Nutritional and Visual Diagnosis in Broccoli (Brassica oleracea var. italica L.) Plants: Disorders in Physiological Activity, Nutritional Efficiency and Metabolism of Carbohydrates

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Site
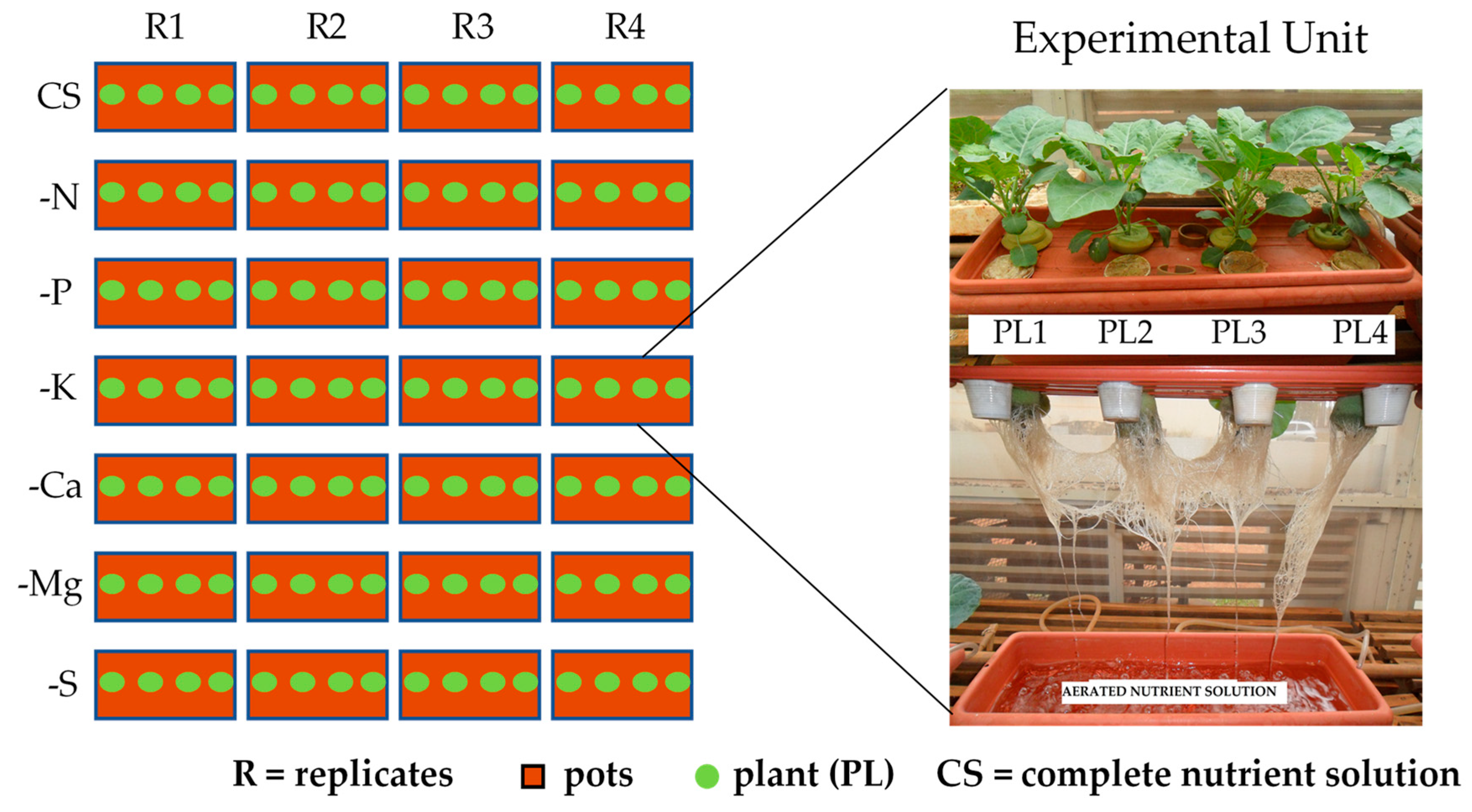
2.2. Experimental Design and Treatments
2.3. Conducting the Experiment
2.4. Growth Evaluations and SPAD (Soil Plant Analysis Development) Reading
2.5. Evaluation of Macronutrient Content and Accumulation
2.6. Nutrient Absorption, Transport and Use Efficiency Indexes
2.7. Gas Exchange Evaluation
2.8. Content of Starch and Total Soluble Sugars (SS)
2.9. Data Analysis
3. Results
3.1. Visual Diagnosis
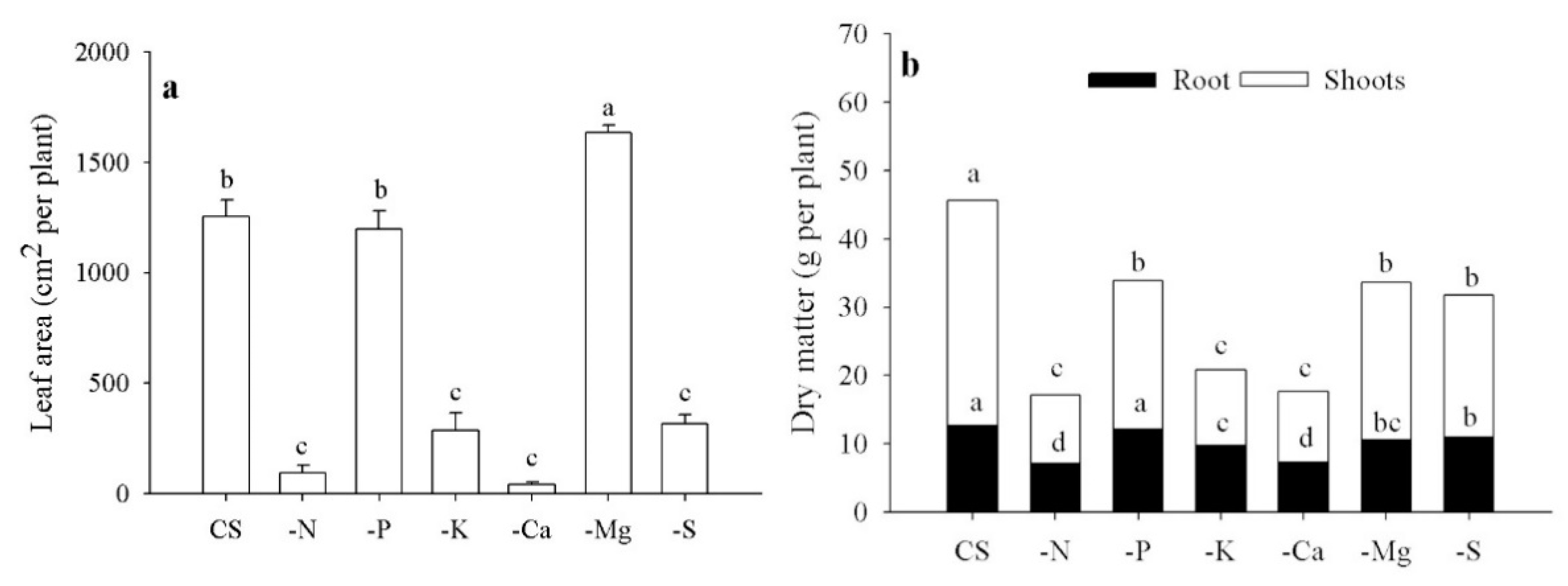
3.2. Growth Evaluations and SPAD Readings
3.3. Macronutrient Content and Accumulation
3.4. Efficiency Indexes
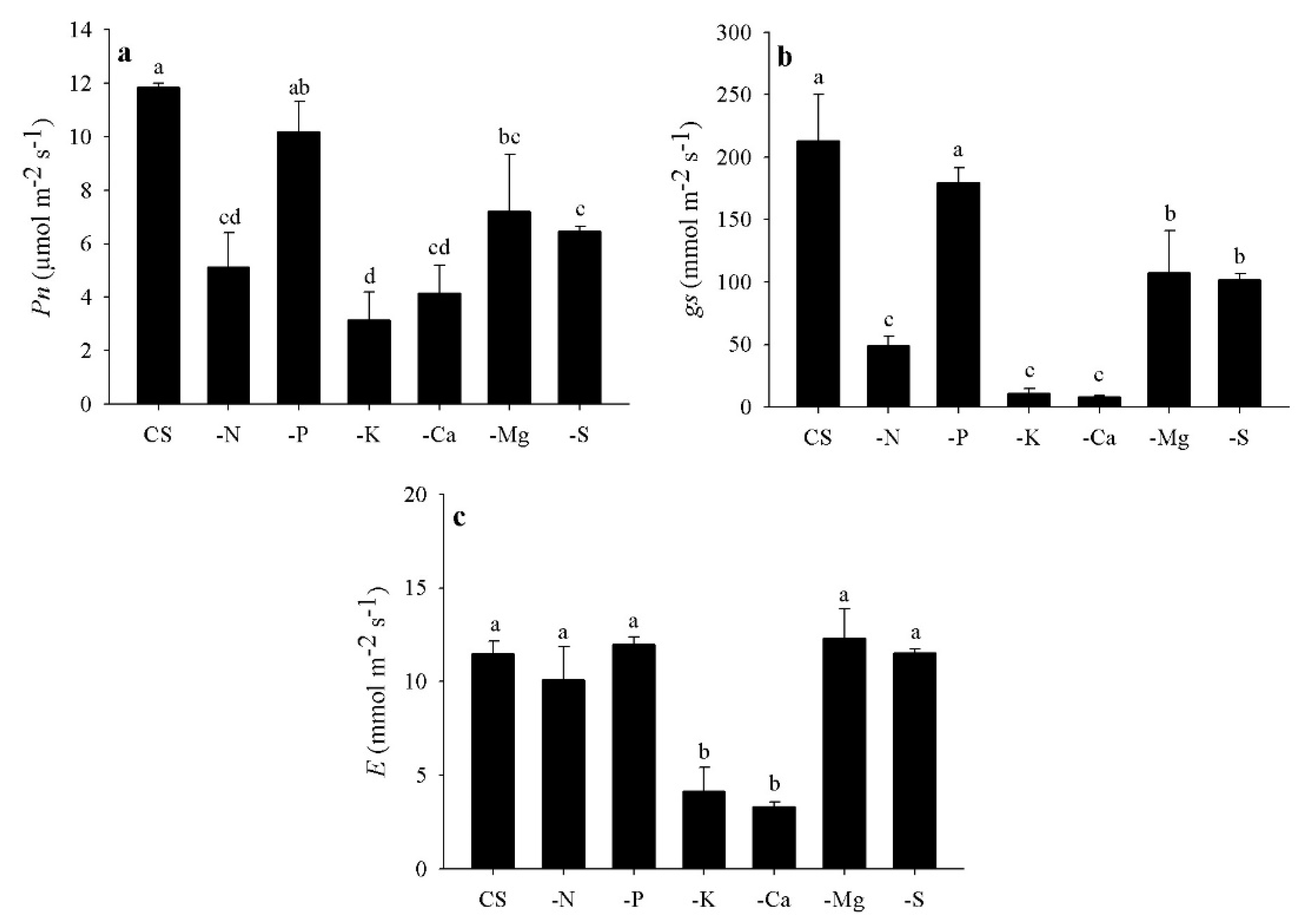
3.5. Gas Exchange
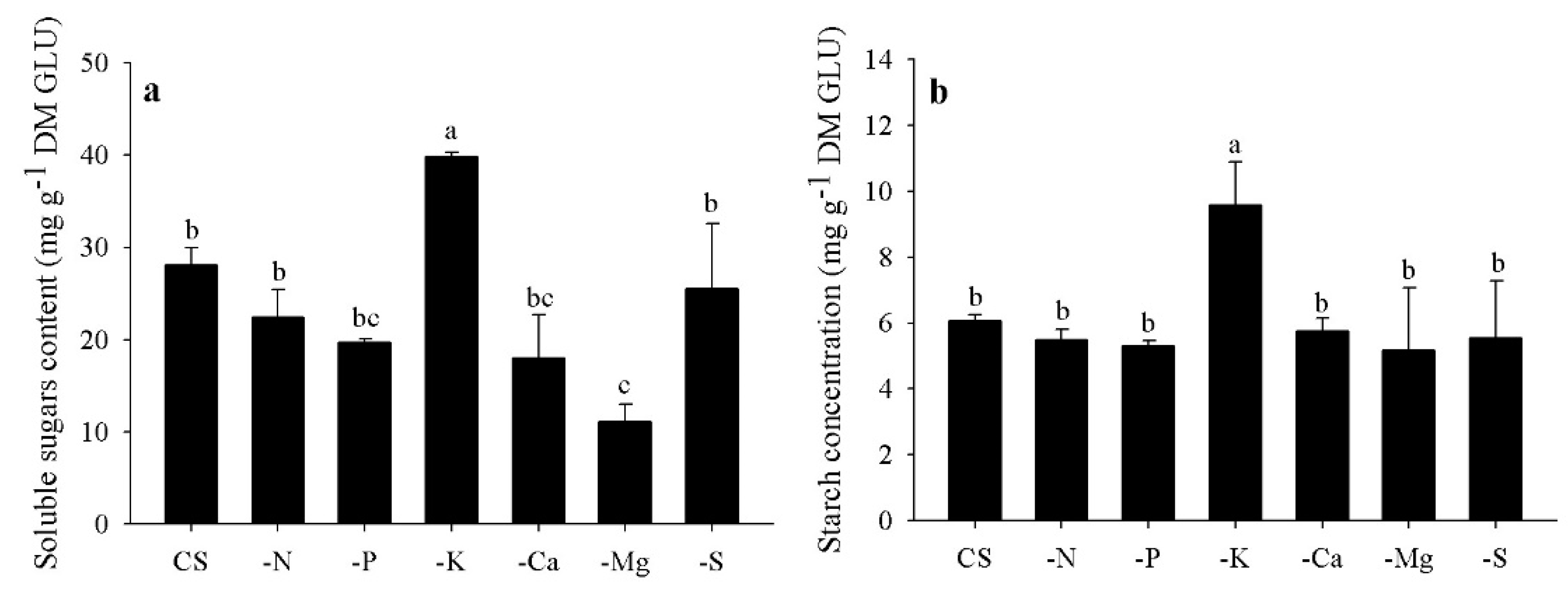
3.6. Starch and Soluble Sugars (SS) Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO; ITPS. Status of the World’s Soil Resources (SWSR)—Main Report. Food and Agriculture Organization of the United Nations and Intergovernmental Technical Panel on Soils. 2015. Available online: http://www.fao.org/3/a-i5228e.pdf (accessed on 13 March 2015).
- Bianco, M.S.; Filho, A.B.C.; Carvalho, L.B. Nutritional status of the cauliflower cultivar ‘Verona’ grown with omission of out added macronutrients. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal; Artmed: Porto Alegre, Brazil, 2017; p. 889. [Google Scholar]
- Schjoerring, J.K.; Cakmak, I.; White, P.J. Plant nutrition and soil fertility: Synergies for acquiring global green growth and sustainable development. Plant Soil 2019, 434, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Berry, W. Symptoms of deficiency in essential minerals. In Plant Physiology Online; Taiz, L., Zeiger, E., Eds.; The American Society of Plant Biologists: Rockville, MD, USA, 2016; Available online: http://6e.plantphys.net/topic05.01.html (accessed on 3 October 2016).
- Cavalcante, V.S.; Prado, R.M.; Vasconcelos, R.L.; Almeida, H.J.; Silva, T.R. Growth and Nutritional Efficiency of Watermelon Plants Grown under Macronutrient Deficiencie. HortScience 2019, 54, 738–742. [Google Scholar] [CrossRef] [Green Version]
- Bouain, N.; Krouk, G.; Lacombe, B.; Rouached, H. Getting to the root of plant mineral nutrition: Combinatorial nutrient stresses reveal emergent properties. Trends Plant Sci. 2019, 24, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Carmona, V.V.; Costa, L.C.; Filho, A.B.C. Symptoms of nutrient deficiencies on cucumbers. Int. J. Plant Soil Sci. 2015, 8, 1–11. [Google Scholar] [CrossRef]
- Cavalcante, V.S.; Prado, R.M.; Almeida, H.J.; Cruz, F.J.R.; Santos, D.M.M. Gaseous exchanges, growth and foliar anatomy of sugarcane plants grown in potassium (K) deprived nutrient solution. Aust. J. Crop Sci. 2015, 9, 577–584. [Google Scholar]
- Santos, E.F.; Macedo, F.G.; Zanchim, B.J.; Lima, G.P.P.; Lavres, J. Prognosis of physiological disorders in physic nut to N, P, and K deficiency during initial growth. Plant Physiol. Biochem. 2017, 115, 249–258. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Database, Crops. 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 4 April 2019).
- Bachiega, P.; Salgado, J.M.; Carvalho, J.E.; Ruiz, A.L.T.G.; Schwarz, K.; Tezotto, T.; Morzelle, M.C. Antioxidant and antiproliferative activities in different maturation stages of broccoli (Brassica oleracea Italica) biofortified with selenium. Food Chem. 2016, 190, 771–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, E.; Bagci, P.O. Production of phytotherapeutics from broccoli juice by integrated membrane processes. Food Chem. 2018, 242, 264–271. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.L. The Water Culture Methods for Growing Plants without Soil; California Agricultural Experiment Station: Berkeley, CA, USA, 1950; p. 39. [Google Scholar]
- Boaretto, A.E.; van Raij, B.; Silva, F.C.; Chitolina, J.C.; Tedesco, M.J.; Carmo, C.A.F.S. Amostragem, acondicionamento e preparo de amostras de plantas para análise química. In Manual de Análises Químicas de Solos, Plantas e Fertilizantes; Silva, F.C., Ed.; EMBRAPA: Brasília, Brazil, 2009; pp. 59–86. [Google Scholar]
- Miyazawa, M.; Pavan, M.A.; Muraoka, T.; Carmo, C.A.F.S.; Melo, W.J. Análises químicas de tecido vegetal. In Manual de Análises Químicas de Solos, Plantas e Fertilizantes; Silva, F.C., Ed.; EMBRAPA: Brasília, Brazil, 2009; pp. 191–233. [Google Scholar]
- Swiader, J.M.; Chyan, Y.; Freiji, F.G. Genotypic differences in nitrate uptake and utilization efficiency in pumpkin hybrids. J. Plant Nutr. 1994, 17, 1687–1699. [Google Scholar] [CrossRef]
- Li, B.; Mckeand, S.E.; Allen, H.L. Genetic variation in nitrogen use efficiency of loblolly pine seedlings. For. Sci. 1991, 37, 613–626. [Google Scholar]
- Siddiqi, M.Y.; Glass, A.D. Utilization index: A modified approach to the estimation and comparison of nutrient utilization efficiency in plants. J. Plant Nutr. 1981, 4, 289–302. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Moller, J.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 135–189. [Google Scholar]
- Costa, L.C.; Carmona, V.M.V.; Filho, A.B.C.; Nascimento, C.S. Sintomas de deficiências de macronutrientes em melancieira. Commun. Sci. 2017, 8, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Pôrto, M.L.; Puiatti, M.; Fontes, P.C.R.; Cecon, P.R.; Alves, J.C.; Arruda, J.A. Índice SPAD para o diagnóstico do estado de nitrogênio na cultura da abobrinha. Hortic. Bras. 2011, 29, 311–315. [Google Scholar] [CrossRef]
- Garcez, T.B.; Megda, M.M.; Artur, A.G.; Monteiro, F.A. Production and Spad reading in Marandu palisadegrass fertilized with nitrogen and magnesium. J. Plant Nutr. 2014, 37, 1475–1486. [Google Scholar] [CrossRef]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, S.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecílio Filho, A.B.; Carmona, V.M.V.; Schiavon, A.A., Jr. Broccoli growth and nutrient accumulation. Científica 2017, 45, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, B.; Nahar, K.; Hossain, S.; Al-Mahmud, J.; Hossen, S.; Masud, A.A.C.; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Gerardeaux, E.; Jordan-Meille, L.; Constantin, J.; Pellerin, S.; Dingkuhn, M. Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ. Exp. Bot. 2010, 67, 451–459. [Google Scholar] [CrossRef]
- Chen, C.-T.; Lee, C.-L.; Yeh, D.-M. Effects of nitrogen, phosphorus, potassium, calcium, or magnesium deficiency on growth and photosynthesis of Eustoma. HortScience 2018, 53, 795–798. [Google Scholar] [CrossRef]
- Gransee, A.; Führs, H. Magnesium mobility in soils as a challenge for soil and plant analysis, magnesium fertilization and root uptake under adverse growth conditions. Plant Soil. 2013, 368, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Hermans, C.; Verbruggen, N. Physiological characterization of Mg deficiency in Arabidopsis thaliana. J. Exp. Bot. 2005, 56, 2153–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermans, C.; Bourgis, F.; Faucher, M.; Strasser, R.J.; Delrot, S.; Verbruggen, N. Magnesium deficiency in sugar beets alters sugar partitioning and phloem loading in young mature leaves. Planta 2005, 220, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Farhat, N.; Elkhouni, A.; Zorrig, W.; Smaoui, A.; Abdelly, C.; Rabhil, M. Effects of magnesium deficiency on photosynthesis and carbohydrate partitioning. Acta Physiol. Plant. 2016, 38, 145–155. [Google Scholar] [CrossRef]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Lunde, C.; Zygadlo, A.; Simonsen, H.T.; Nielsen, P.L.; Blennow, A.; Haldrup, A. Sulfur starvation in rice: The effect on photosynthesis, carbohydrate metabolism, and oxidative stress protective pathways. Physiol. Plant. 2008, 134, 508–521. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 9 | Stock Solution | CS | -N | -P | -K | -Ca | -Mg | -S |
|---|---|---|---|---|---|---|---|---|
| Stock Solution Volume Per L of Final Solution | ||||||||
| mol L−1 | ---------------------------- mL L−1--------------------------------- | |||||||
| KH2PO4 | 1.0 | 1 | 1 | - | - | 1 | 1 | 1 |
| KNO3 | 1.0 | 5 | - | 5 | - | 5 | 3 | 3 |
| Ca(NO3) 5H2O | 1.0 | 5 | - | 5 | 5 | - | 4 | 4 |
| MgSO4 7H2O | 1.0 | 2 | 2 | 2 | 2 | 2 | - | - |
| KCl | 1.0 | - | 5 | 1 | - | - | 2 | 2 |
| CaCl2 2H2O | 1.0 | - | 5 | - | - | - | 1 | 1 |
| NH4H2PO4 | 1.0 | - | - | - | 1 | 5 | - | - |
| NH4NO3 | 1.0 | - | - | - | 2 | - | - | - |
| (NH4)2SO4 | 1.0 | - | - | - | - | - | 2 | - |
| MgNO3 6H2O | 1.0 | - | - | - | - | - | - | 2 |
| Micronutrients | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| Fe EDDHA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| Treat. | Equation | DATD (a) | ValueD (b) | DATmax (c) | Valuemax (d) | R2 |
|---|---|---|---|---|---|---|
| Plant Height (cm) | ||||||
| CS | y = 0.91824672 + 1.12909052x | - | - | 38 | 45.0 | 0.99 ** |
| -N | y = 4.20130816 + 0.55908387x − 0.01824500x2 | 12 | 8.3 | 15 | 8.5 | 0.96 ** |
| -P | y = 6.42223729 + 0.72724158x | 32 | 29.7 | 38 | 36.0 | 0.87 ** |
| -K | y = -3.2585786 + 1.84660483x − 0.03459323x2 | 22 | 20.6 | 27 | 21.4 | 0.94 ** |
| -Ca | y = 1.21979350 + 1.28588367x − 0.03578473x2 | 17 | 12.7 | 18 | 12.8 | 0.94 ** |
| -Mg | y = -0.1353877 + 1.39611512x − 0.01845996x2 | 32 | 25.6 | 38 | 26.3 | 0.94 ** |
| -S | y = 7.49758709 + 0.48716448x | 24 | 19.2 | 38 | 25.0 | 0.90 ** |
| Number of leaves | ||||||
| CS | y = 8.44067676 + 0.31460594x | - | - | 38 | 20.0 | 0.87 ** |
| -N | y = 2.41799644 + 0.86095054x − 0.02517633x2 | 12 | 9.2 | 17 | 9.7 | 0.96 ** |
| -P | y = 4.71305113 + 0.70604835x − 0.01074269x2 | 32 | 16.3 | 32 | 16.3 | 0.87 ** |
| -K | y = 0.50041459 + 1.33187830x − 0.03202433x2 | 22 | 14.3 | 21 | 14.3 | 0.99 ** |
| -Ca | y = 3.97384343 + 0.67795485x − 0.02148872x2 | 17 | 9.3 | 16 | 9.3 | 0.93 ** |
| -Mg | y = 1.72788326 + 0.91283468x − 0.01472306x2 | 32 | 16.0 | 31 | 16.0 | 0.95 ** |
| -S | y = 7.92279412 + 0.23308824x | 24 | 13.5 | 38 | 17.5 | 0.77 ** |
| SPAD readings | ||||||
| CS | y = 28.1581834 + 0.48175923x | - | - | 38 | 50.6 | 0.45 ** |
| -N | y = 36.5755164 − 0.95082810x | 12 | 34.9 | 07 | 35.0 | 0.89 ** |
| -P | y = 19.3321594 + 02.13131758x − 0.03850833x2 | 32 | 48.0 | 38 | 48.8 | 0.41 ** |
| -K | y = -21.525585 + 7.81561651x − 0.18668696x2 | 22 | 60.0 | 21 | 60.3 | 0.93 ** |
| -Ca | y = 34.0619555 + 1.81493501x − 0.06720483x2 | 17 | 45.4 | 14 | 46.3 | 0.82 ** |
| -Mg | y = 34.6999518 − 0.43722765x + 0.02425081x2 | 32 | 45.5 | 32 | 45.5 | 0.62 * |
| -S | y = 34.1812500 | 24 | - | - | - | ns |
| Treat. | N | P | K | Ca | Mg | S |
|---|---|---|---|---|---|---|
| Shoot | ||||||
| ________________________________g kg−1__________________________ | ||||||
| CS | 32.6 ± 1.0 c | 4.0 ± 0.1 c | 49.9 ± 1.6 a | 19.9 ± 1.5 bc | 4.6 ± 0.3 b | 5.6 ± 0.2 a |
| -N | 13.3 ± 1.7 d | 4.9 ± 0.3 b | 25.8 ± 1.9 c | 15.9 ± 1.7 c | 4.0 ± 0.3 bc | 5.2 ± 0.5 a |
| -P | 43.8 ± 1.5 ab | 2.5 ± 0.6 d | 35.0 ± 0.9 b | 23.6 ± 2.7 ab | 4.0 ± 0.2 bc | 5.3 ± 0.4 a |
| -K | 48.1 ± 2.3 a | 10.8 ± 0.6 a | 13.7 ± 0.4 d | 26.2 ± 1.6 a | 4.3 ± 0.2 bc | 5.6 ± 0.5 a |
| -Ca | 37.4 ± 0.3 bc | 5.6 ± 0.1 b | 38.3 ± 1.1 b | 2.6 ± 0.1 d | 6.5 ± 0.5 a | 4.3 ± 0.5 b |
| -Mg | 40.4 ± 1.0a bc | 5.1 ± 0.2 b | 38.2 ± 2.2 b | 25.4 ± 1.1 a | 1.3 ± 0.1 d | 4.3 ± 0.1 b |
| -S | 34.2 ± 1.9 bc | 5.1 ± 0.2 b | 37.7 ± 1.8 b | 26.0 ± 0.2 a | 3.7 ± 0.1 c | 1.1 ± 0.1 c |
| MSD (a) | 10.5 | 0.8 | 5.9 | 4.9 | 0.7 | 0.9 |
| CV (b) | 12.8 | 6.6 | 7.5 | 10.6 | 7.0 | 8.7 |
| Root | ||||||
| ________________________________g kg−1__________________________ | ||||||
| CS | 28.5 ± 1.7 bc | 10.3 ± 0.1 a | 23.5 ± 1.6 c | 19.1 ± 1.9 a | 5.1 ± 0.6 b | 3.9 ± 0.1 bc |
| -N | 14.8 ± 0.3 d | 4.1 ± 0.3 c | 30.6 ± 2.9 b | 7.2 ± 0.4 e | 4.2 ± 0.2 bc | 3.5 ± 0.1 cd |
| -P | 34.2 ± 2.0 ab | 2.9 ± 0.3 d | 38.4 ± 2.1 a | 9.5 ± 0.3 de | 4.3 ± 0.3 bc | 4.6 ± 0.3 a |
| -K | 22.9 ± 0.4 c | 7.0 ± 0.9 b | 10.5 ± 1.6 d | 13.3 ± 1.7 b | 5.1 ± 1.0 b | 3.2 ± 0.2 d |
| -Ca | 4.9 ± 0.1 e | 2.5 ± 0.3 d | 3.7 ± 0.2 e | 1.8 ± 0.2 f | 7.9 ± 0.4 a | 2.5 ± 0.1 e |
| -Mg | 37.9 ± 2.9 a | 7.5 ± 0.2 b | 38.5 ± 2.1 a | 12.8 ± 0.6 bc | 1.9 ± 0.2 d | 4.2 ± 0.4 ab |
| -S | 31.3 ± 1.2 b | 7.2 ± 0.4 b | 36.6 ± 2.5 ab | 10.8 ± 0.6 cd | 3.2 ± 0.2 c | 1.9 ± 0.2 f |
| MSD | 6.0 | 0.9 | 6.1 | 2.3 | 1.1 | 0.4 |
| CV | 10.5 | 7.2 | 10.3 | 9.6 | 19.9 | 6.2 |
| Treat. | N | P | K | Ca | Mg | S |
|---|---|---|---|---|---|---|
| Shoots | ||||||
| ________________________________mg per plant__________________________ | ||||||
| CS | 1072.6 ± 87.0 a | 132.1 ± 14.7 a | 1641.2 ± 146.3 a | 656.9 ± 89.1 a | 150.9 ± 7.5 a | 183.4 ± 12.2 a |
| -N | 133.7 ± 17.7 d | 48.7 ± 4.1 b | 259.3 ± 42.4 c | 159.2 ± 29.7 b | 40.1 ± 3.2 de | 52.4 ± 5.9 d |
| -P | 1000.2 ± 88.3 a | 58.7 ± 16.5 b | 760.8 ± 74.8 b | 541.6 ± 58.2 a | 91.2 ± 11.1 b | 122.3 ± 15.8 b |
| -K | 495.2 ± 66.5 c | 112.4 ± 14.7 a | 209.1 ± 144.0 c | 271.4 ± 39.2 b | 44.6 ± 4.7 d | 57.7 ± 8.2 d |
| -Ca | 389.3 ± 3.1 c | 58.6 ± 0.9 b | 397.6 ± 27.4 c | 27.0 ± 1.1 c | 67.6 ± 5.1 c | 44.3 ± 4.7 d |
| -Mg | 932.4 ± 44.1 a | 117.9 ± 10.9 a | 885.8 ± 124.6 b | 587.1 ± 57.8 a | 30.0 ± 2.9 e | 99.4 ± 7.4 c |
| -S | 712.3 ± 77.6 b | 106.8 ± 0.8 a | 785.3 ± 8.1 b | 541.6 ± 26.0 a | 77.1 ± 2.8 c | 23.0 ± 2.9 e |
| MSD (a) | 145.6 | 25.3 | 223.0 | 115.8 | 13.9 | 21.1 |
| CV (b) | 9.3 | 12.1 | 13.8 | 12.7 | 8.4 | 11.0 |
| Root | ||||||
| ________________________________mg per plant__________________________ | ||||||
| CS | 345.3 ± 41.4 a | 124.6 ± 10.9 a | 297.0 ± 21.7 b | 232.5 ± 32.6 a | 61.1 ± 5.8 a | 47.4 ± 4.9 ab |
| -N | 106.2 ± 5.1 c | 29.7 ± 1.9 cd | 219.3 ± 13.8 c | 51.6 ± 4.9 c | 30.1 ± 2.5 bc | 24.8 ± 1.1 cd |
| -P | 417.8 ± 53.9 a | 35.0 ± 4.5 c | 468.0 ± 57.4 a | 115.2 ± 6.7 b | 51.7 ± 4.8 a | 55.8 ± 7.2 a |
| -K | 222.8 ± 7.6 b | 67.8 ± 9.6 b | 102.6 ± 17.2 d | 130.1 ± 17.3 b | 49.4 ± 10.2 a | 30.9 ± 1.9 c |
| -Ca | 35.7 ± 1.3 c | 18.0 ± 1.7 d | 26.9 ± 1.5 e | 12.8 ± 1.1 d | 57.4 ± 1.7 a | 18.4 ± 0.5 d |
| -Mg | 402.3 ± 70.5 a | 78.9 ± 5.2 b | 406.8 ± 48.7 a | 135.7 ± 10.2 b | 19.7 ± 3.1 c | 44.1 ± 5.4 b |
| -S | 343.6 ± 13.2 a | 79.3 ± 3.9 b | 401.6 ± 28.1 a | 118.2 ± 6.1 b | 34.7 ± 1.8 b | 20.5 ± 2.4 d |
| MSD | 86.2 | 10.3 | 74.8 | 34.4 | 11.8 | 9.3 |
| CV | 14.0 | 14.6 | 11.9 | 13.2 | 11.8 | 11.8 |
| Whole plant | ||||||
| ________________________________mg per plant__________________________ | ||||||
| CS | 1417.9 ± 77.9 a | 256.6 ± 14.4 a | 1938.3 ± 152.8 a | 889.5 ± 76.6 a | 212.1 ± 4.8 a | 230.8 ± 8.6 a |
| -N | 239.9 ± 14.5 e | 78.3 ± 5.8 c | 478.7 ± 46.4 c | 210.8 ± 29.9 d | 70.2 ± 4.8 e | 77.3 ± 6.6 de |
| -P | 1418.0 ± 80.6 a | 93.7 ± 11.9 c | 1228.8 ± 103.7 b | 656.8 ± 55.7 b | 142.9 ± 11.9 b | 178.1 ± 14.4 b |
| -K | 718.0 ± 60.8 c | 180.3 ± 12.8 b | 311.7 ± 139.7 c | 401.6 ± 31.3 c | 94.1 ± 12.1 d | 88.6 ± 7.9 d |
| -Ca | 424.9 ± 4.4 d | 76.6 ± 2.2 c | 424.5 ± 28.8 c | 39.9 ± 0.9 e | 124.9 ± 5.7 c | 62.7 ± 4.8 ef |
| -Mg | 1334.7 ± 99.6 a | 196.9 ± 12.9 b | 1292.6 ± 165.1 b | 722.9 ± 51.5 b | 49.7 ± 4.9 f | 143.4 ± 9.9 c |
| -S | 1055.9 ± 90.8 b | 186.1 ± 3.1 b | 1186.9 ± 33.4 b | 659.9 ± 21.8 b | 111.8 ± 3.9 c | 43.5 ± 5.3 f |
| MSD | 161.8 | 23.4 | 253.2 | 102.7 | 17.6 | 20.1 |
| CV | 7.5 | 6.7 | 11.2 | 8.7 | 6.6 | 7.4 |
| Treat. | N | P | K | Ca | Mg | S |
|---|---|---|---|---|---|---|
| Absorption Efficiency | ||||||
| ________________________________mg g−1__________________________ | ||||||
| CS | 117.9 ± 7.5 a | 21.2 ± 0.4 a | 160.3 ± 6.2 a | 73.7 ± 4.4 a | 17.6 ± 1.4 a | 19.2 ± 0.9 a |
| -N | 32.6 ± 1.9 e | 10.8 ± 0.7 d | 66.4 ± 5.8 d | 28.5 ± 2.9 d | 9.7 ± 0.4 b | 10.7 ± 0.9 c |
| -P | 111.8 ± 8.7 a | 6.9 ± 0.4 e | 101.0 ± 6.3 c | 51.5 ± 5.2 c | 11.3 ± 0.9 b | 14.0 ± 0.6 b |
| -K | 77.3 ± 7.8 c | 19.2 ± 0.4 b | 26.0 ± 1.4 e | 43.0 ± 1.8 c | 9.6 ± 0.3 b | 9.4 ± 0.3 cd |
| -Ca | 58.5 ± 0.4 d | 10.5 ± 0.4 d | 57.0 ± 2.4 d | 5.4 ± 0.2 e | 17.2 ± 0.9 a | 8.6 ± 0.8 d |
| -Mg | 125.5 ± 8.5 a | 18.5 ±1.3 b | 119.1 ±12.2 b | 68.0 ±5.7 ab | 4.6 ± 0.4 c | 13.4 ± 1.0 b |
| -S | 94.0 ± 5.6 b | 16.9 ± 0.4 c | 108.2 ± 2.8 bc | 60.1 ± 1.4 b | 10.1 ± 0.2 b | 3.9 ± 0.4 e |
| MSD (a) | 15.1 | 1.5 | 14.5 | 8.4 | 1.8 | 1.7 |
| CV (b) | 7.4 | 4.5 | 6.9 | 7.7 | 6.8 | 6.7 |
| Transport efficiency | ||||||
| ________________________________%__________________________ | ||||||
| CS | 75.9 ± 0.8 b | 51.7 ± 0.9 c | 85.3 ± 1.0 b | 74.0 ± 2.6 c | 71.4 ± 2.6 a | 79.6 ± 0.8 a |
| -N | 54.7 ± 2.9 d | 61.8 ± 1.2 b | 53.9 ± 4.2 e | 74.7 ±3.2 bc | 57.1 ± 2.1 bc | 67.7 ± 2.1 b |
| -P | 69.2 ± 2.7 c | 58.4 ±5.9 bc | 61.9 ± 3.3 d | 81.4 ±1.7 ab | 62.3 ± 1.6 b | 67.2 ± 3.3 b |
| -K | 70.1 ± 3.8 bc | 63.8 ± 3.8 b | 59.7 ± 4.0 de | 68.8 ± 5.3 c | 50.4 ± 3.9 d | 66.4 ± 2.7 b |
| -Ca | 91.5 ± 0.2 a | 76.4 ± 1.7 a | 93.5 ± 0.1 a | 67.8 ± 2.6 c | 54.0 ±1.9 cd | 70.5 ± 2.2 b |
| -Mg | 69.8 ± 3.3 c | 59.7 ± 2.3 b | 69.6 ± 2.9 c | 81.0 ± 2.3 ab | 60.4 ± 3.1 b | 69.1 ± 2.6 b |
| -S | 66.6 ± 1.3 c | 57.4 ± 1.3 bc | 66.2 ± 1.4 cd | 82.0 ± 1.3 a | 68.9 ± 1.1 a | 52.8 ± 0.9 c |
| MSD | 5.8 | 6.8 | 6.6 | 6.9 | 5.8 | 5.2 |
| CV (%) | 3.5 | 4.8 | 4.1 | 3.9 | 4.1 | 3.3 |
| Use efficiency | ||||||
| ________________________________g mg−1__________________________ | ||||||
| CS | 1.5 ± 0.0 a | 8.3 ± 0.2 b | 1.1 ± 0.0 b | 2.4 ± 0.1 b | 10.0 ± 0.5 b | 9.2 ± 0.2 b |
| -N | 1.2 ± 0.0 b | 3.8 ± 0.1 e | 0.6 ± 0.0 e | 1.4 ± 0.1 cd | 4.2 ± 0.1 d | 3.8 ± 0.2 d |
| -P | 0.8 ± 0.1 cd | 13.6 ± 1.4 a | 0.9 ± 0.0 c | 1.8 ± 0.2 c | 8.3 ± 0.6 c | 6.7 ± 0.7 c |
| -K | 0.8 ± 0.0 e | 2.3 ± 0.0 f | 1.7 ± 0.1 a | 1.0 ± 0.0 d | 4.5 ± 0.0 d | 4.7 ± 0.3 d |
| -Ca | 0.7 ± 0.0 de | 4.0 ± 0.2 de | 0.7 ± 0.0 de | 7.8 ± 0.3 a | 2.5 ± 0.1 e | 5.0 ± 0.5 d |
| -Mg | 0.8 ± 0.0 cd | 5.7 ± 0.1 c | 0.8 ± 0.0 c | 1.5 ± 0.0 c | 22.8 ± 1.2 a | 7.8 ± 0.2 bc |
| -S | 0.9 ± 0.0 c | 5.4 ± 0.5 cd | 0.8 ± 0.0 cd | 1.5 ± 0.0 c | 9.0 ± 0.4 bc | 23.4 ± 1.5 a |
| MSD | 0.1 | 1.4 | 0.1 | 0.4 | 1.4 | 1.6 |
| CV (%) | 7.5 | 9.9 | 6.1 | 7.9 | 6.9 | 8.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, H.J.d.; Carmona, V.M.V.; Cavalcante, V.S.; Filho, A.B.C.; Prado, R.d.M.; Flores, R.A.; Borges, B.M.M.N.; Mauad, M. Nutritional and Visual Diagnosis in Broccoli (Brassica oleracea var. italica L.) Plants: Disorders in Physiological Activity, Nutritional Efficiency and Metabolism of Carbohydrates. Agronomy 2020, 10, 1572. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101572
Almeida HJd, Carmona VMV, Cavalcante VS, Filho ABC, Prado RdM, Flores RA, Borges BMMN, Mauad M. Nutritional and Visual Diagnosis in Broccoli (Brassica oleracea var. italica L.) Plants: Disorders in Physiological Activity, Nutritional Efficiency and Metabolism of Carbohydrates. Agronomy. 2020; 10(10):1572. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101572
Chicago/Turabian StyleAlmeida, Hilário Júnior de, Victor Manuel Vergara Carmona, Valéria Santos Cavalcante, Arthur Bernardes Cecílio Filho, Renato de Mello Prado, Rilner Alves Flores, Bernardo Melo Montes Nogueira Borges, and Munir Mauad. 2020. "Nutritional and Visual Diagnosis in Broccoli (Brassica oleracea var. italica L.) Plants: Disorders in Physiological Activity, Nutritional Efficiency and Metabolism of Carbohydrates" Agronomy 10, no. 10: 1572. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101572