CAX1a TILLING Mutations Modify the Hormonal Balance Controlling Growth and Ion Homeostasis in Brassica rapa Plants Subjected to Salinity


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Experimental Design, Treatments, and Plant Sampling
2.3. Analysis of Na+, Ca2+, and K+ Concentrations
2.4. Hormone Extraction and Analysis
2.5. Statistical Analysis
3. Results
3.1. Plant Biomass and Cation Concentration
3.2. Phytohormone Concentrations
3.3. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wu, H. Plant salt tolerance and Na+ sensing and transport. Crop J. 2018, 6, 215–225. [Google Scholar] [CrossRef]
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.; Hernandez, J. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Pavlović, I.; Pěnčík, A.; Novák, O.; Vujčić, V.; Radić Brkanac, S.; Lepeduš, H.; Strnad, M.; Salopek-Sondi, B. Short-term salt stress in Brassica rapa seedlings causes alterations in auxin metabolism. Plant Physiol. Biochem. 2018, 125, 74–84. [Google Scholar] [CrossRef]
- Köster, P.; Wallrad, L.; Edel, K.H.; Faisal, M.; Alatar, A.A.; Kudla, J. The battle of two ions: Ca2+ signalling against Na+ stress. Plant Biol. 2018, 21, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Shoresh, M.; Spivak, M.; Bernstein, N. Involvement of calcium-mediated effects on ROS metabolism in the regulation of growth improvement under salinity. Free Radic. Biol. Med. 2011, 51, 1221–1234. [Google Scholar] [CrossRef] [PubMed]
- Kravchik, M.; Bernstein, N. Effects of salinity on the transcriptome of growing maize leaf cells point at cell-age specificity in the involvement of the antioxidative response in cell growth restriction. BMC Genom. 2013, 14, 24. [Google Scholar] [CrossRef] [Green Version]
- Ku, Y.-S.; Sintaha, M.; Cheung, M.-Y.; Lam, H.-M. Plant hormone signaling crosstalks between biotic and abiotic stress responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Khan, M.I.R. A new perspective of phytohormones in salinity tolerance: Regulation of proline metabolism. Environ. Exp. Bot. 2014, 100, 34–42. [Google Scholar] [CrossRef]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M.; Khan, M.M.A. Calcium chloride and gibberellic acid protect linseed (Linum usitatissimum L.) from NaCl stress by inducing antioxidative defence system and osmoprotectant accumulation. Acta Physiol. Plant. 2010, 32, 121–132. [Google Scholar] [CrossRef]
- Dar, T.A.; Uddin, M.; Khan, M.M.A.; Hakeem, K.R.; Jaleel, H. Jasmonates counter plant stress: A Review. Environ. Exp. Bot. 2015, 115, 49–57. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Manishankar, P.; Wang, N.; Köster, P.; Alatar, A.A.; Kudla, J. Calcium signaling during salt stress and in the regulation of ion homeostasis. J. Exp. Bot. 2018, 69, 4215–4226. [Google Scholar] [CrossRef] [Green Version]
- Yousuf, P.Y.; Ahmad, A.; Hemant, M.; Ganie, A.H.; Aref, I.M.; Iqbal, M. Potassium and calcium application ameliorates growth and oxidative homeostasis in salt-stressed indian mustard (Brassica Juncea) plants. Pak. J. Bot. 2015, 47, 1629–1639. [Google Scholar]
- Latef, A.; Hamed, A.A. Ameliorative effect of calcium chloride on growth, antioxidant enzymes, protein patterns and some metabolic activities of canola (Brassica napus L.) under seawater stress. J. Plant Nutr. 2011, 34, 1303–1320. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M.; Jamil, A.; Ur-Rehman, S. Does seed priming induce changes in the levels of some endogenous plant hormones in hexaploid wheat plants under salt stress? J. Integr. Plant Biol. 2006, 48, 181–189. [Google Scholar] [CrossRef]
- Pittman, J.K.; Hirschi, K.D. CAX-ing a wide net: Cation/H+ transporters in metal remediation and abiotic stress signaling. Plant Biol. 2016, 18, 741–749. [Google Scholar] [CrossRef]
- Pokotylo, I.V.; Kretinin, S.V.; Kravets, V.S. Role of phospholipase D in metabolic reactions of transgenic tobacco cax1 cells under the influence of salt stress. Cytol. Genet. 2012, 46, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Han, N.; Shao, Q.; Bao, H.; Wang, B. Cloning and characterization of a Ca2+/H+ antiporter from halophyte Suaeda salsa L. Plant Mol. Biol. Rep. 2011, 29, 449–457. [Google Scholar] [CrossRef]
- Till, B.J.; Reynolds, S.H.; Greene, E.A.; Codomo, C.A.; Enns, L.C.; Johnson, J.E.; Burtner, C.; Odden, A.R.; Young, K.; Taylor, N.E.; et al. Large-scale discovery of induced point mutations with high-throughput TILLING. Genome Res. 2003, 13, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Lochlainn, S.Ó.; Amoah, S.; Graham, N.S.; Alamer, K.; Rios, J.J.; Kurup, S.; Stoute, A.; Hammond, J.P.; Østergaard, L.; King, G.J.; et al. High Resolution Melt (HRM) analysis is an efficient tool to genotype EMS mutants in complex crop genomes. Plant Methods 2011, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, N.S.; Hammond, J.P.; Lysenko, A.; Mayes, S.; Lochlainn, S.O.; Blasco, B.; Bowen, H.C.; Rawlings, C.J.; Rios, J.J.; Welham, S.; et al. Genetical and comparative genomics of Brassica under altered Ca supply identifies Arabidopsis Ca-transporter orthologs. Plant Cell 2014, 26, 2818–2830. [Google Scholar] [CrossRef] [Green Version]
- Navarro-León, E.; Ruiz, J.M.; Albacete, A.; Blasco, B. Effect of CAX1a TILLING mutations and calcium concentration on some primary metabolism processes in Brassica rapa plants. J. Plant Physiol. 2019, 237, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Wolf, B. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Albacete, A.; Ghanem, M.E.; Martinez-Andujar, C.; Acosta, M.; Sanchez-Bravo, J.; Martinez, V.; Lutts, S.; Dodd, I.C.; Perez-Alfocea, F. Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2008, 59, 4119–4131. [Google Scholar] [CrossRef]
- Wan, H.; Chen, L.; Guo, J.; Li, Q.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Fu, T.; Shen, J. Genome-wide association study reveals the genetic architecture underlying salt tolerance-related traits in rapeseed (Brassica napus L.). Front. Plant Sci. 2017, 8, 593. [Google Scholar] [CrossRef]
- Navarro-León, E.; López-Moreno, F.J.; de la Torre-González, A.; Ruiz, J.M.; Esposito, S.; Blasco, B. Study of salt-stress tolerance and defensive mechanisms in Brassica rapa CAX1a TILLING mutants. Environ. Exp. Bot. 2020, 175, 104061. [Google Scholar] [CrossRef]
- Mei, H.; Zhao, J.; Pittman, J.K.; Lachmansingh, J.; Park, S.; Hirschi, K.D. In planta regulation of the Arabidopsis Ca2+/H+ antiporter CAX1. J. Exp. Bot. 2007, 58, 3419–3427. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Albacete, A.; Martínez-Andújar, C.; Acosta, M.; Romero-Aranda, R.; Dodd, I.C.; Lutts, S.; Pérez-Alfocea, F. Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 2008, 59, 3039–3050. [Google Scholar] [CrossRef] [Green Version]
- de la Torre-González, A.; Navarro-León, E.; Albacete, A.; Blasco, B.; Ruiz, J.M. Study of phytohormone profile and oxidative metabolism as key process to identification of salinity response in tomato commercial genotypes. J. Plant Physiol. 2017, 216, 164–173. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.; Huang, B. Cytokinin-mitigation of salt-induced leaf senescence in perennial ryegrass involving the activation of antioxidant systems and ionic balance. Environ. Exp. Bot. 2016, 125, 1–11. [Google Scholar] [CrossRef]
- Pavlů, J.; Novák, J.; Koukalová, V.; Luklová, M.; Brzobohatý, B.; Černý, M. Cytokinin at the crossroads of abiotic stress signalling pathways. Int. J. Mol. Sci. 2018, 19, 2450. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; He, J.; Chen, J.; Yang, S.; Zha, D. Alleviation of exogenous 6-benzyladenine on two genotypes of eggplant (Solanum melongena Mill.) growth under salt stress. Protoplasma 2014, 251, 169–176. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Albacete, A.; Smigocki, A.C.; Frébort, I.; Pospíšilová, H.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Lutts, S.; Dodd, I.C.; et al. Root-synthesized cytokinins improve shoot growth and fruit yield in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2011, 62, 125–140. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, W.-Y.; Yun, D.-J. A new insight of salt stress signaling in plant. Mol. Cells 2016, 39, 447–459. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Zörb, C.; Geilfus, C.-M.; Dietz, K.-J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef]
- Zörb, C.; Geilfus, C.-M.; Mühling, K.H.; Ludwig-Müller, J. The influence of salt stress on ABA and auxin concentrations in two maize cultivars differing in salt resistance. J. Plant Physiol. 2013, 170, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Amjad, M.; Akhtar, J.; Anwar-ul-Haq, M.; Yang, A.; Akhtar, S.S.; Jacobsen, S.-E. Integrating role of ethylene and ABA in tomato plants adaptation to salt stress. Sci. Hortic. (Amsterdam) 2014, 172, 109–116. [Google Scholar] [CrossRef]
- Trivellini, A.; Lucchesini, M.; Ferrante, A.; Carmassi, G.; Scatena, G.; Vernieri, P.; Mensuali-Sodi, A. Survive or die? A molecular insight into salt-dependant signaling network. Environ. Exp. Bot. 2016, 132, 140–153. [Google Scholar] [CrossRef]
- Arif, Y.; Sami, F.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salicylic acid in relation to other phytohormones in plant: A study towards physiology and signal transduction under challenging environment. Environ. Exp. Bot. 2020, 175, 104040. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotoxicol. Environ. Saf. 2018, 147, 1010–1016. [Google Scholar] [CrossRef]




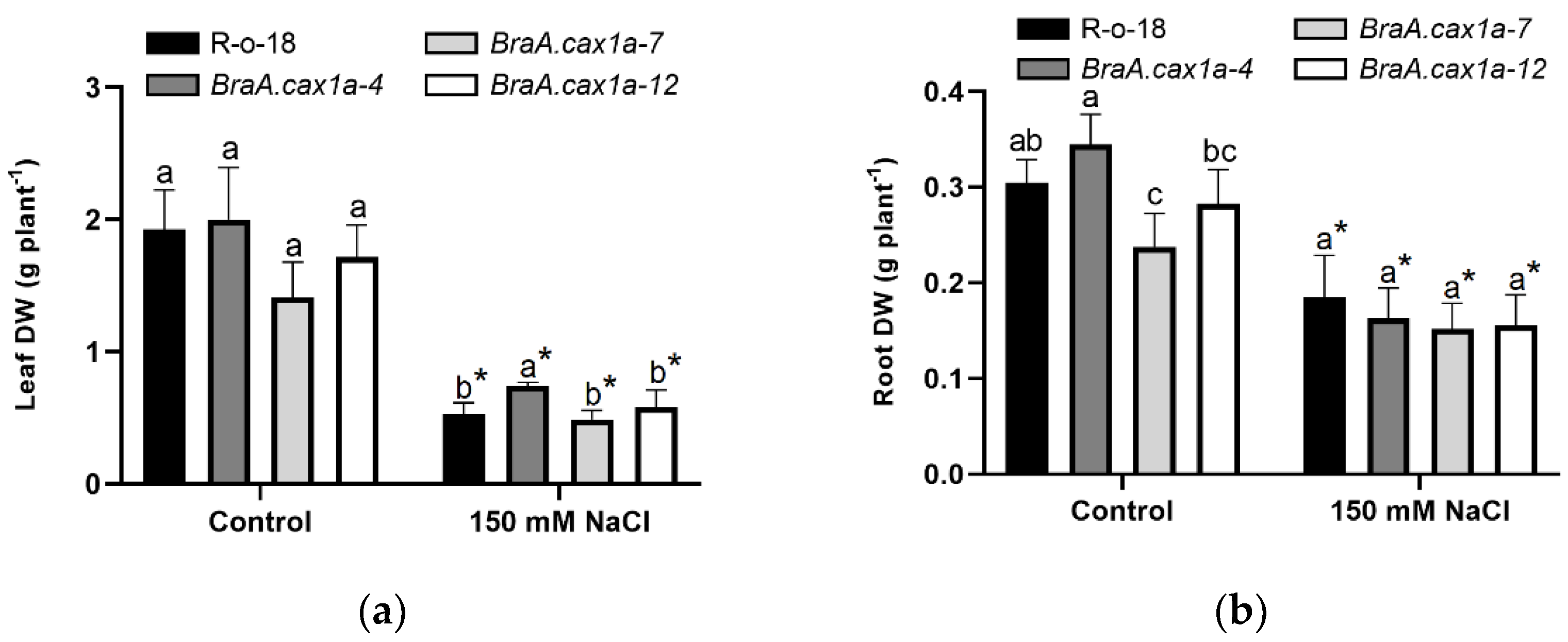{kind=link}
{kind=link}
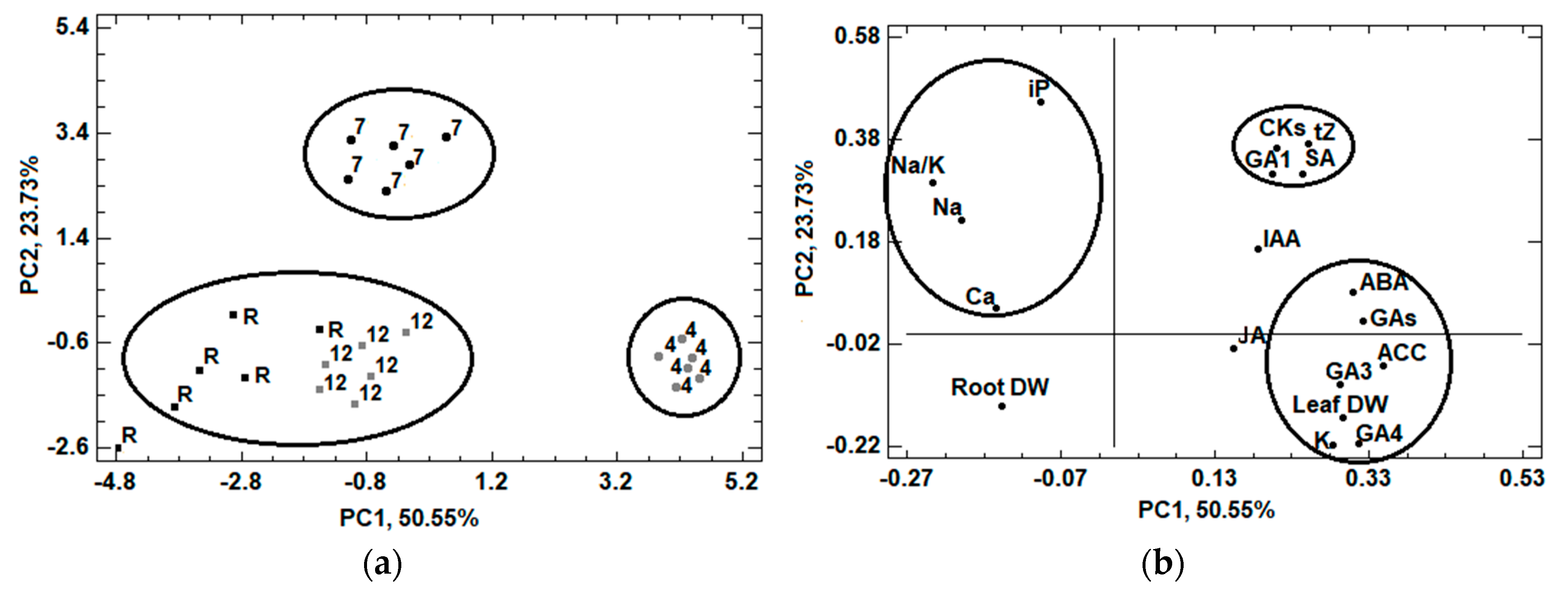{kind=link}
| Ca2+ | Na+ | K+ | Na+/K+ | ||
|---|---|---|---|---|---|
| Control | R-o-18 | 15.09 b | 3.29 a | 39.24 ab | 0.08 a |
| BraA.cax1a-4 | 17.61 a | 3.64 a | 38.81 ab | 0.09 a | |
| BraA.cax1a-7 | 17.85 a | 3.23 a | 36.69 b | 0.09 a | |
| BraA.cax1a-12 | 18.51 a | 3.76 a | 41.14 a | 0.09 a | |
| p-value | * | NS | * | NS | |
| LSD0.05 | 2.22 | 1.05 | 4.11 | 0.02 | |
| 150 mM NaCl | R-o-18 | 9.50 a | 36.74 a | 22.17 b | 1.65 ab |
| BraA.cax1a-4 | 8.54 a | 26.65 b | 28.15 a | 0.96 c | |
| BraA.cax1a-7 | 9.57 a | 41.07 a | 22.15 b | 1.85 a | |
| BraA.cax1a-12 | 10.19 a | 40.68 a | 25.85 a | 1.57 b | |
| p-value | NS | * | ** | *** | |
| LSD0.05 | 2.22 | 9.81 | 2.55 | 0.26 | |
| Analysis of variance | |||||
| Salinity (S) | *** | *** | *** | *** | |
| Mutation (M) | * | * | ** | *** | |
| S × M | NS | * | * | *** | |
| LSD0.05 | 1.44 | 4.54 | 2.22 | 0.12 | |
| IAA | GA1 | GA3 | GA4 | iP | tZ | ABA | ACC | SA | JA | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | R-o-18 | 2.40 bc | 0.05 b | nd | 0.02 b | 6.83 a | 180.47 a | 6.43 a | 489.79 c | 282.53 a | 131.42 b |
| BraA.cax1a-4 | 3.43 b | 0.10 a | 0.08 | 0.05 a | 4.55 b | 129.43 ab | 4.38 ab | 848.95 a | 283.45 a | 68.98 c | |
| BraA.cax1a-7 | 1.62 c | 0.07 b | nd | 0.01 b | 2.94 b | 119.26 b | 4.13 b | 343.59 d | 317.96 a | 102.67 bc | |
| BraA.cax1a-12 | 5.42 a | 0.06 b | 0.05 | 0.04 a | 8.54 a | 176.00 a | 6.49 a | 623.31 b | 306.47 a | 190.93 a | |
| p-value | ** | ** | * | *** | * | * | *** | NS | ** | ||
| LSD0.05 | 1.55 | 0.02 | 0.03 | 1.29 | 56.09 | 2.12 | 59.33 | 165.18 | 54.96 | ||
| 150 mM NaCl | R-o-18 | 2.03 b | 0.08 c | 0.03 c | 0.04 b | 5.10 b | 231.93 b | 13.93 b | 515.55 c | 611.82 c | 125.07 c |
| BraA.cax1a-4 | 3.14 a | 0.16 ab | 0.18 a | 0.11 a | 2.91 c | 355.02 a | 21.86 a | 1234.98 a | 942.16 ab | 201.57 ab | |
| BraA.cax1a-7 | 2.95 ab | 0.18 a | 0.05 bc | 0.03 b | 7.82 a | 382.54 a | 16.86 ab | 695.43 b | 1035.42 a | 168.46 bc | |
| BraA.cax1a-12 | 2.79 ab | 0.09 bc | 0.07 b | 0.04 b | 2.94 c | 255.38 b | 15.01 b | 722.12 b | 795.16 bc | 224.16 a | |
| p-value | * | * | *** | ** | ** | ** | * | *** | ** | ** | |
| LSD0.05 | 1.04 | 0.07 | 0.04 | 0.0 4 | 2.05 | 63.88 | 5.62 | 164.17 | 206.85 | 50.51 | |
| Analysis of variance | |||||||||||
| Salinity (S) | NS | *** | ** | * | *** | *** | *** | *** | *** | ||
| Mutation (M) | *** | *** | *** | ** | NS | NS | *** | ** | *** | ||
| S × M | ** | NS | NS | *** | *** | ** | *** | * | ** | ||
| LSD0.05 | 0.86 | 0.03 | 0.02 | 1.11 | 39.07 | 2.76 | 80.24 | 121.67 | 34.31 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-León, E.; López-Moreno, F.J.; Atero-Calvo, S.; Albacete, A.; Ruiz, J.M.; Blasco, B. CAX1a TILLING Mutations Modify the Hormonal Balance Controlling Growth and Ion Homeostasis in Brassica rapa Plants Subjected to Salinity. Agronomy 2020, 10, 1699. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10111699
Navarro-León E, López-Moreno FJ, Atero-Calvo S, Albacete A, Ruiz JM, Blasco B. CAX1a TILLING Mutations Modify the Hormonal Balance Controlling Growth and Ion Homeostasis in Brassica rapa Plants Subjected to Salinity. Agronomy. 2020; 10(11):1699. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10111699
Chicago/Turabian StyleNavarro-León, Eloy, Francisco Javier López-Moreno, Santiago Atero-Calvo, Alfonso Albacete, Juan Manuel Ruiz, and Begoña Blasco. 2020. "CAX1a TILLING Mutations Modify the Hormonal Balance Controlling Growth and Ion Homeostasis in Brassica rapa Plants Subjected to Salinity" Agronomy 10, no. 11: 1699. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10111699