The Cultivation Techniques and Quality Characteristics of a New Germplasm of Vitis adenoclada Hand.-Mazz Grape

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Experimental Site and Plant Material
2.2. Description of the Variety’s Characteristics
2.3. A Brief Technique for the Cultivation and Management of GH3 in Karst Rocky Desertification Mountains
2.3.1. Selection of Land, Preparation of Land, Varieties, and Planting
2.3.2. Training System and Pruning
2.3.3. Fertilizer Management
2.3.4. Pest Control
2.4. Berry Sampling and Physical Chemical Index Analysis
2.5. Widely Targeted Metabolomic Analysis
2.5.1. Sample Preparation and Extraction
2.5.2. UPLC Conditions
2.5.3. ESI–QTRAP–MS/MS
2.6. Statistical Analysis
3. Results
3.1. The Variety’s Characteristics
3.1.1. Botanical Characteristics
3.1.2. Phenological Period
3.1.3. Stress Resistance
3.2. Grape Berry Development
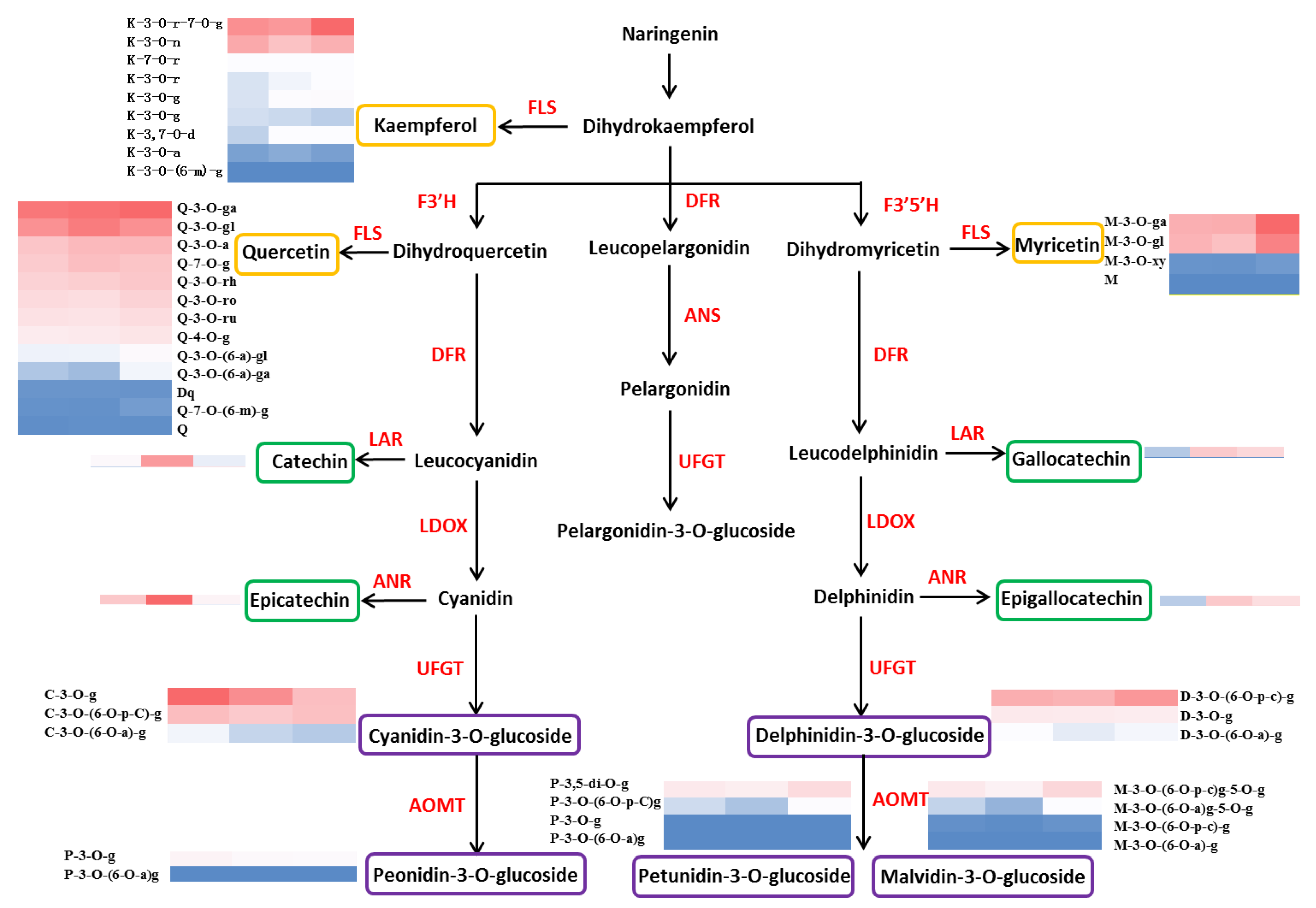
3.3. Metabolism and Component Characteristics of Sugars, Organic Acids, and Phenolic Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meng, J.X.; Zhang, G.H.; Fan, X.C.; Zhang, Y.; Zhang, X.L.; Liu, C.H.; Jiang, J.F. Studies on distribution of wild grapes in China. J. Plant. Genet. Resour. 2020, 21, 1539–1548. [Google Scholar]
- Liu, K.Y. Identification, Evaluation and Genetic Diversity of Vitis adenoclada Hand.–Mazz. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2012. [Google Scholar]
- Liu, K.Y.; Fang, F.; Shi, X.H.; Yang, G.S.; Zhong, X.H.; Ni, J.J.; Xu, F.; Bai, M.; Nie, S.Q.; Chen, X.Y. Resistance of Vitis adenoclada Hand.–Mazz. and Vitis davidii Foëx to Sphaceloma ampelinum and Plasmopara viticola. J. Hunan Agric. Univ. 2013, 39, 46–51. [Google Scholar] [CrossRef]
- Ren, F.H.; Zhong, W.M.; Zhang, W.E.; Hu, J.M.; Pan, X.J. Drought resistance evaluation of wild Vitis seedlings in Karst region. J. Northwest A F Univ. 2019, 47, 98–106. [Google Scholar] [CrossRef]
- Pan, X.J.; Zhang, W.E.; Yang, X.Y.; Li, S.Y. Drought-resistance mechanism of four kinds of wild Vitis seedlings in Karst regions of Guizhou province. Acta Bot. Boreal. Occident. Sin. 2010, 30, 955–961. [Google Scholar]
- Fang, F. Resistance Identification and Evaluation of Sphaceloma Ampelinum de Bary and Grape Downy Mildew to V. adenoclada Hand.–Mazz. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2010. [Google Scholar]
- Wang, M.; Su, C.C.; Shi, X.H.; Yang, G.S.; Zhong, X.H.; Liu, K.Y.; Xu, F.; Jin, Y.; Bai, M. Preliminary study on photosynthetic characteristics of different types of grapes. Sino-Overseasgrapevine Wine 2017, 4, 25–28. [Google Scholar]
- Li, H.X.; Chen, H.; Wang, Z.; Zhong, X.H.; Wang, X.R.; Liu, K.Y. A Survey Study on the New Types of Agronomic Characteristic of Vitis adenoclona Hand.–Mazz. Northwest. Hortic. 2018, 12, 63–67. [Google Scholar]
- Keller, M. The Science of Grapevines: Anatomy and Physiology II Developmental Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2010; pp. 169–225. [Google Scholar]
- Coombe, B.G. Adoption of a system for identifying grapevine growth stages. Aust. J. Grape. Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Wei, Z.W.; Liu, X.H.; Huang, Y.; Lu, J.; Zhang, Y.L. Volatile aroma compounds in wines from Chinese wild/hybrid species. J. Food Biochem. 2018, 43, e12684. [Google Scholar] [CrossRef]
- Liu, C.H.; Jiang, J.F.; Fan, X.C.; Zhang, Y. The utilization of Chinese wild grape species in production and breeding. J. Plant Genet. Resour. 2014, 15, 720–727. [Google Scholar]
- Luo, S.; He, P.; Zheng, X. Inheritance of RAPD markers in an interspecific F1 hybrid of grape between Vitis quinquangularis and V. vinifera. Sci. Hortic. 2002, 93, 19–28. [Google Scholar] [CrossRef]
- Cheng, G.; Zhou, S.H.; Wen, R.D.; Xie, T.L.; Huang, Y.; Yang, Y.; Guan, J.X.; Xie, L.J.; Zhang, J. Anthocyanin characteristics of wines in Vitis germplasms cultivated in southern China. Food Sci. Technol. 2018, 38, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Wu, P.; Long, F.; Lin, Q.; He, H.W.; Zou, Y.; Pan, Y.J.; Li, X.G.; Lv, C.A.; Han, X.H. Breeding and cultivation techniques of a new wild Vitis quinquangularis Yeniang No. 3. South China Fruits 2020, 4, 167–168, 171. [Google Scholar]
- Jiang, Y. The Quality Evaluation and Main Components of Chinese Wild Grape Berry. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2012. [Google Scholar]
- Claudia, M.E.; Alex, D.G.; Alicia, S.; José, C.; Alonso, E.; Claudio, M.; Alejandro, M.; Ariel, O.; Patricio, H. Identification of SNPs and InDels associated with berry size in table grapes integrating genetic and transcriptomic approaches. BMC Plant Biol. 2020, 20, 365. [Google Scholar] [CrossRef]
- Shiraishi, M.; Shinomiya, R.; Chijiwa, H. Preliminary genetic analysis of sucrose accumulation in berries of table grapes. Sci. Hortic. 2012, 137, 107–113. [Google Scholar] [CrossRef]
- Jocimar, R.D.S.M.; Valeria, D.S.P.C.; Lima, M.D.S.; Pereira, G.E.; Venturini Filho, W.G.; Moura, M.F.; Tecchio, M.A. Grape juices produced from new hybrid varieties grown on brazilian rootstocks-bioactive compounds, organic acids and antioxidant capacity. Food Chem. 2019, 289, 714–722. [Google Scholar] [CrossRef]
- Yang, B.H.; Yao, H.; Zhang, J.X.; Li, Y.Q.; Ju, Y.L.; Zhao, X.F.; Sun, X.Y.; Fang, Y.L. Effect of regulated deficit irrigation on the content of soluble sugars, organic acids and endogenous hormones in Cabernet Sauvignon in the Ningxia region of China. Food Chem. 2020, 312, 126020. [Google Scholar] [CrossRef]
- Silva, F.L.N.; Schmidt, E.M.; Messias, C.L.; Eberlin, M.N.; Helena, A.C.; Sawaya, F. Quantitation of organic acids in wine and grapes by direct infusion electro-spray ionization mass spectrometry. Anal. Methods 2015, 7, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.Z.; Cheng, G.; Li, Q.; He, Y.N.; Wang, Y.; Lan, Y.B.; Li, S.Y.; Zhu, Y.R.; Song, W.F.; Zhang, X.; et al. Light-induced variation in phenolic compounds in cabernet sauvignon grapes (Vitis vinifera L.) involves extensive transcriptome reprogramming of biosynthetic enzymes, transcription factors, and phytohormonal regulators. Front. Plant Sci. 2017, 8, 547. [Google Scholar] [CrossRef] [Green Version]
- Koyama, R.; de Assis, A.M.; Yamamoto, L.Y.; Borges, W.F.S.; de Sá Borges, R.R.; Prudêncio, S.H.; Roberto, S.R. Exogenous abscisic acid increases the anthocyanin concentration of berry and juice from ‘Isabel’ Grapes (Vitis labrusca L.). HortScience 2014, 49, 460–464. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.S.; Silani, I.S.V.; Toaldo, I.M.; Corrêa, L.C.; Biasoto, A.C.T.; Pereira, G.E.; Bordignon-Luiz, M.T.; Ninow, J.L. Phenolic compounds, organic acids and antioxidant activity of grape juices produced from new Brazilian varieties planted in the Northeast Region of Brazil. Food Chem. 2014, 161, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Zhou, S.H.; Zhang, J.; Huang, X.Y.; Bai, X.J.; Xie, T.L.; Guo, R.R.; Liu, J.B.; Yu, H.; Xie, L.J. Comparison of transcriptional expression patterns of phenols and carotenoids in ‘kyoho’ grapes under a two-crop-a-year cultivation system. PLoS ONE 2019, 14, e0210322. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.Y.; Wang, L.; Belwal, T.; Zhang, X.C.; Lu, H.Y.; Chen, C.K.; Li, L. Exogenous melatonin and abscisic acid expedite the flavonoids biosynthesis in grape berry of Vitis vinifera cv. Kyoho. Molecules 2020, 25, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Averilla, J.N.; Oh, J.; Wu, Z.X.; Liu, K.H.; Jang, C.H.; Kim, H.J.; Kim, J.S. Improved extraction of resveratrol and antioxidants from grape peel using heat and enzymatic treatments. J. Sci. Food Agric. 2019, 99, 4043–4053. [Google Scholar] [CrossRef] [PubMed]
- Falginella, L.; Di Gaspero, G.; Castellarin, S.D. Expression of flavonoid genes in the red grape berry of ‘Alicante Bouschet’ varies with the histological distribution of anthocyanins and their chemical composition. Planta 2016, 2036, 1037–1051. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Indicator | Value |
|---|---|---|
| Leaf | Leaf length | 16.61 ± 1.43 |
| Leaf width | 12.82 ± 0.99 | |
| Petiole length | 8.28 ± 1.26 | |
| Cluster | Cluster length | 13.70 ± 1.60 |
| Cluster width | 8.04 ± 1.63 | |
| Cluster weight | 132.30 ± 19.48 | |
| Berry | Longitudinal diameter | 1.59 ± 0.03 |
| Transverse diameter | 1.49 ± 0.02 | |
| Shape index | 1.07 ± 0.01 | |
| Berry weight | 2.33 ± 0.06 | |
| Number of seeds | 4.16 ± 0.15 | |
| Skin weight | 0.39 ± 0.02 | |
| Skin to berry ratio | 16.89 ± 0.73 |
| Parameters | Developmental Stages | ||||
|---|---|---|---|---|---|
| E-L 32 | E-L 33 | E-L 35 | E-L 37 | E-L 38 | |
| Berry fresh weight (g) | 1.33 ± 0.03 d | 1.41 ± 0.03 d | 1.60 ± 0.05 c | 1.79 ± 0.31 b | 2.33 ± 0.06 a |
| Total soluble solids (°Brix) | 4.67 ± 0.15 e | 7.37 ± 0.15 d | 13.80 ± 0.10 c | 16.80 ± 0.10 b | 21.10 ± 0.10 a |
| pH | 2.11 ± 0.07 d | 2.14 ± 0.03 d | 2.72 ± 0.03 c | 2.89 ± 0.06 b | 3.16 ± 0.06 a |
| Titratable acidity (g/L) | 53.40 ± 0.53 a | 51.19 ± 0.75 b | 17.71 ± 0.84 c | 12.43 ± 0.24 d | 5.06 ± 0.07 e |
| L* | 42.36 ± 0.23 a | 41.26 ± 0.67 a | 35.87 ± 0.95 b | 26.66 ± 0.16 c | 23.75 ± 1.12 d |
| a* | –2.83 ± 0.15 e | –2.67 ± 0.15 d | –0.43 ± 0.67 c | 1.79 ± 0.19 b | 1.91 ± 0.18 a |
| b* | 19.81 ± 0.45 a | 16.90 ± 0.30 b | 12.86 ± 1.02 c | 2.81 ± 0.14 d | 2.82 ± 0.13 d |
| Compounds | Proportion (%) | Kegg Map | |
|---|---|---|---|
| Sugars | Glucose | 49.73 ± 0.74 | KO00010, KO00500, KO00052, KO00520 |
| Fructose | 48.10 ± 0.14 | KO00051 | |
| Sucrose | 2.16 ± 0.02 | KO00500, KO00052, KO00520 | |
| Organic Acids | Citric acid | 49.40 ± 4.41 | KO00020, KO00630, KO01200, KO01210, KO01230 |
| Succinic acid | 29.43 ± 7.41 | KO00020, KO00360, KO00630, KO01200 | |
| Malic acid | 12.66 ± 2.41 | KO00020, KO00630, KO01200 | |
| Tartaric acid | 7.18 ± 4.02 | KO00630 | |
| Fumaric acid | 1.29 ± 0.26 | KO00020, KO01200 | |
| Lactic acid | 0.04 ± 0.01 | KO00620 | |
| Phenolic Compounds | Number | Proportion (%) | Kegg Map | Proportion of Total Phenolics (%) | |
|---|---|---|---|---|---|
| Phenolic acids | Benzoic acids | 12 | 51.08 ± 2.44 | KO00940 | 2.40 ± 0.03 |
| Benzeneacetic acids | 2 | 4.50 ± 0.23 | KO00940 | ||
| Cinnamic acids | 8 | 44.42 ± 2.54 | KO00940 | ||
| Stilbenes | Resveratrol | 1 | 10.13 ± 0.83 | KO00945 | 13.55 ± 1.87 |
| Piceid | 1 | 89.87 ± 0.83 | KO00945 | ||
| Flavonoids | Flavonols | 26 | 73.26 ± 2.42 | KO00941, KO00944 | 84.06 ± 1.90 |
| Flavan-3-ols | 6 | 4.98 ± 0.89 | KO00941 | ||
| Anthocyanins | 16 | 21.77 ± 2.99 | KO00941 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, D.-D.; Cheng, G.; Li, H.-Y.; Zhou, S.-H.; Yao, N.; Zhang, J.; Xie, L.-J. The Cultivation Techniques and Quality Characteristics of a New Germplasm of Vitis adenoclada Hand.-Mazz Grape. Agronomy 2020, 10, 1851. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10121851
Wu D-D, Cheng G, Li H-Y, Zhou S-H, Yao N, Zhang J, Xie L-J. The Cultivation Techniques and Quality Characteristics of a New Germplasm of Vitis adenoclada Hand.-Mazz Grape. Agronomy. 2020; 10(12):1851. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10121851
Chicago/Turabian StyleWu, Dai-Dong, Guo Cheng, Hong-Yan Li, Si-Hong Zhou, Ning Yao, Jin Zhang, and Lin-Jun Xie. 2020. "The Cultivation Techniques and Quality Characteristics of a New Germplasm of Vitis adenoclada Hand.-Mazz Grape" Agronomy 10, no. 12: 1851. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10121851