‘Bora King’ (BK) daikon radish (MQ/BK), while non-grafted pac choi (MQ) and daikon radish (BK) as well as self-grafted pac choi (MQ/MQ) and daikon radish (BK/BK) were used as controls. A randomized complete block design with four replications (blocks) and ten grafted plants per treatment per replication (block) was used in the grafting experiment. MQ and BK were seeded into 72-cell trays on 8 and 14 February 2019, respectively. Fish & seaweed organic liquid fertilizer 2N-1.3P-0.8K (Neptune’s Harvest, Gloucester, MA, USA) and 0N-0P-41.5K potassium sulfate (Big K; JHBiotech, Inc., Ventura, CA, USA) were applied at concentrations of 200 mg L−1 N and 200 mg L−1 K2O at the seedling growth stage on 18 and 26 Feb. Plants were grafted on 1 March 2019 using the aforementioned grafting method. Grafted plants were healed in an air-conditioned laboratory room with the same set up of healing chamber as in 2016. Supplemental light was provided for 10 h each day during the healing process. The air temperature and RH of the laboratory room were 23.8 ± 0.5 °C and 47.6 ± 13.1%. The plastic film was completely removed at 8 DAG. Water was gently sprayed onto the soil surface using a wash bottle when needed to avoid wetting foliage. All the grafted plants were moved into a greenhouse at 13 DAG where the average day and night temperatures were 27.1 °C and 20.7 °C, respectively. Graft survival rate was determined for each grafting treatment in each replication by counting the number of live and dead plants at 17 DAG. At 19 DAG, 24 plants from each treatment with healthy and consistent growth were chosen and randomly reassigned to four blocks with six plants in each block for further evaluation of the growth of the grafted plants in a greenhouse pot study, following a randomized complete block design. Plants were transplanted into 11.36 L black plastic pots filled with PRO-MIX premium organic vegetable and herb mix (Premier Tech Ltd., Quakertown, PA, USA) which contained 60–75% peat moss plus peat humus, compost, perlite, gypsum, limestone, organic fertilizer, and mycorrhizae. Drip irrigation was used by placing one 1.89 L h−1 emitter in each pot; the plants were watered twice a day for 3 min per cycle between 21 and 39 DAG and irrigation increased to 4 min per cycle thereafter. Adventitious roots developed from the graft union area were monitored and removed once a week as needed after the plants were transplanted into the pots.
2.3. Plant Growth Measurements
In the 2019 follow-up experiment, leaf relative chlorophyll content and canopy size were measured at 33 and 41 DAG. A SPAD 502 Plus Chlorophyll Meter (Spectrum Technologies, Aurora, IL, USA) was used to measure leaf relative chlorophyll content on three randomly chosen plants per treatment per block by averaging four readings obtained from two distal areas of the leaf blade for each of the two most recent mature leaves per plant. The canopy size was measured on 3 plants of MQ/BK, MQ/MQ, and MQ for each block using digital photographs processed with ImageJ/Fiji (version 2.0.0) [
14]. A ruler held in the frame of each photograph set the scale for pixels per linear cm and enabled digital measurement of length and width of the plant canopy. The canopy size was then determined by multiplying the canopy length and width.
2.4. Gas Exchange Measurements
Gas-exchange was measured in the 2019 follow-up experiment at 34 and 46 DAG between 10:00 am and 3:00 pm by using an open gas exchange system (Li-6800; Li-Cor Inc., Lincoln, NE, USA) on three plants per treatment per block. Leaf transpiration rate (E, mmol H
2O m
−2 s
−1), net CO
2 assimilation rate (A, mmol CO
2 m
−2 s
−1), intercellular CO
2 concentration (Ci, μmol CO
2 mol
−1 air), and stomatal conductance to water (gsw, mmol H
2O m
−2 s
−1) were measured at steady-state on the third (fully expanded) leaf from the top of each plant [
15]. The PPFD was set at 800 μmol m
−2 s
−1, with CO
2 concentration at 400 ppm, vapor pressure deficit at 1.2 kPa, and leaf temperature at 27–29 °C. Instantaneous water use efficiency (iWUE) (µmol CO
2 mmol
−1 H
2O) was calculated as A/E [
16] and stomatal conductance (Gs, mol m
−2 s
−1) was calculated as gsw/1.6 [
17].
2.5. Yield Components and Biomass Accumulation at Harvest
For the 2016 pilot study, the above-ground part (above soil line) of all the plants of MQ/BK, non-grafted MQ, and non-grafted BK were harvested at 68 DAG. The number of leaves longer than 4 cm were counted for each plant. The MQ/BK and non-grafted BK were then uprooted, and the taproots were separated and rinsed with water to remove excess potting soil from the roots. Taproot length (from the stem base to the end of the radish taproot) of each harvested plant was recorded, and the diameter of the widest part of each radish taproot was measured with a digital caliper. For the 2019 follow-up experiment, harvest and destructive sampling were carried out at 47 DAG. Five out of six plants per treatment per replication were randomly sampled. The above-ground part (above soil line) of each plant was removed from the pot, and leaves longer than 4 cm were counted and scanned with a leaf area meter (LI-3100; Li-Cor Inc., Lincoln, NE, USA). Only taproots from MQ/BK, BK, and BK/BK were harvested and cleaned. Pak choi and daikon radish leaves and taproots from the 2016 and 2019 experiments were first weighed then dried at 65 °C for 7 d (until constant weight) to determine the above-ground and below-ground fresh and dry biomass.
2.6. Mineral Nutrient Contents in Leaf and Root Tissues
In the 2019 experiment, the dried root samples from the BK, BK/BK, and MQ/BK treatments and dried leaf samples from the MQ, MQ/MQ, and MQ/BK treatments were ground using a Thomas Wiley Laboratory mill (Model 4; Arthur H. Thomas Company, Philadelphia, PA, USA) and sent to Waters Agricultural Laboratories (Camilla, GA, USA) to measure the concentrations of the macronutrients N, phosphate (P), sulfate (S), potassium (K), calcium (Ca), and magnesium (Mg) and the micronutrients boron (B), zinc (Zn), manganese (Mn), iron (Fe), and copper (Cu). Nutrient accumulation was calculated by multiplying the nutrient concentration by dry tissue biomass.
2.7. Statistical Analyses
The pilot study followed a completely randomized design with five replications and one plant per replication. In the follow-up experiment, a randomized complete block design with four replications (blocks) and ten plants per experimental unit was used before plants were transplanted to larger pots, when the number was reduced to six plants per experimental unit. Data were analyzed using a linear mixed model in the GLIMMIX procedure of SAS (SAS Version 9.4 for Windows; SAS Institute, Cary, NC, USA). Some data were transformed by taking the square root to meet the assumptions of the model (normality, homogeneity, linearity) as needed, while results were presented using the original data following statistical analysis. Fisher’s least significant difference (LSD) test (α = 0.05) was conducted for multiple comparisons of different measurements among treatments.
4. Conclusions
Successful grafts were produced between B. rapa var. chinensis and R. sativus var. longipinnatus in this study, resulting in a novel ‘pac choi–daikon radish’ product that may help save growing space and have added-value as perceived by farmers and consumers. More research is needed to optimize the seeding time and management of seedling production to help further improve the graft survival rate. Grafting pac choi with daikon radish did not severely impair the growth of the above-ground parts as grafted pac choi had similar SPAD value, canopy size, leaf number, and above-ground DW compared with non-grafted pac choi. Interestingly, grafting with radish increased the photosynthetic ability of the pac choi. However, grafting the daikon radish with pac choi decreased the taproot formation as reflected by the reduced length, diameter, FW and DW of the taproot. Future studies could explore different approaches such as cultivar selection and nutrient management to better balance the sizes of the above- and below-ground parts of this new pac choi–daikon radish product. Given the wide range of B. rapa var. chinensis and R. sativus var. longipinnatus cultivars, it would be interesting to explore different grafting combinations to characterize the range of graft performance. We only tested the graft performance under greenhouse conditions, and the grafted plants need to be further evaluated in field growing systems where biotic and abiotic stressors can be intensified. Generally, grafting between pac choi and daikon radish showed more negative impacts on mineral nutrient levels in radish taproots than in pac choi leaves. Sensory properties of the ‘pac choi–daikon radish’ product are unknown, and this aspect deserves further assessment. The inter-generic grafting between B. rapa var. chinensis and R. sativus var. longipinnatus could also provide a unique model system to further our understanding of scion-rootstock synergy and above- and below-ground sink competition in horticultural crops toward improving the use of grafting technology in sustainable vegetable production.





{kind=link}
{kind=link}