Transcriptomic Analysis of Female Panicles Reveals Gene Expression Responses to Drought Stress in Maize (Zea mays L.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Drought Stress Treatments
2.3. Measurement of Morphology and Microscopic Observation of Female Panicles
2.4. RNA Isolation and Illumina Sequencing
2.5. Read Mapping and Differential Expression Analysis
2.6. GO and KEGG Enrichment Analysis
2.7. Differential Expression Verification by Quantitative Real-Time PCR (qRT-PCR)
2.8. Statistical Analysis
3. Results
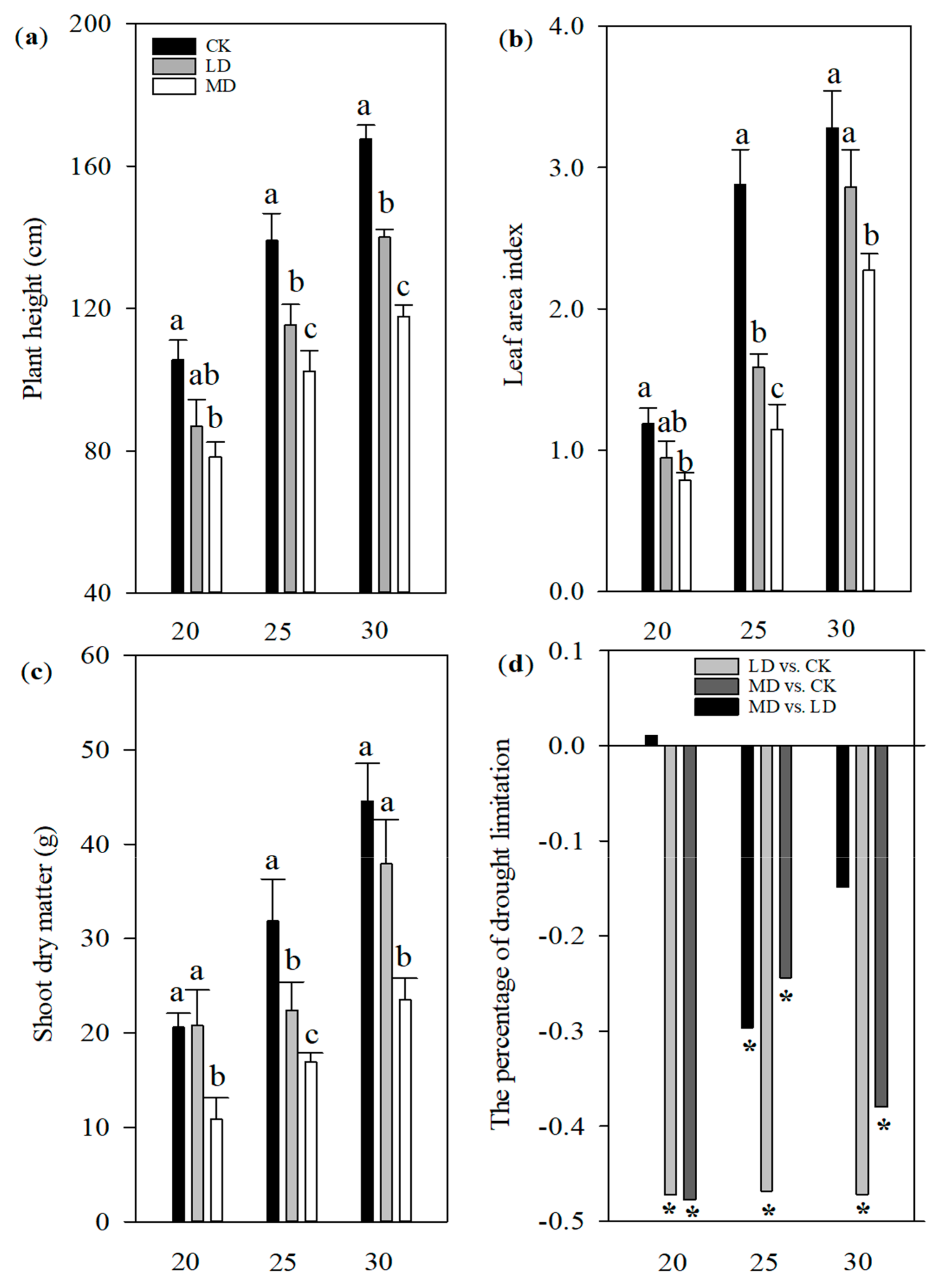
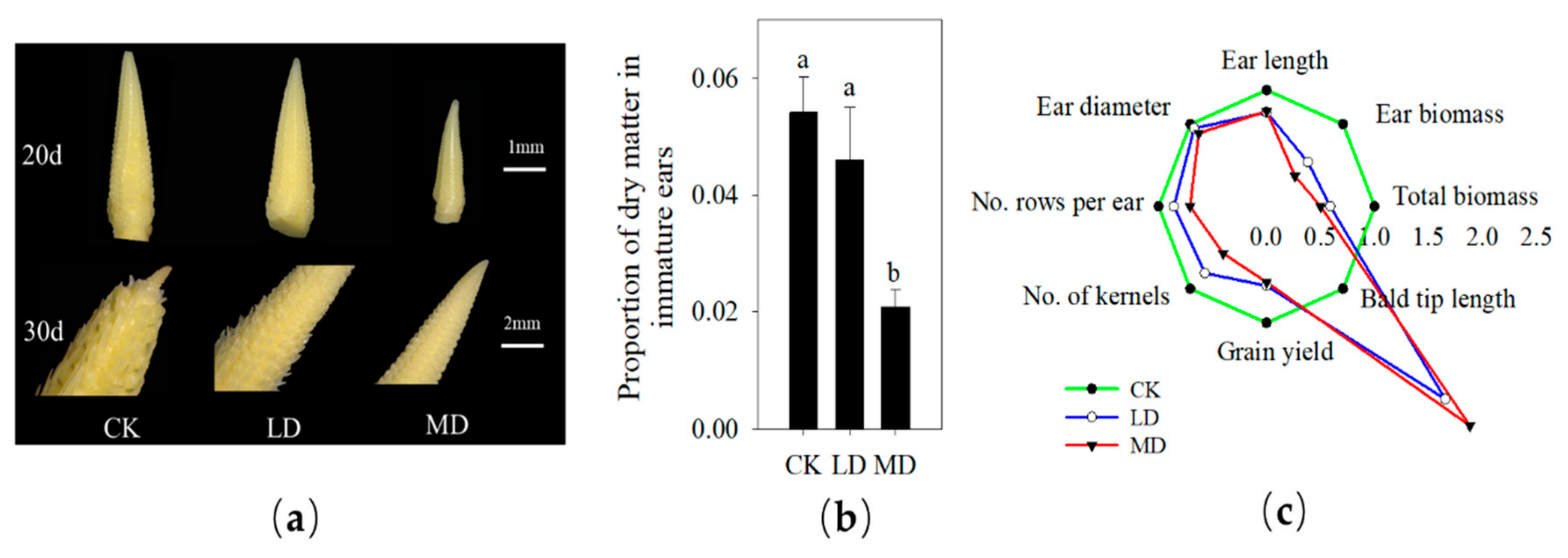
3.1. Female Panicle Development, Phenotypic Change and Yield Components
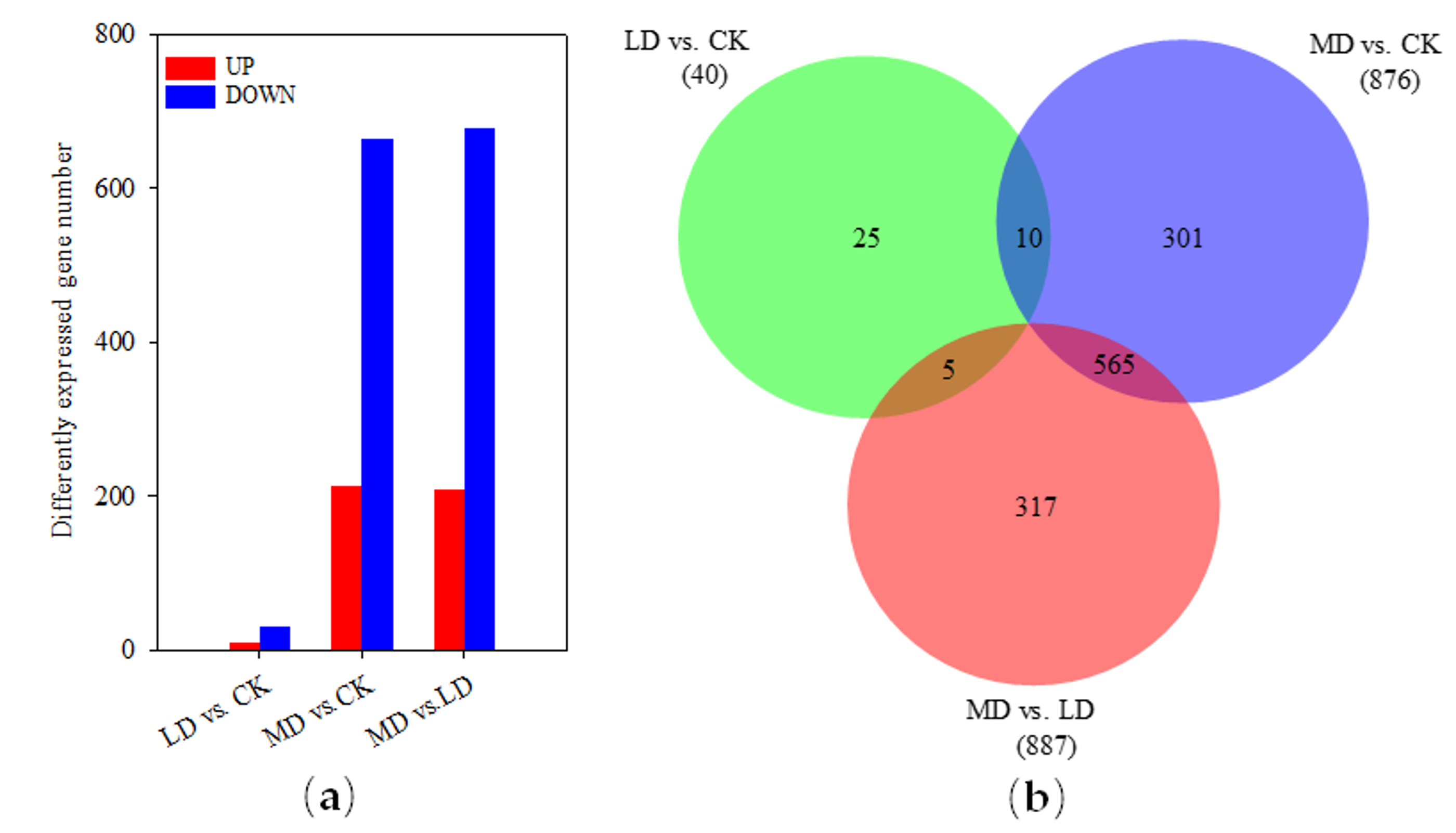
3.2. Overview of RNA Sequencing and Mapping
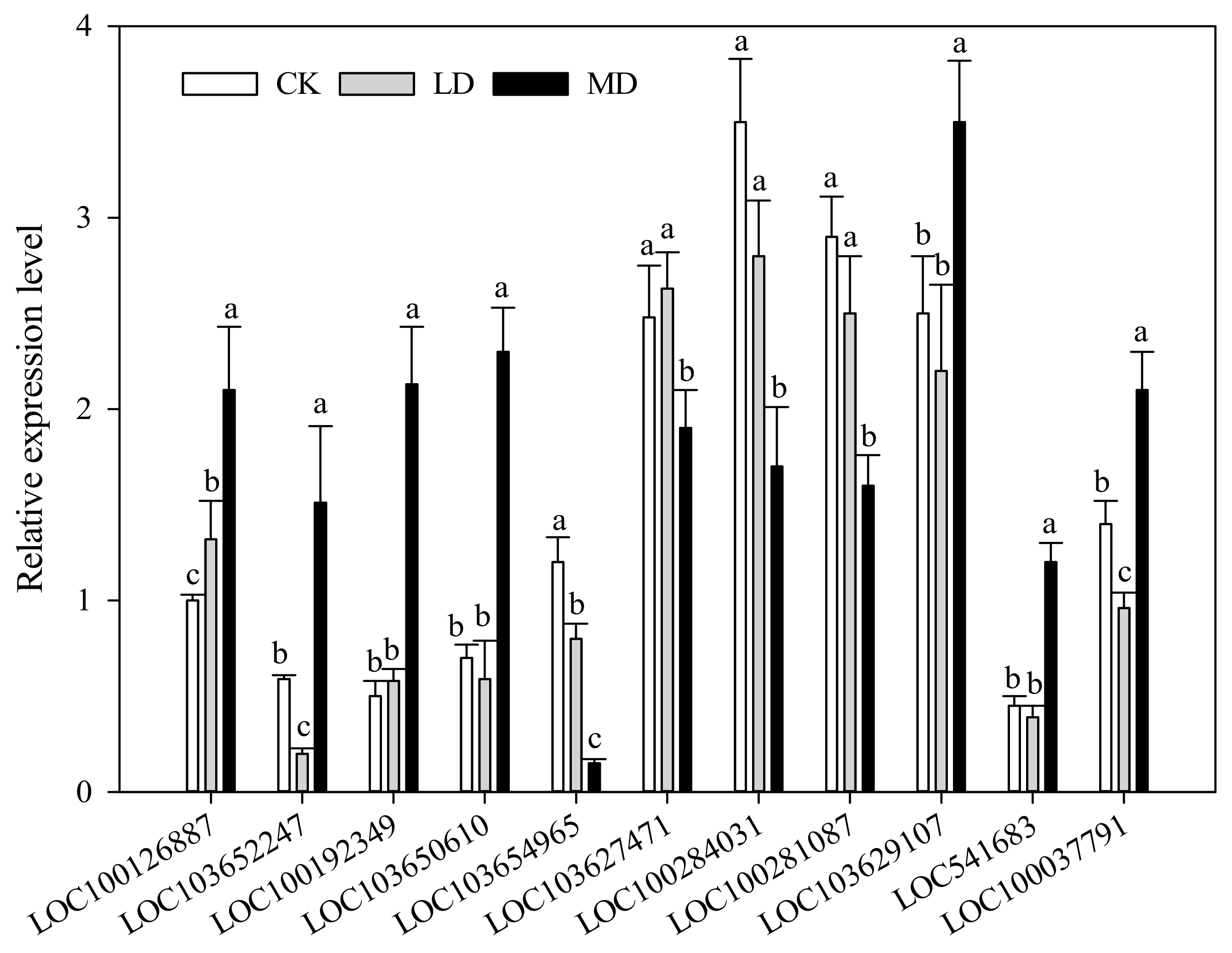
3.3. Gene Expression Validation by qRT-PCR
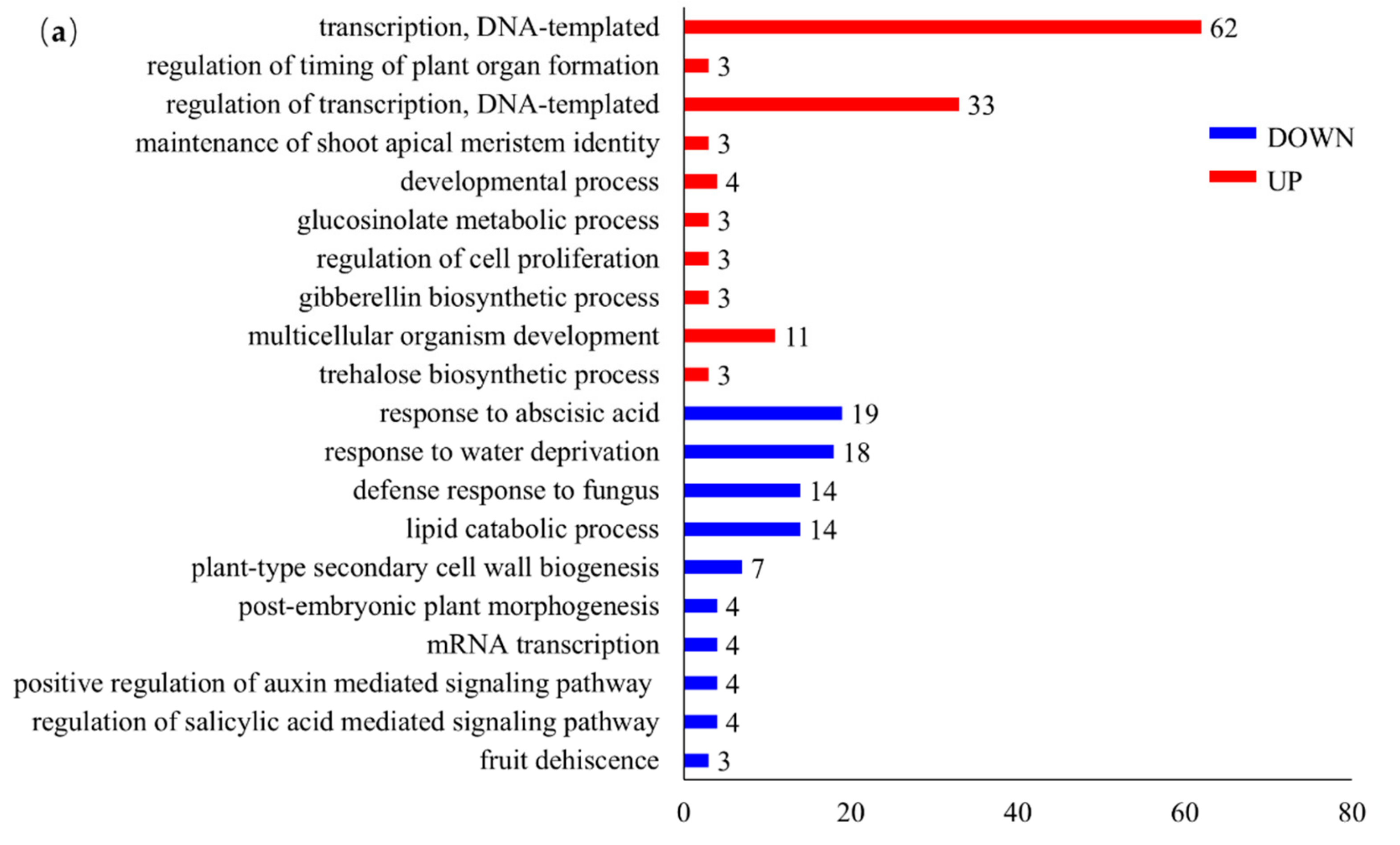
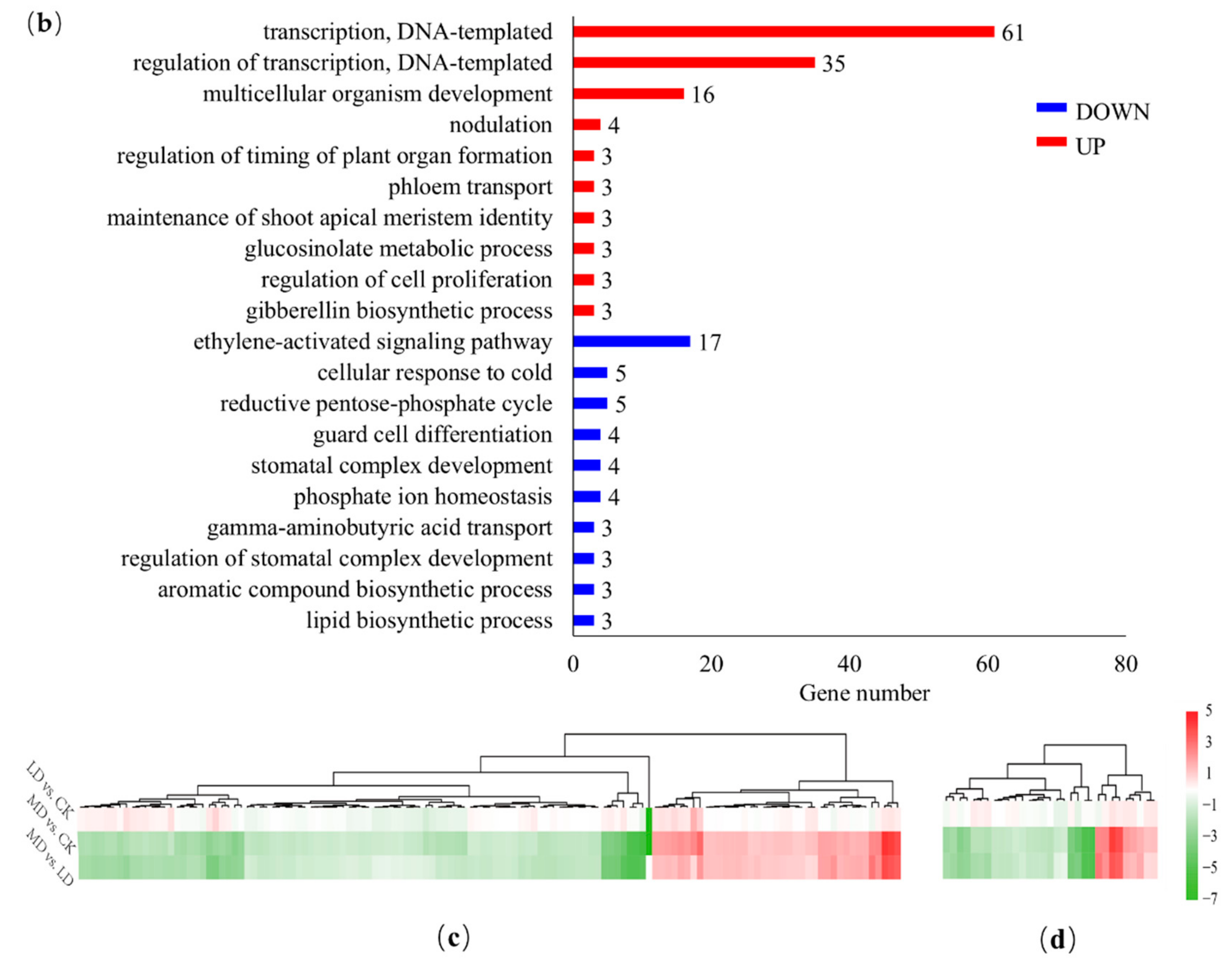
3.4. GO Annotation and Enrichment
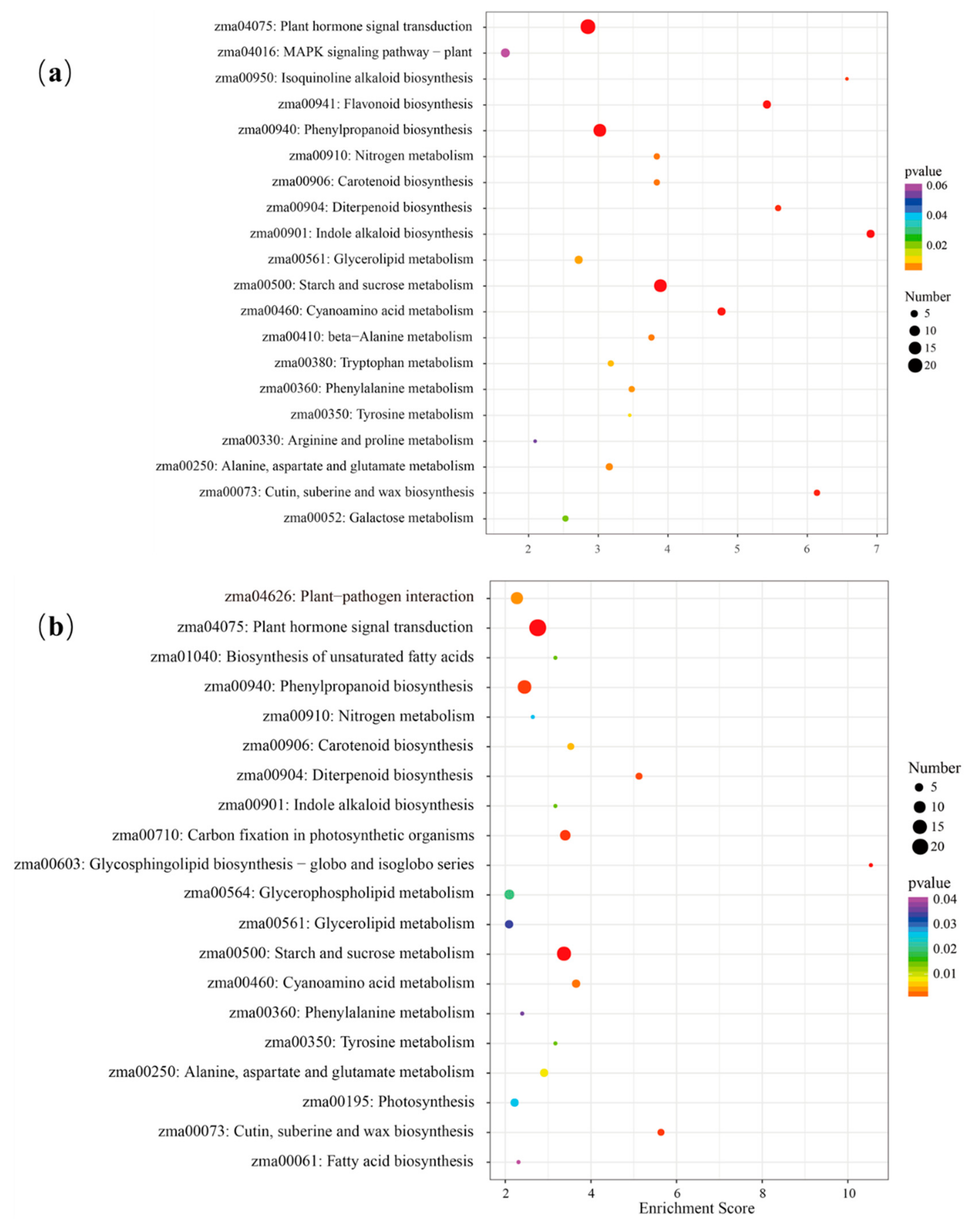
3.5. Metabolic Pathways Related to Soil Drought Stress
4. Discussion
4.1. Responses of Plant Growth and Female Panicle Differentiation to Soil Drought Stress
4.2. Genes Involved in Development and Growth in Response to Soil Drought Stress
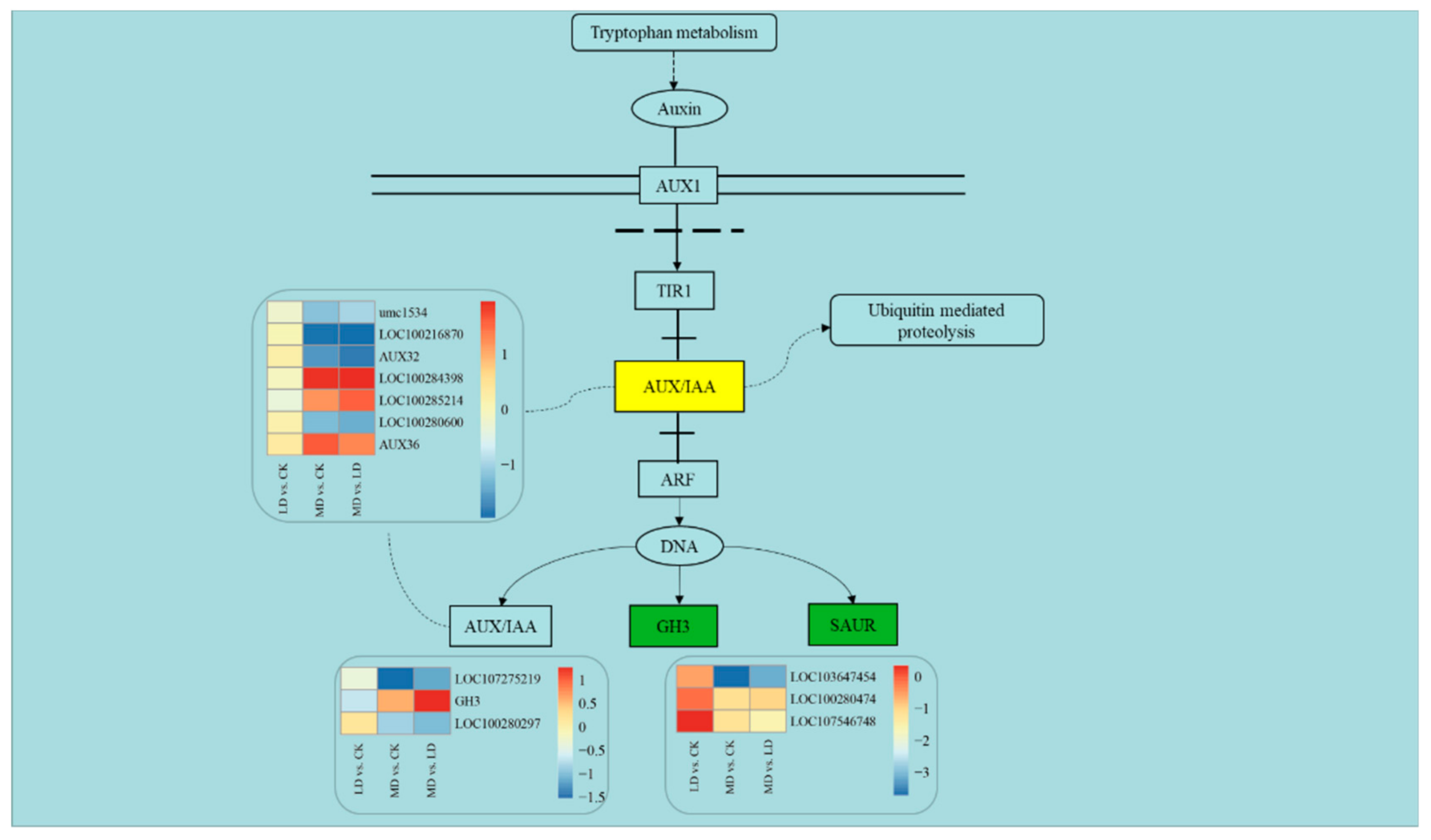
4.3. Genes Involved in Auxin Signaling in Response to Soil Drought Stress
4.4. Reactive Oxygen Scavenging System and Ion Channel in Response to Soil Drought Stress
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lisar, S.Y.S.; Motafakkerazad, R.; Hossain, M.M.; Rahman, I.M.M. Water Stress in Plants: Causes, Effects and Responses. In Water Stress; Rahman, M.M., Hasegawa, H., Eds.; Intech: Rijeka, Croatia, 2012; pp. 1–14. ISBN 978-953-307-963-9. [Google Scholar]
- Dai, A. Drought under global warming: A review. WIREs Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Rollins, J.A.; Habte, E.; Templer, S.E.; Colby, T.; Schmidt, J.; Von Korff, M. Leaf proteome alterations in the context of physiological and morphological responses to drought and heat stress in barley (Hordeum vulgare L.). J. Exp. Bot. 2013, 64, 3201–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potopová, V.; Boroneanţ, C.; Boincean, B.; Soukup, J. Impact of agricultural drought on main crop yields in the Republic of Moldova. Int. J. Climatol. 2016, 36, 2063–2082. [Google Scholar] [CrossRef] [Green Version]
- Daryanto, S.; Wang, L.X.; Jacinthe, P.A. Global synthesis of drought effects on food legume production. PLoS ONE 2015, 10, e0127401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.R.; Lu, X.M.; Zhang, P.Y.; Wang, G.R.; Wei, L.; Wang, T.C. Systematic Analysis of Differentially Expressed Maize ZmbZIP Genes between Drought and Rewatering Transcriptome Reveals bZIP Family Members Involved in Abiotic Stress Responses. Int. J. Mol. Sci. 2019, 20, 4103. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Rao, M.J.; Anjum, M.A.; Ejaz, S.; Zakir, I.; Ali, M.A.; Ahmad, N.; Ahmad, S. Oxidative stress and antioxidant defense in plants under drought conditions. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K.R., Nahar, K., Alharby, H.F., Eds.; Springer: Cham, Switzerland, 2019; pp. 207–219. ISBN 978-3-030-06117-3. [Google Scholar]
- Meeks, M.; Murray, S.; Hague, S.; Hays, D. Measuring maize seedling drought response in search of tolerant germplasm. Agronomy 2013, 3, 135–147. [Google Scholar] [CrossRef]
- Vaughan, M.M.; Block, A.; Christensen, S.A.; Allen, L.H.; Schmelz, E.A. The effects of climate change associated abiotic stresses on maize phytochemical defenses. Phytochem. Rev. 2018, 17, 37–49. [Google Scholar] [CrossRef]
- Calanca, P.P. Effects of abiotic stress in crop production. In Quantification of Climate Variability, Adaptation and Mitigation for Agricultural Sustainability; Ahmed, M., Stockle, C.O., Eds.; Springer: Cham, Switzerland, 2017; pp. 165–180. ISBN 978-3-319-32057-1. [Google Scholar]
- EL Sabagh, A.; Hossain, A.; Islam, M.S.; Barutcular, S.; Fahad, S.; Ratnasekera, D.; Kumar, N.; Meena, R.S.; Vera, P.; Saneoka, H. Role of osmoprotectants and soil amendments for sustainable soybean (Glycine max L.) production under drought condition: A review. J. Exp. Biol. Agric. Sci. 2018, 6, 32–41. [Google Scholar] [CrossRef]
- Yang, M.; Geng, M.Y.; Shen, P.F.; Chen, X.H.; Li, Y.J.; Wen, X.X. Effect of post-silking drought stress on the expression profiles of genes involved in carbon and nitrogen metabolism during leaf senescence in maize (Zea mays L.). Plant Physiol. Biochem. 2019, 135, 304–309. [Google Scholar] [CrossRef]
- Chiuta, N.; Mutengwa, C. Response of yellow quality protein maize inbred lines to drought stress at seedling stage. Agronomy 2018, 8, 287. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.P.; Li, G.S.; Ryu, C.H.; Ma, C.; Zhang, S.S.; Lloyd, A.; Hunter, B.G.; Larkins, B.A.; Drews, G.N.; Wang, X.F.; et al. Opaque-2 regulates a complex gene network associated with cell differentiation and storage functions of maize endosperm. Plant Cell 2018, 30, 2425–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, D.; Ashraf, N.; Khurana, J.P.; Kameshwari, M.N. The ‘Omics’ Approach for Crop Improvement Against Drought Stress. In Genetic Enhancement of Crops for Tolerance to Abiotic Stress: Mechanisms and Approaches; Rajpal, V.R., Sehgal, D., Kumar, A., Raina, S.N., Eds.; Springer: Cham, Switzerland, 2019; pp. 183–204. ISBN 978-3-319-91955-3. [Google Scholar]
- Dastogeer, K.M.G.; Li, H.; Sivasithamparam, K.; Jones, M.G.K.; Wylie, S.J. Fungal endophytes and a virus confer drought tolerance to Nicotiana benthamiana plants through modulating osmolytes, antioxidant enzymes and expression of host drought responsive genes. Environ. Exp. Bot. 2018, 149, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Fang, Y.J.; Xiong, L.Z. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, F.; Qi, W.W.; Lv, Y.D.; Yan, S.M.; Xu, L.M.; Yang, W.Y.; Yuan, Y.; Chen, Y.H.; Zhao, H.; Song, R.T. OPAQUE11 is a central hub of the regulatory network for maize endosperm development and nutrient metabolism. Plant Cell 2018, 30, 375–396. [Google Scholar] [CrossRef] [Green Version]
- An, Y.X.; Chen, L.; Li, Y.X.; Li, C.H.; Shi, Y.S.; Song, Y.C.; Zhang, D.F.; Li, Y.; Wang, T.Y. Candidate loci for the kernel row number in maize revealed by a combination of transcriptome analysis and regional association mapping. BMC Plant Biol. 2019, 19, 201. [Google Scholar] [CrossRef]
- Wilson, J. Control of Maize Development by MicroRNA and Auxin Regulated Pathways; East Carolina University: Greenville, NC, USA, 2018. [Google Scholar]
- Opitz, N.; Paschold, A.; Marcon, C.; Malik, W.A.; Lanz, C.; Piepho, H.P.; Hochholdinger, F. Transcriptomic complexity in young maize primary roots in response to low water potentials. BMC Genomics 2014, 15, 741. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Kim, H.C.; Shin, S.; Kim, K.H.; Moon, J.C.; Kim, J.Y.; Lee, B.M. Transcriptome analysis of flowering time genes under drought stress in maize leaves. Front. Plant Sci. 2017, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Li, L. The Major Metabolic Pathways at Maize Developing Young Tassel in Response to Drought Stress and Identification of Drought-Tolerant Candidate SNAC Genes; Xinjiang Agricultural University: Wulumuqi, Xinjiang, China, 2015. [Google Scholar]
- Wang, B.M.; Liu, C.; Zhang, D.F.; He, C.M.; Zhang, J.R.; Li, Z.X. Effects of maize organ-specific drought stress response on yields from transcriptome analysis. BMC Plant Biol. 2019, 19, 335. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Wang, Q.X.; Wang, K.J.; Li, C.H.; Hao, J.P. Maize. In Crop Cultivation Science: North; Yu, Z.W., Ed.; China Agriculture Press: Beijing, China, 2003; pp. 69–111. ISBN 9787109179363. [Google Scholar]
- Chen, D.Q.; Wang, S.W.; Cao, B.B.; Cao, D.; Leng, G.H.; Li, H.B.; Yin, L.N.; Shan, L.; Deng, X.P. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oury, V.; Caldeira, C.F.; Prodhomme, D.; Pichon, J.P.; Gibon, Y.; Turc, O. Is change in ovary carbon status a cause or a consequence of maize ovary abortion in water deficit during flowering? Plant Physiol. 2016, 171, 997–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khandagale, S.G.; Dubey, R.B.; Sharma, V.; Khan, R. Response of physiological traits of maize to moisture stress induced at different developmental stages. Int. J. Chem. Stud. 2018, 6, 2757–2761. [Google Scholar]
- Ma, C.Y.; Li, B.; Wang, L.N.; Xu, M.L.; Zhu, L.E.; Jin, H.Y.; Wang, Z.C.; Ye, J.R. Characterization of phytohormone and transcriptome reprogramming profiles during maize early kernel development. BMC Plant Biol. 2019, 19, 197. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.J.; Li, H.W.; Jiang, Y.P.; Zhao, G.Q.; Wang, H.Z.; Yang, S.J.; Yang, Q.H.; Guo, J.M.; Shao, R.X. Effects of drought on photosynthesis and ear development characteristics of maize. Acta Ecol. Sin. 2020, 3, 1–9. [Google Scholar]
- Li, Y.B.; Song, H.; Zhou, L.; Xu, Z.Z.; Zhou, G.S. Tracking chlorophyll fluorescence as an indicator of drought and rewatering across the entire leaf lifespan in a maize field. Agric. Water Manag. 2019, 211, 190–201. [Google Scholar] [CrossRef]
- Xu, Z.Z.; Zhou, G.S.; Shimizu, H. Are plant growth and photosynthesis limited by pre-drought following rewatering in grass? J. Exp. Bot. 2009, 60, 3737–3749. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.P.; Peluso, P.; Shi, J.H.; Liang, T.; Stitzer, M.C.; Wang, B.; Campbell, M.S.; Stein, J.C.; Wei, X.H.; Chin, C.S.; et al. Improved maize reference genome with single-molecule technologies. Nature 2017, 546, 524–527. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.; Pimentel, H.; Trapnell, C.; Pachter, L. Identification of novel transcripts in annotated genomes using RNA-Seq. Bioinformatics 2011, 27, 2325–2329. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level–the DESeq Package; European Molecular Biology Laboratory: Heidelberg, Germany, 2012. [Google Scholar]
- Abendroth, L.J.; Elmore, R.W.; Boyer, M.J.; Marlay, S.K. Vegetative Stages (VE to VT). In Corn Growth and Development; Iowa State University: Ames, IA, USA, 2011; pp. 13–27. [Google Scholar]
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Ray, D.K.; Ramankutty, N.; Foley, J.A. Closing yield gaps through nutrient and water management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef]
- Zhang, X.B.; Lei, L.; Lai, J.S.; Zhao, H.M.; Song, W.B. Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings. BMC Plant Biol. 2018, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Hayano-Kanashiro, C.; Calderón-Vázquez, C.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Simpson, J. Analysis of gene expression and physiological responses in three Mexican maize landraces under drought stress and recovery irrigation. PLoS ONE 2009, 4, e7531. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.J.; Wang, M.P.; Wei, Y.Y.; Xia, Z.L. Overexpression of the maize psbA gene enhances drought tolerance through regulating antioxidant system, photosynthetic capability, and stress defense gene expression in tobacco. Front. Plant Sci. 2016, 6, 1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avramova, V.; AbdElgawad, H.; Vasileva, I.; Petrova, A.S.; Holek, A.; Mariën, J.; Asard, H.; Beemster, G.T.S. High antioxidant activity facilitates maintenance of cell division in leaves of drought tolerant maize hybrids. Front. Plant Sci. 2017, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Witt, S.; Galicia, L.; Lisec, J.; Cairns, J.; Tiessen, A.; Araus, J.L.; Palacios-Rojas, N.; Fernie, A.R. Metabolic and phenotypic responses of greenhouse-grown maize hybrids to experimentally controlled drought stress. Mol. Plant 2012, 5, 401–417. [Google Scholar] [CrossRef] [Green Version]
- Bartels, D.; Souer, E. Molecular responses of higher plants to dehydration. In Plant Responses to Abiotic Stress; Hirt, H., Shinozaki, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 9–38. ISBN 9783-540-20037-6. [Google Scholar]
- Boonjung, H.; Fukai, S. Effects of soil water deficit at different growth stages on rice growth and yield under upland conditions. 2. Phenology, biomass production and yield. Field Crops Res. 1996, 48, 47–55. [Google Scholar] [CrossRef]
- Su, Z.; Ma, X.; Guo, H.H.; Sukiran, N.L.; Guo, B.; Assmann, S.M.; Ma, H. Flower development under drought stress: Morphological and transcriptomic analyses reveal acute responses and long-term acclimation in Arabidopsis. Plant Cell 2013, 25, 3785–3807. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, V.H.; Lee, E.A.; Lukens, L.L.; Swanton, C.J. The relationship between floret number and plant dry matter accumulation varies with early season stress in maize (Zea mays L.). Field Crops Res. 2019, 238, 129–138. [Google Scholar] [CrossRef]
- Nielsen, R.L. Ear Size Determination in Corn; Corny News Network Articles; Purdue University: West Lafayette, IN, USA, 2007. [Google Scholar]
- Hu, X.J.; Wang, H.W.; Diao, X.Z.; Liu, Z.F.; Li, K.; Wu, Y.J.; Liang, Q.J.; Wang, H.; Huang, C.L. Transcriptome profiling and comparison of maize ear heterosis during the spikelet and floret differentiation stages. BMC Genomics 2016, 17, 959. [Google Scholar] [CrossRef] [Green Version]
- Jeffares, D.C. Molecular Genetic Analysis of the Maize Terminal Ear1 Gene and in Silico Analysis of Related Genes; Massey University: Palmerston North, New Zealand, 2001. [Google Scholar]
- Vroemen, C.W.; Mordhorst, A.P.; Albrecht, C.; Kwaaitaal, M.A.C.J.; Vries, S.C. The CUP-SHAPED COTYLEDON3 gene is required for boundary and shoot meristem formation in Arabidopsis. Plant Cell 2003, 15, 1563–1577. [Google Scholar] [CrossRef]
- Hibara, K.; Karim, M.R.; Takada, S.; Taoka, K.; Furutani, M.; Aida, M.; Tasaka, M. Arabidopsis CUP-SHAPED COTYLEDON3 regulates postembryonic shoot meristem and organ boundary formation. Plant Cell 2006, 18, 2946–2957. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.H.; Chen, X.J.; Xie, K.; Xing, Q.K.; Wu, Y.W.; Li, J.; Du, C.H.; Sun, Z.X.; Guo, Z.J. Dlf1, a WRKY transcription factor, is involved in the control of flowering time and plant height in rice. PLoS ONE 2014, 9, e102529. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.H.; Yuan, Y.; Fu, D.; Shen, C.J.; Yang, Y.J. Identification and expression profiling of the auxin response factors in Dendrobium officinale under abiotic stresses. Int. J. Mol. Sci. 2017, 18, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porco, S.; Larrieu, A.; Du, Y.J.; Gaudinier, A.; Goh, T.; Swarup, K.; Swarup, R.; Kuempers, B.; Bishopp, A.; Lavenus, J.; et al. Lateral root emergence in Arabidopsis is dependent on transcription factor LBD29 regulation of auxin influx carrier LAX3. Development 2016, 143, 3340–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinesh, D.C.; Villalobos, L.I.A.C.; Abel, S. Structural biology of nuclear auxin action. Trends Plant Sci. 2016, 21, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.R.; Lee, D.K.; Do Choi, Y.; Kim, J.K. OsIAA6, a member of the rice Aux/IAA gene family, is involved in drought tolerance and tiller outgrowth. Plant Sci. 2015, 236, 304–312. [Google Scholar] [CrossRef]
- Asghar, M.A.; Li, Y.; Jiang, H.K.; Sun, X.; Ahmad, B.; Imran, S.; Yu, L.; Liu, C.Y.; Yang, W.Y.; Du, J.B. Crosstalk between Abscisic Acid and Auxin under Osmotic Stress. Agron. J. 2019, 111, 2157–2162. [Google Scholar] [CrossRef]
- Kelley, K.B.; Riechers, D.E. Recent developments in auxin biology and new opportunities for auxinic herbicide research. Pestic. Biochem. Phys. 2007, 89, 1–11. [Google Scholar] [CrossRef]
- Han, S.; Hwang, I. Integration of multiple signaling pathways shapes the auxin response. J. Exp. Bot. 2017, 69, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Zhou, J.J.; Zhang, J.Z. Aux/IAA gene family in plants: Molecular structure, regulation, and function. Int. J. Mol. Sci. 2018, 19, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Q.S.; Hou, D.; Li, L.; Cheng, Z.C.; Ge, W.; Liu, J.; Li, X.P.; Mu, S.H.; Gao, J. Genome-wide analysis and expression characteristics of small auxin-up RNA (SAUR) genes in moso bamboo (Phyllostachys edulis). Genome 2016, 60, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Gray, W.M. SAUR proteins as effectors of hormonal and environmental signals in plant growth. Mol. Plant 2015, 8, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Han, D.G.; Zhang, Z.Y.; Ding, H.B.; Chai, L.J.; Liu, W.; Li, H.X.; Yang, G.H. Isolation and characterization of MbWRKY2 gene involved in enhanced drought tolerance in transgenic tobacco. J. Plant Interact. 2018, 13, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Malagoli, M.; Wirtz, M.; Hell, R. Drought Stress in Maize Causes Differential Acclimation Responses of Glutathione and Sulfur Metabolism in Leaves and Roots. BMC Plant Biol. 2016, 16, 247. [Google Scholar] [CrossRef]
- Zhang, X.; Tao, L.; Qiao, S.; Du, B.H.; Guo, C.H. Roles of Glutathione S-transferase in Plant Tolerance to Abiotic Stresses. J. Chin. Biotechnol. 2017, 37, 92–98. [Google Scholar]
- Kong, X.X.; Li, B.Z.; Yang, J.S. Research progress in microalgae resistance to cadmium stress. Microbiol. Chin. 2017, 44, 1980–1987. [Google Scholar]
- George, S.; Venkataraman, G.; Parida, A.A. chloroplast-localized and auxin-induced glutathione S-transferase from phreatophyte Prosopis juliflora confer drought tolerance on tobacco. J. Plant Physiol. 2010, 167, 311–318. [Google Scholar] [CrossRef]
- Nakamura, A.; Umemura, I.; Gomi, K.; Hasegawa, Y.; Kitano, H.; Sazuka, T.; Matsuoka, M. Production and characterization of auxin-insensitive rice by overexpression of a mutagenized rice IAA protein. Plant J. 2006, 46, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.H.; Yang, Y.; Xing, M.H.; Yue, C.P.; Wei, F.; Zhang, Y.J.; Zhao, W.E.; Huang, J.Y. Physiological and transcriptome analyses of Opisthopappus taihangensis in response to drought stress. Cell Biosci. 2019, 9, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, S.; Li, H.; Jiang, Y.; Tang, Y.; Zhao, G.; Zhang, Y.; Yang, S.; Qiu, H.; Wang, Y.; Guo, J.; et al. Transcriptomic Analysis of Female Panicles Reveals Gene Expression Responses to Drought Stress in Maize (Zea mays L.). Agronomy 2020, 10, 313. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020313
Jia S, Li H, Jiang Y, Tang Y, Zhao G, Zhang Y, Yang S, Qiu H, Wang Y, Guo J, et al. Transcriptomic Analysis of Female Panicles Reveals Gene Expression Responses to Drought Stress in Maize (Zea mays L.). Agronomy. 2020; 10(2):313. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020313
Chicago/Turabian StyleJia, Shuangjie, Hongwei Li, Yanping Jiang, Yulou Tang, Guoqiang Zhao, Yinglei Zhang, Shenjiao Yang, Husen Qiu, Yongchao Wang, Jiameng Guo, and et al. 2020. "Transcriptomic Analysis of Female Panicles Reveals Gene Expression Responses to Drought Stress in Maize (Zea mays L.)" Agronomy 10, no. 2: 313. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020313