Fungal Diseases in Two North-West Spain Vineyards: Relationship with Meteorological Conditions and Predictive Aerobiological Model

 ,
,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
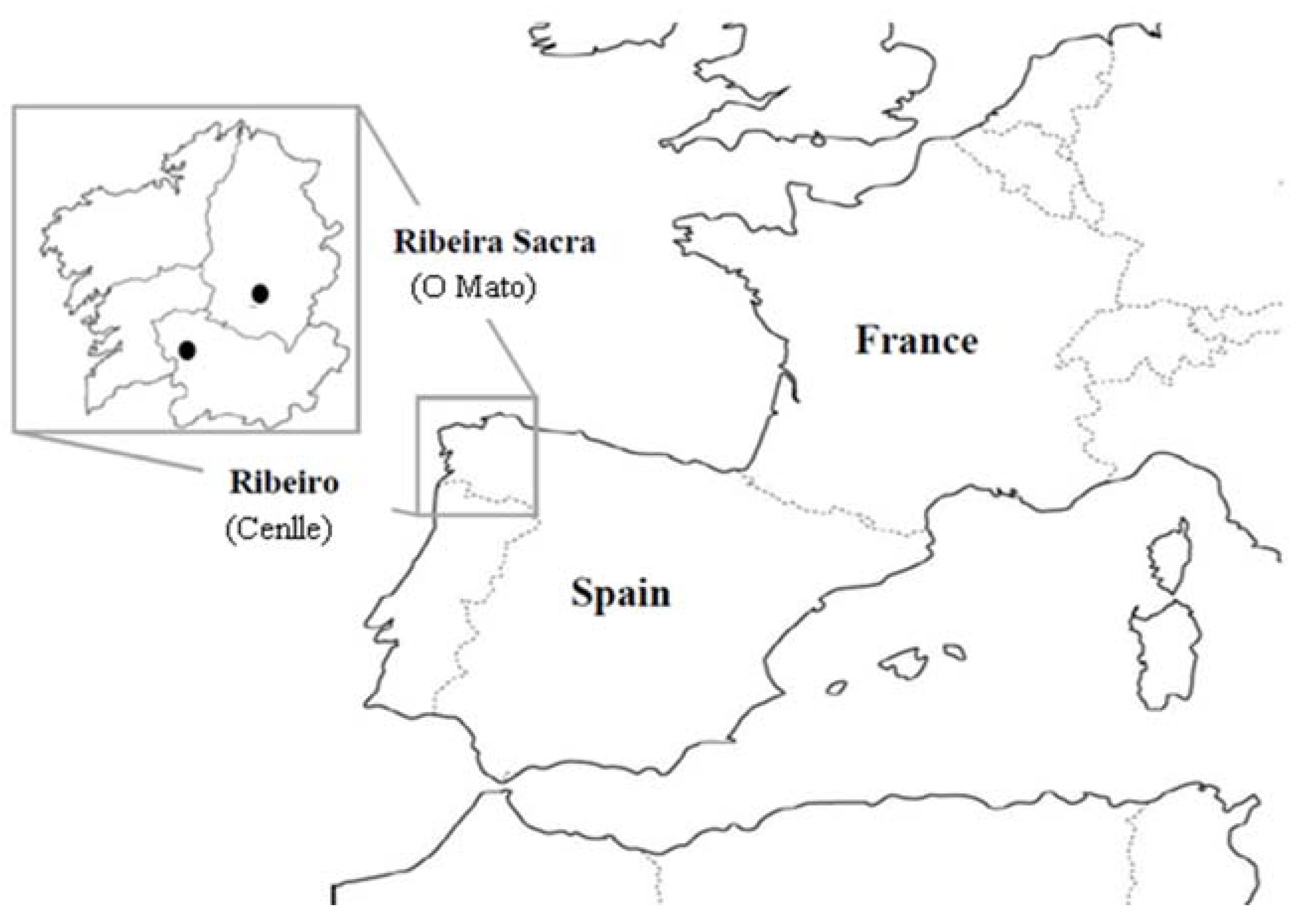
2.1. Location and Climatic Characteristics of the Study Area
2.2. Fieldwork and Laboratory Analysis
2.3. Statistical Analysis
3. Results
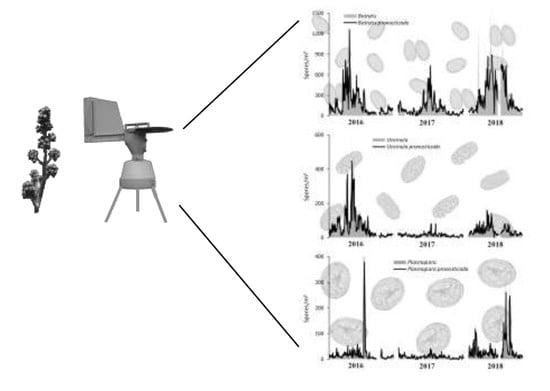
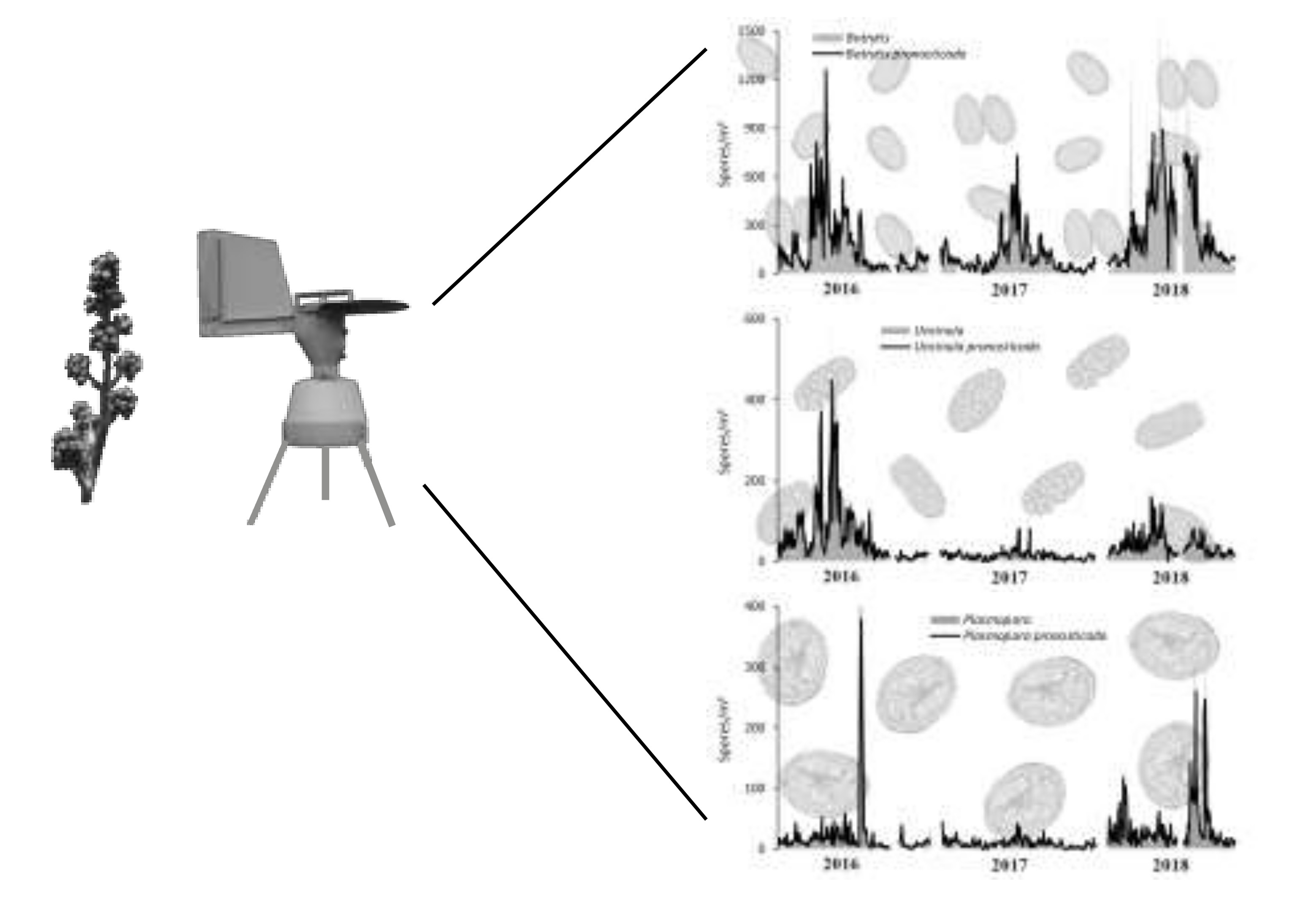
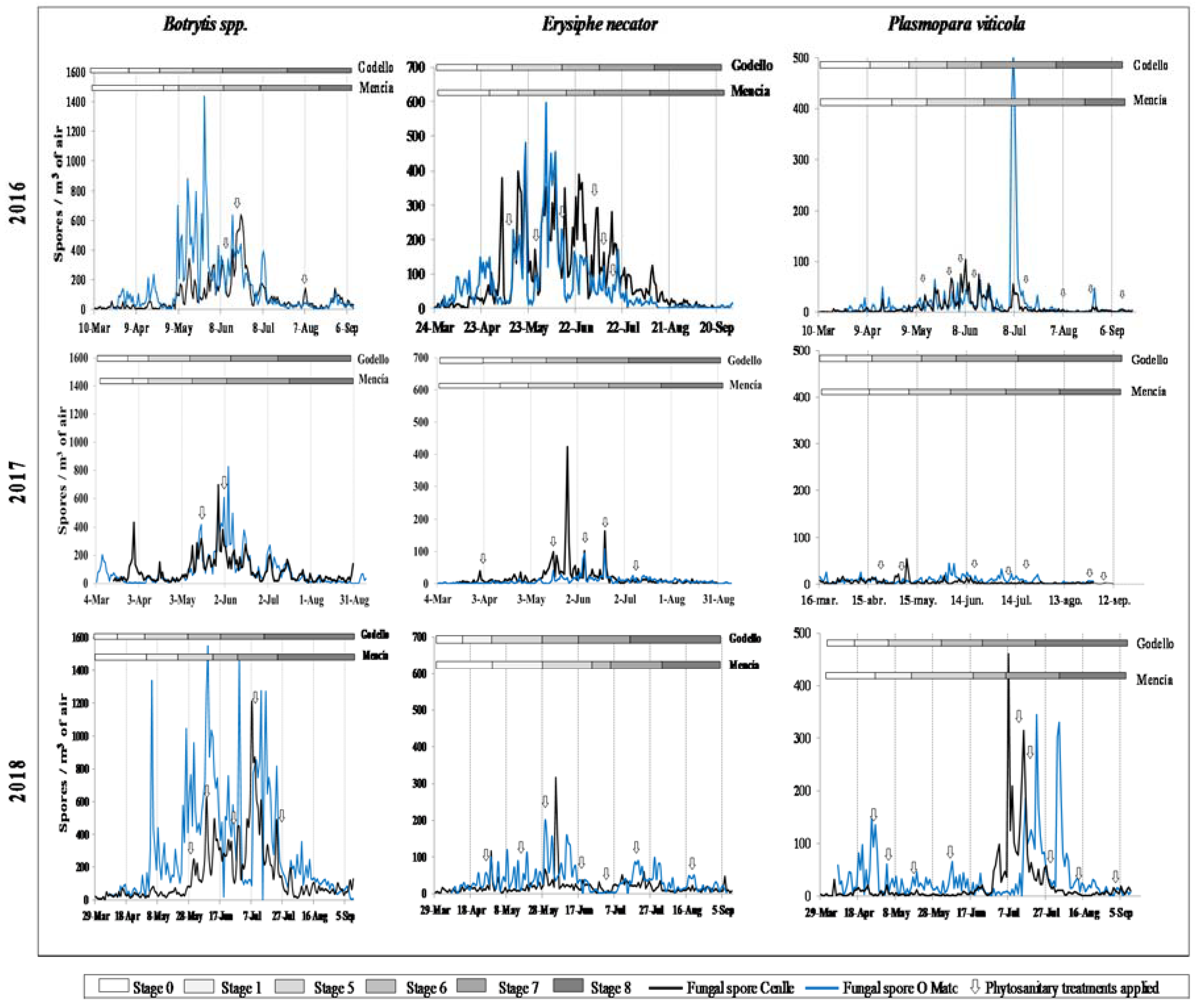
3.1. Total Spore Concentrations and Spatial Distribution
3.2. Analysis of Meteorological Parameters
3.3. Statistical Results
3.4. Predictive Models
4. Discussion
4.1. Relationship with Phenological Stage and Meteorological Influence
4.2. Predictive Models
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ciliberti, N.; Fermaud, M.; Roudet, J.; Languasco, L.; Rossi, V. Environmental effects on the production of Botrytis cinerea conidia on different media, grape bunch trash, and mature berries. Aust. J. Grape Wine Res. 2016, 22, 262–270. [Google Scholar] [CrossRef]
- Bois, B.; Zito, S.; Calonnec, A. Climate vs. grapevine pests and diseases worldwide: The first results of a global survey. OENO One 2017, 51, 133–139. [Google Scholar] [CrossRef]
- Boso, S.; Gago, P.; Santiago, J.L.; de la Fuente, M.; Martínez, M.C. Factors Affecting the Vineyard Populational Diversity of Plasmopara viticola. Plant Pathol. J. 2019, 35, 125–136. [Google Scholar]
- Martínez-Bracero, M.; Alcázar, P.; Velasco-Jiménez, M.J.; Galán, C. Fungal spores affecting vineyards in Montilla-Moriles southern Spain. Eur. J. Plant Pathol. 2019, 153, 1–13. [Google Scholar] [CrossRef]
- Hidalgo, L. Tratado de Viticultura General, 3rd ed.; Mundi–Prensa: Madrid, Spain, 2002; p. 983. [Google Scholar]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Kan, J.A.L.V. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Kassemeyer, H.H.; Berkelmann-Löhnertz, B. Fungi of grapes. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Berlin/Heidelberg, Germany, 2009; pp. 61–87. [Google Scholar]
- Pszczolkowski, P.; Latorre, B.A.; Ceppi Di Lecco, C. Efectos de los mohos presentes en uvas cosechadas tardíamente sobre la calidad de los mostos y vinos Cabernet Sauvignon. Cienc. Investig. Agrar. 2001, 28, 157–163. [Google Scholar]
- Aleixandre, J.L.; Giner, J.F.; Aleixandre-Tudó, J.L. Evaluación del efecto terroir sobre la calidad de la uva y el vino (I). Enovinicultura 2013, 20, 1–11. [Google Scholar]
- Coertze, S.; Holz, G.; Sadie, A. Germination and establishment of infection on grape berries by single airborne conidia of Botrytis cinerea. Plant Dis. 2001, 85, 668–677. [Google Scholar] [CrossRef] [Green Version]
- Elmer, P.; Michailides, T. Epidemiology of Botrytis cinerea in Orchard and Vine Crops. In Botrytis: Biology, Pathology and Control; Springer: Dordrecht, The Netherlands, 2007; pp. 243–272. [Google Scholar]
- Evans, K.J. Overview of R&D for managing botrytis bunch rot in Australia. In Proceedings of the Australian Society of Viticulture and Oenology Seminar on Grapevine Pests and Disease “Breaking the mould—A Pest and Disease Update”, Mildura, Victoria, Australia, 22–25 July 2008; pp. 4–15. [Google Scholar]
- Dufour, M.C.; Lambert, C.; Bouscaut, J.; Mérillon, J.M.; Corio–Costet, M.F. Benzothiadiazole–primed defence responses and enhanced differential expression of defence genes in Vitis vinifera infected with biotrophic pathogens Erysiphe necator and Plasmopara viticola. Plant Pathol. 2013, 62, 370–382. [Google Scholar] [CrossRef]
- Grove, G.G.; Moyer, M. Podredumbre por Botrytis en la uva para Producción Comercial en Washington: Biología y Manejo de la Enfermedad; Washington State University FS046ES; Washington State University: Washington, DC, USA, 2015; pp. 1–5. [Google Scholar]
- Gadoury, D.; Seem, R.; Wilcox, W.; Henick-Kling, T.; Conterno, L.; Day, A.; Ficke, A. Effects of diffuse colonization of grape berries by Uncinula necator on bunch rots, berry microflora and juice and wine quality. Phytopathology 2007, 97, 1356–1365. [Google Scholar] [CrossRef] [Green Version]
- Oriolani, E.J.A.; Moschini, R.C.; Salas, S.; Martínez, M.I.; Banchero, S. Weather–based models for predicting grape powdery mildew (Uncinula necator (Schwein) Burrill) epidemics. Revista de la Facultad de Ciencias Agrarias Universidad Nacional de Cuyo 2015, 47, 197–211. [Google Scholar]
- Barrios, G.; Reyes, J. Modelización del mildiu en la vid. Phytoma 2004, 164, 1–9. [Google Scholar]
- Cambra, M.; Bernal, I. Plasmopara Viticola (Berk. y Curtis) Berl. & de Toni. Mildiu de la vid. Fichas de Diagnóstico en Laboratorio de Organismos Nocivos de los Vegetales. Ficha 64, 2nd ed.; MAPA: Madrid, Spain, 2006. [Google Scholar]
- Rossi, V.; Caffi, T. Effect of water on germination of Plasmopara viticola oospores. Plant Pathol. 2007, 56, 957–966. [Google Scholar] [CrossRef]
- Díaz, T.; Riquelme, A. Control Integrado en el cultivo de uva de mesa en la Región de Murcia. Phytoma 2012, 239, 48–54. [Google Scholar]
- Lucas, A. Control integrado de plagas y enfermedades en el viñedo. Dossier gestión integrada. Vida Rural, 4 December 2012, pp. 46–51.
- Merlet, H.; Navarro, A.; Rosales, J. Manual Técnico Productivo y Económico Vid. 2016, (Pub. CIREN N° 193). Available online: http://www.bibliotecadigital.ciren.cl/handle/123456789/26087 (accessed on 3 February 2020).
- González-Domínguez, E.; Caffi, T.; Ciliberti, N.; Rossi, V. A mechanistic model of Botrytis cinerea on grapevines that includes weather, vine growth stage, and the main infection pathways. PLoS ONE 2015, 10, e0140444. [Google Scholar]
- Porras, A. Mejora de la Tecnología de la Pulverización de Productos Fitosanitarios sobre Plantaciones de vid en Espaldera. Ph.D. Thesis, Universidad de Córdoba, Córdoba, Spain, 2006. [Google Scholar]
- Rojas, V.; Dennis, M. Nuevas Alternativas de Control Para el oídio de la vid (Uncinula Necator Schw.); Universidad de Chile, Fac. de CC. Agronomicas: Santiago, Chile, 2003; p. 58. [Google Scholar]
- Rodríguez-Rajo, F.J.; Jato, V.; Fernández-González, M.; Aira, M.J. The use of aerobiological methods for forecasting Botrytis spore concentrations in a vineyard. Grana 2010, 49, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Arafat, K.H. Application of statistical model for forecasting powdery mildew of grapes under Egyptian conditions based on meteorological data. Int. J. Plant Pathol. 2015, 6, 48–57. [Google Scholar] [CrossRef] [Green Version]
- MAPAMA. Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente, Registro de Variedades Comerciales. 2017. Available online: www.mapama.gob.es (accessed on 3 February 2020).
- Lorenz, D.H.; Eichorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Phänologische Entwicklungsstadien der Weinrebe (Vitis vinifera L. ssp. vinifera). Codierung und Beschreibung nach der erweiterten BBCH–Skala. Vitic. Enol. Sci. 1994, 49, 66–70. [Google Scholar]
- Hirst, J.M. An automatic volumetric spore–trap. Ann. Appl. Biol. 1952, 36, 257–265. [Google Scholar] [CrossRef]
- Galán, C.; Cariñanos, P.; Alcázar, P.; Domínguez, E. Manual de Calidad y Gestión de la Red Española de Aerobiología; Servicio de Publicaciones, Universidad de Córdoba: Córdoba, Spain, 2007; p. 61. [Google Scholar]
- Galán, C.; Ariatti, A.; Bonini, M.; Clot, B.; Crouzy, B.; Dahl, A.; Fernandez-González, D.; Frenguelli, G.; Gehrig, R.; Isard, S.; et al. Recommended terminology for aerobiological studies. Aerobiologia 2017, 33, 293–295. [Google Scholar] [CrossRef]
- Gargallo, P.; García-Casarejos, N. Impactos ambientales y medidas de mitigación en el sector vitivinícola español. E3S Web of Conferences 50, 01029. In Proceedings of the XII Congreso Internacional Terroir, Zaragoza, Spain, 18–22 June 2018. [Google Scholar]
- Gautam, H.R.; Bhardwaj, M.L.; Robitash, K. Climate change and its impact on plant diseases. Curr. Sci. 2013, 105, 1685–1691. [Google Scholar]
- Casanova, J. Situación actual de la Viticultura Ecológica: Técnicas de producción de la uva y productos autorizados. Vida Rural 2003, 171, 41–45. [Google Scholar]
- Pérez-Sanz, R.; Manzano, Y.; Santiago, L.; de La Iglesia, G.; Campillo, C.; Alberte, L.; Miranda, J.S.; Juárez, I. Metodología para la Validación de Modelos de Desarrollo asociados al clima para el seguimiento del Mildiu, Oidio y Podredumbre Gris en Viñedos de Castilla y León. In Proceedings of the XXVIII Jornadas de Viticultura y Enología de la Tierra de Barros: Cultural Santa Ana, Centro Universitario, Almendralejo, Spain, 8–12 May 2006. [Google Scholar]
- Orlandini, S.; Magarey, R.D.; Park, E.W.; Sporleder, M.; Kroschel, J. Methods of agroclimatology: Modeling approaches for pests and diseases. In Agroclimatology: Linking Agriculture to Climate; Hatfield, J.L., Sivakumar, M.V.K., Prueger, J.H., Eds.; Agron. Monogr. 60; ASA, CSSA, and SSSA: Madison, WI, USA, 2017. [Google Scholar]
- Fernández-González, M.; Rodríguez-Rajo, F.J.; Jato, V.; Aira, M.J. Incidence of fungals in a vineyard of the denomination of origin Ribeiro (Ourense NW Spain). Ann. Agric. Environ. Med. 2009, 16, 263–271. [Google Scholar]
- Calonnec, A.; Cartolaro, P.; Naulin, J.M.; Bailey, D.; Langlais, M. A host–pathogen simulation model: Powdery mildew of grapevine. Plant Pathol. 2008, 57, 493–508. [Google Scholar] [CrossRef]
- Dugan, F.M.; Lupien, S.L.; Grove, G.G. Incidence, aggressiveness and in planta interactions of Botrytis cinerea and other filamentous fungi quiescent in grape berries and dormant buds in central Washington State. Phytopathology 2002, 150, 375–381. [Google Scholar] [CrossRef]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Galet, P. Précis de Viticulture, 7th ed.; Imp. JF: Montpellier, France, 2000. [Google Scholar]
- Boso, S.; Santiago, J.L.; Villaverde-Alonso, V.; Gago, P.; Martínez, M.C.; Rodríguez, E. Evaluación de la incidencia a enfermedades fúngicas en diferentes clones del cv. Albariño (Vitis vinifera L.). Phytoma 2009, 210, 1–5. [Google Scholar]
- Díaz, M.R.; Iglesias, I.; Jato, M.V. Airborne concentrations of Botrytis, Uncinula and Plasmopara spores in a vineyard in Leiro-Ourense (N.W. Spain). Aerobiologia 1997, 13, 31–35. [Google Scholar] [CrossRef]
- Díaz, M.R.; Iglesias, I.; Jato, M.V. Seasonal variation of airborne fungal spore concentrations in a vineyard of North-West Spain. Aerobiologia 1998, 14, 221–227. [Google Scholar] [CrossRef]
- Oliveira, M.; Guerner-Moreira, J.; Mesquita, M.M.; Abreu, I. Important phytopathogenic airborne fungal spores in a rural area: Incidence of Botrytis cinerea and Oidium spp. Ann. Agric. Environ. Med. 2009, 16, 197–204. [Google Scholar]
- Rodríguez-Rajo, F.J.; Seijo Coello, M.C.; Jato, V. Estudio de los niveles de los principales fitopatógenos para la optimización de cosechas de Vitis vinifera en Valdeorras Ourense (1998). Botanica Complutensis 2002, 26, 121–135. [Google Scholar]
- Fernández-González, M.; Rodríguez Rajo, F.J.; Jato, V.; Aira, M.J.; Ribeiro, H.; Oliveira, M.; Abreu, I. Forecasting ARIMA models for atmospheric vineyard pathogens in Galicia and Northern Portugal: Botrytis cinerea spores. Ann. Agric. Environ. Med. 2012, 19, 255–262. [Google Scholar] [PubMed]
- Martínez, C.; Boso, S. Evaluación de la Virulencia de Distintas Poblaciones de Hongos Responsables del Mildiu, oídio y Botrytis en Distintas Denominaciones de Origen Gallegas; Grupo de Viticultura de la MBG–CSIC: Pontevedra, Spain, 2015. [Google Scholar]
- Cortiñas, J.A.; Aira, M.J.; Fernández–González, M.; Rodríguez-Rajo, F.J.; Vázquez–Ruiz, R.A. Potential sustainable wine–growing model in the Ribeira Sacra D.O. (NW Spain). Cienc. Tec. Vitivinic. 2018, 33, 114–145. [Google Scholar]
- Latorre, B.A.; Rioja, M.E. Efecto de la temperatura y humedad relativa sobre la germinación de conidias de Botrytis cinerea. Cien. Inv. Agric. 2001, 29, 67–72. [Google Scholar]
- Molitor, D.; Baus, O.; Hoffmann, L.; Beyer, M. Meteorological conditions determine the thermal-temporal position of the annual Botrytis bunch rot epidemic on Vitis vinifera L. cv. Riesling grapes. OENO One 2016, 50, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Viret, O.; Cole, M. Botrytis cinerea infection in grape flowers: Defense reaction, latency and disease expression. Phytopathology 2003, 93, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Viret, O.; Keller, M.; Jaudzems, V.G.; Cole, F.M. Botrytis cinerea infection of grape flowers: Light and electron microscopical studies of infection sites. Phytopathology 2004, 94, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Latorre, B.A.; Rioja, M.E.; Lillo, C. Efecto de la temperatura en el desarrollo de la infección producida por Botrytis cinerea en flores y bayas de uva de mesa. Cien. Inv. Agric. 2002, 29, 145–151. [Google Scholar] [CrossRef]
- Ciliberti, N.; Fermaud, M.; Languasco, L.; Rossi, V. Influence of fungal strain, temperature, and wetness duration on infection of grapevine inflorescences and young berry clusters by Botrytis cinerea. Phytopathology 2015, 105, 325–333. [Google Scholar] [CrossRef]
- Calonnec, A.; Cartolaro, P.; Poupot, C.; Dubourdieu, D.; Darriet, P. Effects of Uncinula necator on the yield and quality of grapes (Vitis vinifera) and wine. Plant Pathol. 2004, 53, 434–445. [Google Scholar] [CrossRef]
- Campbell, P.; Bendek, C.; Latorre, B.A. Risk of powdery mildew (Erysiphe necator) outbreaks on grapevines in relation to cluster development. Cien. Inv. Agric. 2007, 34, 5–11. [Google Scholar] [CrossRef]
- Fernández-González, M.; Piña–Rey, A.; González–Fernández, E.; Aira, M.J.; Rodríguez-Rajo, F.J. First assessment of Goidanich Index and aerobiological data for Plasmopara viticola infection risk management in north-west Spain. J. Agric. Sci. 2019, 157, 129–139. [Google Scholar] [CrossRef]
- Thind, T.S.; Arora, J.K.; Mohan, C.; Raj, P. Epidemiology of powdery mildew, downy mildew and anthracnose diseases of grapevine. In Diseases of Fruits and Vegetables Volume I; Springer: Dordrecht, The Netherlands, 2004; pp. 621–638. [Google Scholar]
- Chellemi, D.O.; Marois, J.J. Effect of fungicides and water on sporulation of Uncinula necator. Plant Dis. 1991, 75, 455–457. [Google Scholar] [CrossRef]
- Sivapalan, A. Effects of impacting rain drops on the growth and development of powdery mildew fungi. Plant Pathol. 1993, 42, 256–263. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Gubler, W.D.; Grove, G.G. Epidemiology of Powdery Mildews in Agricultural Pathosystems. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; American Phytopathological Society, APS Press: St. Paul, MN, USA, 2002; pp. 169–199. [Google Scholar]
- Campbell, P. Efecto de Factores Ambientales y Métodos de Control Sobre la Germinación y Desarrollo de Uncinula necator en Vitis vinifera. Master’s Thesis, Facultad de Agronomía e Ingeniería Forestal, Universidad Católica de Chile, Santiago, Chile, 2003. [Google Scholar]
- Almendro, J.P. Índices climáticos propios de la vid en el sector central de tierra de barros. In Actas de las IV Jornadas de Historia de Almendralejo y Tierra de Barros; Asociación Histórica de Almendralejo: Almendralejo, Spain, 2013; pp. 121–131. [Google Scholar]
- Thiessen, L.D.; Thiessen, L.; Neill, T.M.; Mahaffee, W.F. Assessment of Erysiphe necator ascospore release models for use in the Mediterranean climate of Western Oregon. Plant Dis. 2018, 102, 1500–1508. [Google Scholar] [CrossRef] [Green Version]
- Magarey, P.A.; Magarey, R.D.; Emmett, R.W. Principles for managing the foliage diseases of grapevine with low input of pesticides. In Proceedings of the 6th International Congress on Organic Viticulture, Basel, Switzerland, 25–26 August 2000; p. 140. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ribeiro | Ribeira Sacra | ||||||
|---|---|---|---|---|---|---|---|
| Year | 2016 | 2017 | 2018 | 2016 | 2017 | 2018 | |
| Annual Average Meteorological Data | Max T (°C) | 22.8 | 23.9 | 23.7 | 20.6 | 21.9 | 21.4 |
| Min T (°C) | 7.3 | 6.4 | 8.2 | 6.7 | 6.1 | 7.3 | |
| Mean T (°C) | 13.8 | 13.9 | 15.2 | 12.8 | 13.1 | 13.7 | |
| Mean RH (%) | 78.4 | 75.8 | 75.9 | 80.9 | 76.9 | 78.6 | |
| Sunshine (hours) | 5.3 | 5.9 | 4.9 | 8.1 | 8.4 | 8.4 | |
| Wind-Speed (Km/h) | 2.1 | 1.8 | 2.2 | 2.1 | 1.8 | 2.5 | |
| Annual Total | Rainfall (L/m2) | 1211.8 | 814.5 | 775.2 | 940.4 | 616.2 | 660.5 |
| Maximum | Daily rainfall (l/m2) | 49.0 | 100.8 | 39.4 | 42.8 | 77.5 | 25.7 |
| Date | 10 Jan | 10 Dec | 28 Feb | 10 Jan | 10 Dec | 11 Mar | |
| Against | S0 | S1 | S5 | S6 | S7 | S8 | |
|---|---|---|---|---|---|---|---|
| Botrytis | Cenlle 2016 | 1 | 2 | ||||
| Cenlle 2017 | 1 | 1 | |||||
| Cenlle 2018 | 2 | 2 | 1 | ||||
| O Mato 2016 | |||||||
| O Mato 2017 | |||||||
| O Mato 2018 | |||||||
| Mildew + Oidium | Cenlle 2016 | 2 | 2 | 2 | |||
| Cenlle 2017 | 2 | 1 | 3 | 1 | |||
| Cenlle 2018 | 1 | 1 | 1 | 1 | 2 | ||
| O Mato 2016 | 1 | 2 | 1 | ||||
| O Mato 2017 | |||||||
| O Mato 2018 | 2 | 1 | 2 | ||||
| Mildew | Cenlle 2016 | 1 | |||||
| Cenlle 2017 | 2 | ||||||
| Cenlle 2018 | 2 | 2 | |||||
| O Mato 2016 | 2 | 2 | |||||
| O Mato 2017 | |||||||
| O Mato 2018 | 1 | 1 | 2 |
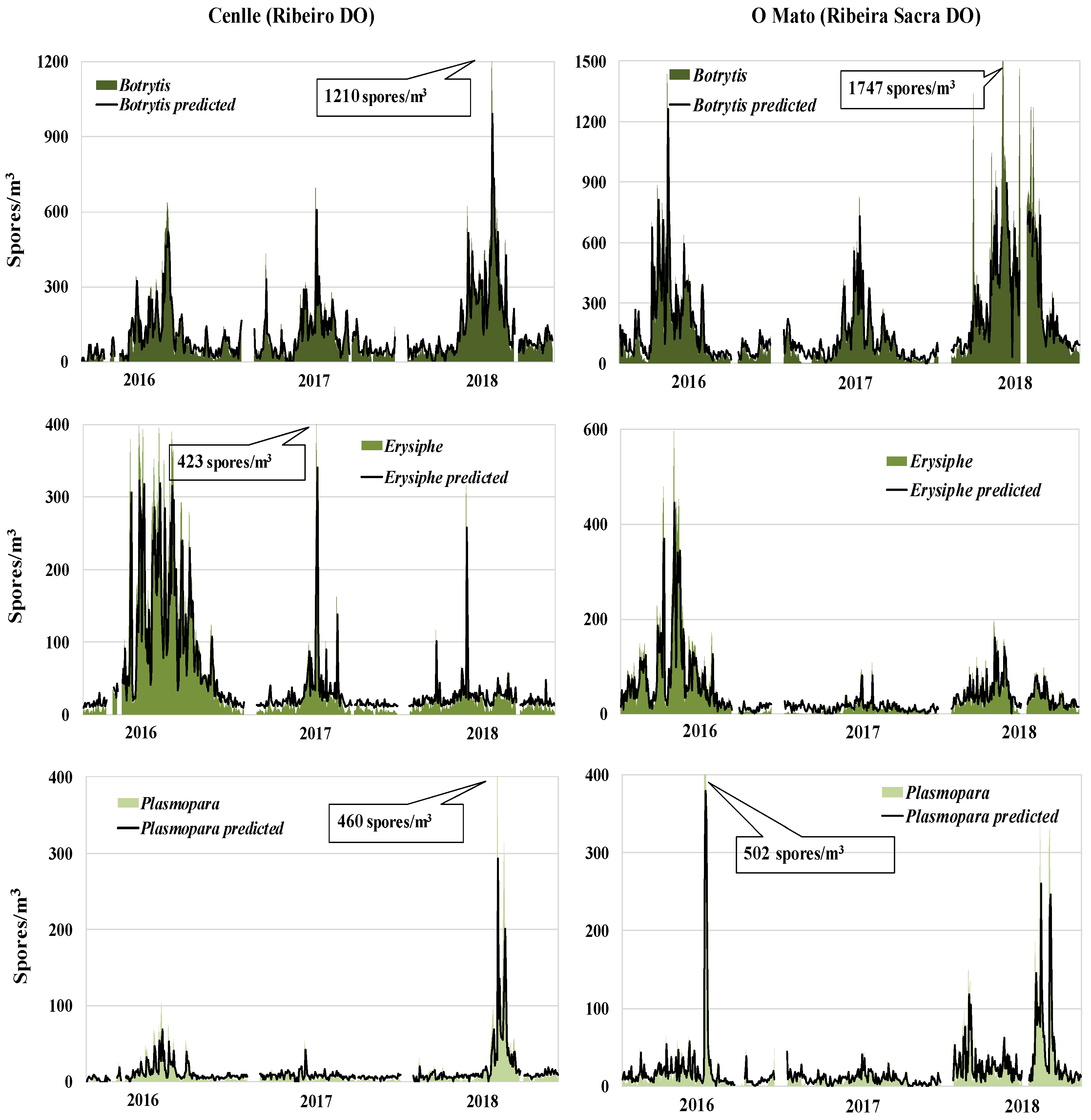
| Cenlle | Botrytis | Erysiphe | Plasmopara |
|---|---|---|---|
| Study Period | 19 September 2016 to 18 September 2016 | ||
| SSIn | 16,806 | 17,269 | 1910 |
| Daily Maximum | 637 | 399 | 104 |
| Maximum Date | 22 June | 16 May | 8 June |
| Study Period | 16 March 2017 to 30 August 2017 | ||
| SSIn | 15,378 | 3344 | 679 |
| Daily Maximum | 696 | 423 | 54 |
| Maximum Date | 28 May 2017 | 26 May 2017 | 8 May 2017 |
| Study Period | 29 March 2018 to 10 September 2018 | ||
| SSIn | 24,214 | 3116 | 3605 |
| Daily maximum | 1210 | 316 | 460 |
| Maximum date | 7 July 2018 | 4 June 2018 | 7 July 2018 |
| O Mato | |||
| Study period | 24 March 2016 to 30 September 2016 | ||
| SSIn | 27,656 | 12,946 | 3618 |
| Daily Maximum | 1435 | 597 | 502 |
| Maximum Date | 27 May 2016 | 3 June 2016 | 7 July 2016 |
| Study Period | 04 March 2017 to 08 September 2017 | ||
| SSIn | 17,240 | 1686 | 1363 |
| Daily Maximum | 826 | 108 | 45 |
| Maximum Date | 4 June 2017 | 19 June 2017 | 3 June 2017 |
| Study Period | 07 April 2018 to 07 September 2018 | ||
| SSIn | 49,620 | 5632 | 5656 |
| Daily Maximum | 1547 | 197 | 345 |
| Maximum date | 9 June 2018 | 29 May 2018 | 22 July 2018 |
| 2016 | 2017 | 2018 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Cenlle | Botrytis | Erysiphe | Plasmopara | Botrytis | Erysiphe | Plasmopara | Botrytis | Erysiphe | Plasmopara |
| Rainfall | −0.167 * Botrytis | −0.365 * Erysiphe-7 | 0.355 * Plasmopara-2 | 0.401 ** Botrytis-4 | 0.196 * Erysiphe-7 | 0.172 * Plasmopara-4 | −0.190 * Botrytis-1 | N.S. | −0.255 ** Plasmopara |
| RH | 0.382 * Botrytis-2 | −0.321 * Erysiphe-2 | 0.342 * Plamopara-2 | 0.507 ** Botrytis-4 | 0.248 ** Erysiphe-5 | N.S. | 0.263 ** Botrytis-5 | 0.203 ** Erysiphe−7 | N.S. |
| T max | 0.288 ** Botrytis | 0.440 ** Erysiphe-2 | −0.335 * Plasmopara-2 | −0.228 ** Botrytis-4 | −0.180 * Erysiphe-7 | −0.300** Plasmopara-6 | 0.387 ** Botrytis | N.S. | 0.387 ** Plasmopara |
| T min | 0.323 ** Botrytis | −0.674 * Erysiphe-2 | 0.346 * Plasmopara-2 | N.S. | N.S. | −0.411 ** Plasmopara-7 | 0.634 ** Botrytis-1 | 0.197 * Erysiphe | 0.489 ** Plasmopara |
| T mean | 0.307 ** Botrytis | −0.666 ** Erysiphe-2 | N.S. | −0.188 * Botrytis-4 | −0.154 * Erysiphe-5 | −0.362 ** Plasmopara-6 | 0.556 ** Botrytis | N.S. | 0.480 ** Plasmopara |
| O Mato | |||||||||
| Rainfall | 0.365 ** Botrytis-5 | 0.394 ** Erysiphe-6 | 0.227 ** Plasmopara-6 | N.S. | −0.272 ** Erysiphe-2 | N.S. | 0.224 ** Botrytis-6 | 0.231 ** Erysiphe-7 | N.S. |
| RH | 0.593 ** Botrytis-2 | 0.491** Erysiphe-6 | 0.377** Plamopara-2 | 0.224 ** Botrytis-3 | −0.354 ** Erysiphe-2 | 0.163 * Plasmopara-4 | 0.373 ** B otrytis-7 | 0.359 ** Erysiphe-6 | N.S. |
| T max | −0.386 ** Botrytis-7 | −0.476 ** Erysiphe-7 | −0.309 ** Plasmopara-7 | N.S. | 0.483 ** Erysiphe | 0.186 * Plasmopara | N.S. | −0.297 ** Erysiphe-6 | N.S. |
| T min | −0.153 * Botrytis-7 | −0.231 ** Erysiphe-6 | −0.177* Plasmopara-7 | 0.248 ** Botrytis | 0.509 ** Erysiphe | 0.209 ** Plasmopara | 0.331 ** Botrytis | N.S. | N.S. |
| T mean | −0.367 ** Botrytis-7 | −0.439 ** Erysiphe-7 | −0.332 ** Plasmopara-7 | 0.224 ** Botrytis | 0.582 ** Erysiphe | 0.218 ** Plasmopara | 0.169 * Botrytis | −0.251 ** Erysiphe-6 | N.S. |
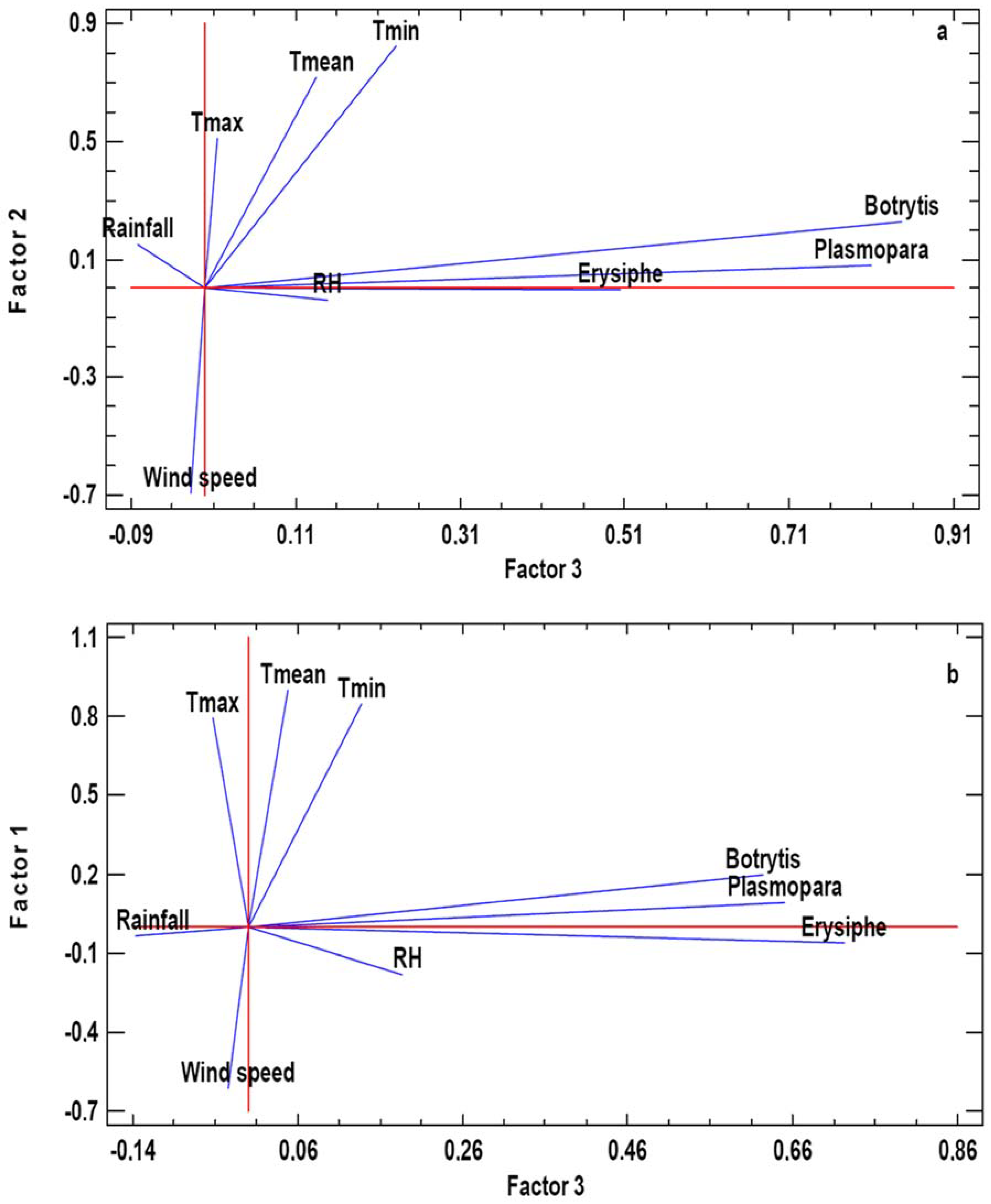
| CENLLE | O MATO | |||||
|---|---|---|---|---|---|---|
| Principal Components | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 |
| Self-value | 3.298 | 1.956 | 1.135 | 3.184 | 1.807 | 1.098 |
| Variance (%) | 36.640 | 21.729 | 12.610 | 35.376 | 20.087 | 12.207 |
| Accumulated percentage | 36.640 | 58.369 | 70.980 | 35.376 | 55.463 | 67.671 |
| Botrytis | −0.003 | 0.227 | 0.846 | 0.196 | 0.414 | 0.624 |
| Erysiphe | 0.012 | −0.001 | 0.504 | −0.065 | 0.011 | 0.723 |
| Plasmopara | −0.008 | 0.076 | 0.809 | 0.088 | −0.109 | 0.650 |
| Max T | −0.800 | 0.512 | 0.015 | 0.796 | −0.518 | −0.043 |
| Min T | −0.158 | 0.826 | 0.232 | 0.848 | 0.047 | 0.137 |
| Mean T | −0.643 | 0.719 | 0.135 | 0.901 | −0.381 | 0.047 |
| RH | 0.937 | −0.037 | 0.149 | −0.185 | 0.881 | 0.186 |
| Rainfall | 0.805 | 0.152 | −0.082 | −0.037 | 0.817 | −0.137 |
| Wind speed | −0.132 | −0.697 | −0.018 | −0.612 | −0.075 | −0.024 |
| Cenlle | Beta | B | Std. Error B | t | p < |
|---|---|---|---|---|---|
| Botrytis | R = 0.843 | R2 = 0.710 | Adjusted R2 = 0.709 | ||
| Intercepted | −197.800 | 42.778 | −4.624 | 0.000 | |
| Botrytis-1 | 0.768 | 0.768 | 0.027 | 28.068 | 0.000 |
| T mean | 0.124 | 3.650 | 0.888 | 4.111 | 0.000 |
| RH-3 | 0.144 | 2.247 | 0.468 | 4.800 | 0.000 |
| Erysiphe | R = 0.782 | R2 = 0.611 | Adjusted R2 = 0.610 | ||
| Intercepted | 10.038 | 2.528 | 3.971 | 0.000 | |
| Erysiphe-1 | 0.782 | 0.782 | 0.028 | 28.199 | 0.000 |
| Plasmopara | R = 0.633 | R2 = 0.401 | Adjusted R2 = 0.399 | ||
| Intercepted | 12.016 | 3.871 | 3.105 | 0.000 | |
| Plasmopara-1 | 0.620 | 0.620 | 0.035 | 17.851 | 0.000 |
| Wind Speed-1 | −0.071 | −3.278 | 1.609 | −2.037 | 0.042 |
| O Mato | |||||
| Botrytis | R = 0.722 | R2 = 0.596 | Adjusted R2 = 0.594 | ||
| Intercepted | −136.199 | 63.573 | −2.142 | 0.033 | |
| Botrytis-1 | 0.734 | 0.827 | 0.035 | 23.587 | 0.009 |
| RH-3 | 0.082 | 2.361 | 0.895 | 2.637 | 0.000 |
| Erysiphe | R = 0.760 | R2 = 0.578 | Adjusted R2 = 0.576 | ||
| Intercepted | −54.391 | 17.578 | −3.094 | 0.002 | |
| Erysiphe-1 | 0.737 | 0.737 | 0.029 | 25.346 | 0.000 |
| RH-3 | 0.107 | 0.885 | 0.240 | 3.680 | 0.000 |
| Plasmopara | R = 0.480 | R2 = 0.560 | Adjusted R2 = 0.558 | ||
| Intercepted | −24.085 | 12.483 | −1.929 | 0.054 | |
| Plasmopara-1 | 0.742 | 0.742 | 0.029 | 25.282 | 0.000 |
| RH-1 | 0.070 | 0.402 | 0.170 | 2.370 | 0.018 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortiñas Rodríguez, J.A.; González-Fernández, E.; Fernández-González, M.; Vázquez-Ruiz, R.A.; Aira, M.J. Fungal Diseases in Two North-West Spain Vineyards: Relationship with Meteorological Conditions and Predictive Aerobiological Model. Agronomy 2020, 10, 219. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020219
Cortiñas Rodríguez JA, González-Fernández E, Fernández-González M, Vázquez-Ruiz RA, Aira MJ. Fungal Diseases in Two North-West Spain Vineyards: Relationship with Meteorological Conditions and Predictive Aerobiological Model. Agronomy. 2020; 10(2):219. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020219
Chicago/Turabian StyleCortiñas Rodríguez, Jose A., Estefanía González-Fernández, María Fernández-González, Rosa A. Vázquez-Ruiz, and María Jesús Aira. 2020. "Fungal Diseases in Two North-West Spain Vineyards: Relationship with Meteorological Conditions and Predictive Aerobiological Model" Agronomy 10, no. 2: 219. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020219