Application of Directly Brewed Compost Extract Improves Yield and Quality in Baby Leaf Lettuce Grown Hydroponically

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Compost Extract Characteristics and Application
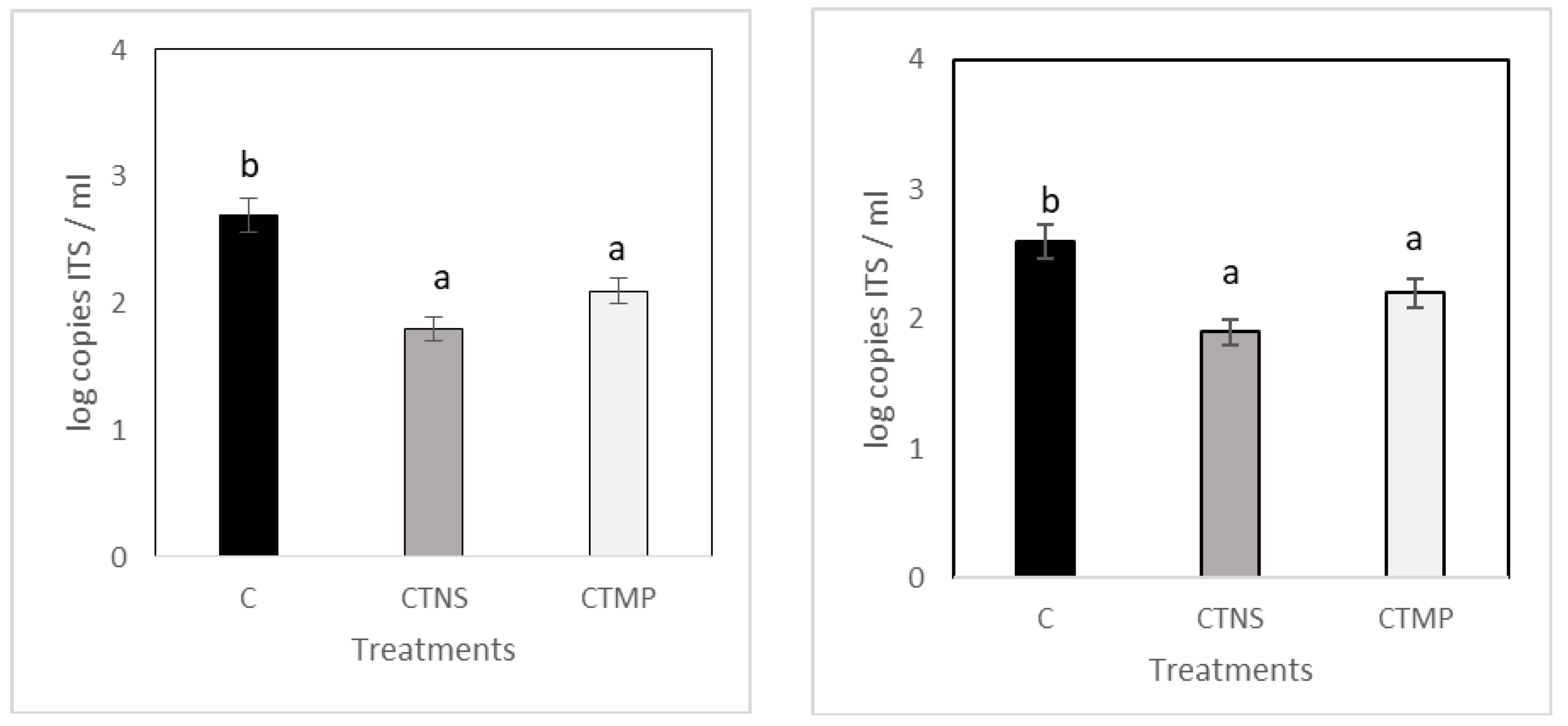
2.3. Pathogen and Inoculation
2.4. Analysis at Harvesting Time
2.5. Postharvest Product Management and Analysis
2.6. Experimental Design and Statistical Analysis
3. Results
3.1. Growth and Yield of Lettuce at Harvesting Time
3.2. Nutritional and Microbiological Quality of Fresh-Cut Product
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Razza, F.; D’Avino, L.; L’Abate, G.; Lazzeri, L. The role of compost in bio-waste management and circular economy. In Designing Sustainable Technologies, Products and Policies; Benetto, E., Gericke, K., Guiton, M., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 133–143. [Google Scholar]
- Bernal-Vicente, A.; Ros, M.; Tittarelli, F.; Intrigliolo, F.; Pascual, J.A. Citrus compost and its water extract for cultivation of melon plants in greenhouse nurseries. Evaluation of nutriactive and biocontrol effects. Bioresour. Technol. 2008, 99, 8722–8728. [Google Scholar] [CrossRef] [PubMed]
- Blaya, J.; Lloret, E.; Ros, M.; Pascual, J.A. Identification of predictor parameters to determine agro-industrial compost suppressiveness against Fusarium oxysporum and Phytophthora capsici diseases in muskmelon and pepper seedlings. J. Sci. Food Agric. 2015, 95, 1482–1490. [Google Scholar] [CrossRef] [PubMed]
- Ronga, D.; Villecco, D.; Zaccardelli, M. Effects of compost and defatted oilseed meals as sustainable organic fertilisers on cardoon (Cynara cardunculus L.) production in the Mediterranean basin. J. Hortic. Sci. Biotechnol. 2019, 94, 664–675. [Google Scholar] [CrossRef]
- Moretti, S.M.L.; Bertoncini, I.B.; Abreu-Junior, C.H. Composting sewage sludge with green waste from tree pruning. Sci. Agric. 2015, 72, 432–439. [Google Scholar] [CrossRef]
- Ros, M.; Hernández, M.T.; García, C.; Bernal, A.; Pascual, J.A. Biopesticide effect on Green compost against fusarium wilt on melon plants. J. Appl. Microbiol. 2015, 98, 845–854. [Google Scholar] [CrossRef]
- Pascual, J.A.; Garcia, C.; Hernandez, T.; Lerma, S.; Lynch, J.M. Effectiveness of municipal waste compost and its humic fraction in suppressing Pythium ultimum. Microb. Ecol. 2002, 44, 59–68. [Google Scholar] [CrossRef]
- Hoitink, H.A.J.; Fahy, P.C. Basis for the control of soilborne plant pathogens with composts. Annu. Rev. Phytopathol. 1986, 24, 93–114. [Google Scholar] [CrossRef]
- Noble, R.; Coventry, E. Suppression of soil-borne plant diseases with composts: A review. Biocontrol Sci. Technol. 2005, 15, 3–20. [Google Scholar] [CrossRef]
- Hoitink, H.A.J.; Grebus, M.E. Composts and the control of plant disease. In Humic Substances in Soil, Peats and Waters: Health and Enviromental Aspects; Hayes, M.H.B., Wilson, W.S., Eds.; Royal Society of Chemistry: Cambrige, UK, 1997; pp. 359–366. [Google Scholar]
- Ronga, D.; Pane, C.; Zaccardelli, M.; Pecchioni, N. Use of spent coffee ground compost in peat-based growing media for the production of basil and tomato potting plants. Commun. Soil Sci. Plant Anal. 2016, 47, 356–368. [Google Scholar] [CrossRef]
- Giménez, A.; Fernández, J.A.; Pascual, J.A.; Ros, M.; López-Serrano, M.; Egea-Gilabert, C. An agroindustrial compost as alternative to peat for production of baby leaf red lettuce in a floating system. Sci. Hortic. 2019, 246, 907–915. [Google Scholar] [CrossRef]
- Giménez, A.; Fernández, J.A.; Egea-Gilabert, C.; Santísima-Trinidad, A.B.; Ros, M.; Pascual, J.A. Agro-industry composts as growing medium for growing baby-leaf lettuces in a floating system—Added-value to suppress Pythium irregulare. Acta Hortic. 2019, 1242, 791–798. [Google Scholar] [CrossRef]
- Zhang, W.; Han, D.Y.; Dick, W.A.; Davis, K.R.; Hoitink, H.A.J. Compost and compost water extract-induced systemic acquired resistance in cucumber and Arabidopsis. Phytopathology 1998, 88, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litterick, A.M.; Harrier, L.; Wallace, P.; Watson, C.A.; Wood, M. The role of uncomposted materials, composts, manures, and compost extracts in reducing pest and disease incidence and severity in sustainable temperate agricultural and horticultural crop production—A review. Crit. Rev. Plant Sci. 2004, 23, 453–479. [Google Scholar] [CrossRef]
- Dafermos, N.G.; Kasselaki, A.M.; Goumas, D.E.; Eyre, M.D.; Spandidakis, K.; Leifert, C. Integration of elicitors and less susceptible hybrids for the control of powdery mildew in organic tomato crops. Plant Dis. 2012, 96, 1506–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giotis, C.; Theodoropoulou, A.; Cooper, J.; Hodgson, R.; Shotton, P.; Shiel, R.; Eyre, M.; Wilcockson, S.; Markellou, E.; Liopa-Tsakalidis, A.; et al. Effect of variety choice, resistant rootstocks and chitin soil amendments on soil-borne diseases in soil-based, protected tomato production systems. Eur. J. Plant Pathol. 2012, 134, 605–617. [Google Scholar] [CrossRef]
- Diver, S.; Greer, L. Sustainable Small-Scale Nursery Production. Horticulturae System Guide. ATTRA. 2001. Available online: http://www.attra.org/attra-pub/PDF/nursery.pdf (accessed on 20 November 2018).
- Eyheraguibel, S.J.; Morard, P. Effects of humic substances derived from organic waste enhancement on the growth and mineral nutrition of maize. Bioresour. Technol. 2008, 99, 4206–4212. [Google Scholar] [CrossRef] [Green Version]
- Arancon, N.Q.; Pant, A.; Radovich, T.; Nguyen, V.H.; Potter, J.; Converse, C. Seed germination and seedling growth of lettuce and tomato as affected by vermicompost water extracts (teas). HortScience 2012, 47, 1722–1728. [Google Scholar] [CrossRef] [Green Version]
- Khaled, H.; Fawy, H.A. Effect of different levels of humic acids on the nutrient content, plant growth, nd soil properties under conditions of salinity. Soil Water Res. 2011, 6, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Morard, P.; Eyheraguibel, B.; Morard, M.; Silvestre, J. Direct effects of humic-like substance on growth, water, and mineral nutrition of various species. J. Plant Nutr. 2011, 34, 46–59. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Muscolo, A.; Vianello, A. Physiological effects of humic substances on higher plants. Soil Biol. Biochem. 2002, 34, 1527–1536. [Google Scholar] [CrossRef]
- St. Martin, C.C.G.; Brathwaite, R.A.I. Compost and compost tea: Principles and prospects as substrates and soil-borne disease management strategies in soil-less vegetable production. Biol. Agric. Hortic. 2012, 28, 1–33. [Google Scholar] [CrossRef]
- Al-Dahmani, J.H.; Abbasi, P.A.; Miller, S.A.; Hoitink, H.A.J. Suppression of bacterial spot of tomato with foliar sprays of compost extracts under greenhouse and field conditions. Plant Dis. 2003, 87, 913–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welke, S.E. The effect of compost extract on the yield of strawberries and the severity of Botrytis cinerea. J. Sustain. Agric. 2005, 25, 57–68. [Google Scholar] [CrossRef]
- Koné, S.B.; Dionne, A.; Tweddell, R.J.; Antoun, H.; Avis, T.J. Suppressive effect of non-aerated compost teas on foliar fungal pathogens of tomato. Biol. Control 2010, 52, 167–173. [Google Scholar] [CrossRef]
- Arancon, N.Q. Suppression of arthropod pests and diseases using vermicompost teas. In Tea Time in the Tropics: A Handbook of Compost Tea Production and Use; Radovich, T., Arancon, N.Q., Eds.; College of Tropical Agriculture and Human Resources, University of Hawaii: Honolulu, HI, USA, 2011; pp. 41–58. [Google Scholar]
- Kim, M.J.; Shim, C.K.; Kim, Y.K.; Hong, S.J.; Park, J.H.; Han, E.J.; Kim, J.H.; Kim, S.C. Effect of aerated compost tea on the growth promotion of lettuce, soybean, and sweet corn in organic cultivation. Plant Pathol. J. 2015, 31, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaccardelli, M.; Pane, C.; Villecco, D.; Palese, A.M.; Celano, G. Compost tea spraying increases yield performance of pepper (Capsicum annuum L.) grown in greenhouse under organic farming system. Ital. J. Agron. 2018, 13, 229–234. [Google Scholar] [CrossRef]
- Radovich, T.; Arancon, N.Q. Tea Time in the Tropics: A Handbook of Compost Tea Production and Use; College of Tropical Agriculture and Human Resources, University of Hawaii: Manoa, HI, USA, 2011. [Google Scholar]
- Egea-Gilabert, C.; Fernández, J.A.; Migliaro, D.; Martínez-Sánchez, J.J.; Vicente, M.J. Genetic variability in wild vs. cultivated Eruca vesicaria populations as assessed by morphological, agronomical and molecular analyses. Sci. Hortic. 2009, 121, 260–266. [Google Scholar] [CrossRef]
- Lara, L.J.; Egea-Gilabert, C.; Niñirola, D.; Conesa, E.; Fernández, J.A. Effect of aeration of the nutrient solution on the growth and quality of purslane (Portulaca oleracea). J. Hortic. Sci. Biotechnol. 2011, 86, 603–610. [Google Scholar] [CrossRef]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.M.; Walker, R.B. Through study of reactivity of various compound classes toward the Folin-Ciocalteu reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Niñirola, D.; Fernández, J.A.; Conesa, E.; Martínez, J.A.; Egea-Gilabert, C. Combined effects of growth cycle and different levels of aeration in nutrient solution on productivity, quality and shelf-life of watercress (Nasturtium officinale R. Br.) plants. HortScience 2014, 49, 567–573. [Google Scholar] [CrossRef]
- Panova, G.G.; Heibner, A.; Grosch, R.; Kläring, H.P. Pythium aphanidermatum may reduce cucumber growth without affecting leaf photosynthesis. J. Phytopathol. 2012, 160, 37–40. [Google Scholar] [CrossRef]
- Trevisan, S.; Francioso, O.; QuaggiottI, S.; Nardi, S. Humic substances biological activity at the plant-soil interface: From environmental aspects to molecular factors. Plant Signal. Behav. 2010, 5, 635–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, D.; Grosch, R. Influence of nutrient solution concentration and a root pathogen (Pythium aphanidermatum) on tomato root growth and morphology. Sci. Hortic. 2003, 97, 109–120. [Google Scholar] [CrossRef]
- Wulff, E.G.; Pham, A.T.H.; Ch´erif, M.; Rey, P.; Tirilly, Y.; Hockenhull, J. Inoculation of cucumber roots with zoospores of mycoparasitic and plant pathogenic Pythium species: Differential zoospore accumulation, colonization ability and plant growth response. Eur. J. Plant Pathol. 1998, 104, 69–76. [Google Scholar] [CrossRef]
- Biernacki, M.; Bruton, B.D. Comparison of leaf-area index with root weight for assessing plant damage by soil-borne pathogens. Acta Hortic. 2000, 510, 163–170. [Google Scholar] [CrossRef]
- Ezz El-Din, A.A.; Hendawy, S.F. Effect of dry yeast and compost tea on growth and oil content of borago officinalis plant. Res. J. Agric. Biol. Sci. 2010, 6, 424–430. [Google Scholar]
- Keeling, A.A.; McCallum, K.R.; Beckwith, C.P. Mature green waste compost enhances growth and nitrogen uptake in wheat (Triticum aestivum L.) and oilseed rape (Brassica napus L.) through the action of water-extractable factors. Bioresour. Technol. 2003, 90, 127–132. [Google Scholar] [CrossRef]
- Khalid, K.A.; Hendawy, S.F.; El-Gezawy, E. Ocimum basilicum L. production under organic farming. Res. J. Agric. Biol. Sci. 2006, 2, 25–32. [Google Scholar]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- European Union. Commission Regulation (EU) n° 1258/2011 of 2 December 2011 Amending Regulation (EC) n° 1881/2006 As Regards Maximum Level for Nitrates in Foodstuffs. Off. J. Eur. Union L 2011, 320, 15–17. [Google Scholar]
- Tsouvaltzis, P.; Koukounaras, A.; Siomos, S.A. Application of amino acids improves lettuce crop uniformity and inhibits nitrate accumulation induced by the supplemental inorganic nitrogen fertilization. Int. J. Agric. Biol. 2014, 16, 951–955. [Google Scholar]
- Colla, G.; Kim, H.J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- Tamme, T.; Reinik, M.; Roasto, M. Nitrates and nitrites in vegetables: Occurrence and health risks. In Bioactive Foods Promoting Health: Fruits and Vegetables; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Salt Lake City, UT, USA, 2009; pp. 307–321. [Google Scholar]
- Burns, I.G.; Zhang, K.; Turner, M.K.; Edmondson, R. Iso-osmotic regulation of nitrate accumulation in lettuce. J. Plant Nutr. 2010, 34, 283–313. [Google Scholar] [CrossRef] [Green Version]
- Gómez, P.; Artés, F.; Madrid, R. Nitrogen fertiliser rate and controlled atmospheres effects on the nitrates levels and quality of fresh processed celery sticks. Acta Hortic. 2003, 604, 493–498. [Google Scholar] [CrossRef]
- Miceli, A.; Vetrano, F.; Sabatino, L.; D’Anna, F.; Moncada, A. Influence of preharvest gibberellic acid treatments on postharvest quality of minimally processed leaf lettuce and rocket. Horticulturae 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Konstantopoulo, E.; Kapotis, E.; Salachas, G.; Petropoulos, S.A.; Karapanos, I.C.; Passam, H.C. Nutritional quality of greenhouse lettuce at harvest and after storage in relation to N application and cultivation season. Sci. Hort. 2010, 125, 93–94. [Google Scholar] [CrossRef]
- Becker, C.; Urlić, B.; Jukić Špika, M.; Kläring, H.P.; Krumbein, A.; Baldermann, S.; Goreta Ban, S.; Perica, S.; Schwarz, D. Nitrogen limited red and green leaf lettuce accumulate flavonoid glycosides, caffeic acid derivatives, and sucrose while losing chlorophylls, β-carotene and xanthophylls. PLoS ONE 2015, 10, e0142867. [Google Scholar] [CrossRef]
- Viršilė, A.; Brazaitytė, A.; Vaštakaitė-Kairienė, V.; Miliauskienė, J.; Jankauskienė, J.; Novičkovas, A.; Samuolienė, G. Lighting intensity and photoperiod serves tailoring nitrate assimilation indices in red and green baby leaf lettuce. J. Sci. Food Agric. 2019, 99, 6608–6619. [Google Scholar] [CrossRef]
- Yahia, E.M.; Gardea-Béjar, A.; Ornelas-Paz, J.J.; Maya-Meraz, O.; Rodríguez-Roque, M.J.; Rios-Velasco, C.; Ornelas-Paz, J.; Salas-Marina, M.A. Preharvest factors affecting postharvest quality. In Postharvest Technology of Perishable Horticultural Commodities; Yahia, E.M., Ed.; Elsevier: Cambridge, UK, 2019; pp. 99–128. [Google Scholar]
- Kołton, A.; Baran, A. Effect of Different Mineral Nitrogen and Compost Nutrition on Some Compounds of Corn Salad (Valerianella Locusta (L.) Latter; Lithuanian Institute of Horticulture; Lithuanian University of Agriculture: Babtai, Kaunas District, Lithuania, 2008; Volume 27, pp. 379–387. [Google Scholar]
- Fiasconaro, M.L.; Lovato, M.E.; Antolín, M.C.; Clementi, L.A.; Torres, N.; Gervasio, S. Role of proline accumulation on fruit quality of pepper (Capsicum annuum L.) grown with a K-rich compost under drought conditions. Sci. Hortic. 2019, 249, 280–288. [Google Scholar] [CrossRef]
- Pant, A.; Radovich, T.J.K.; Hue, N.V.; Arancon, N.Q. Effects of vermicompost tea (aqueous extract) on pak choi yield, quality, and on soil biological properties. Compost Sci. Util. 2011, 19, 279–292. [Google Scholar] [CrossRef]
- Sarwar, M.; Patra, J.K.; Ali, A.; Maqbool, M.; Arshad, M.I. Effect of compost and NPK fertilizer on improving biochemical and antioxidant properties of Moringa oleifera. S. Afr. J. Bot. 2019. [Google Scholar] [CrossRef]
- Haghighi, M. Sewage sludge application in soil improved leafy vegetable growth. J. Biol. Environ. Sci. 2011, 5, 165–167. [Google Scholar]
- Tavarinia, S.; Cardellia, R.; Saviozzia, A.; Degl’Innocentia, E.; Guidia, L. Effects of green compost on soil biochemical characteristics and nutritive quality of leafy vegetables. Compost Sci. Util. 2011, 19, 114–122. [Google Scholar] [CrossRef]
- Marin, A.; Ferreres, F.; Barberá, G.G.; Gil, M.I. Weather variability influences color and phenolic content of pigmented baby leaf lettuces throughout the season. J. Agric. Food Chem. 2015, 63, 1673–1681. [Google Scholar] [CrossRef]
- Oh, M.M.; Carey, E.E.; Rajashekar, C.B. Environmental stresses induce health-promoting phytochemicals in lettuce. Plant Physiol. Biochem. 2009, 47, 578–583. [Google Scholar] [CrossRef]
- Pérez-López, U.; Sgherrib, C.; Miranda-Apodacaa, J.; Micaellib, F.; Lacuestac, M.; Mena-Petitea, A.; Quartaccib, M.F.; Muñoz-Rueda, A. Concentration of phenolic compounds is increased in lettuce grown under high light intensity and elevated CO2. Plant Physiol. Biochem. 2018, 123, 233–241. [Google Scholar] [CrossRef]
- Ouzounis, T.; Parjikolaei, B.R.; Fretté, X.; Rosenqvist, E.; Ottosen, C.O. Predawn and high intensity application of supplemental blue light decreases the quantum yield of PSII and enhances the amount of phenolic acids, flavonoids, and pigments in Lactuca sativa. Front. Plant Sci. 2015, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kalt, W.; Forney, C.F.; Martin, A.; Prior, R.L. Antioxidant capacity, vitamin C, phenolics, and anthocyanins after fresh storage of small fruits. J. Agric. Food Chem. 1999, 47, 4638–4644. [Google Scholar] [CrossRef]
- Serafini, S.; Bugianesi, R.; Salucci, M.; Azzini, E.; Raguzzini, A.; Maiani, G. Effect of acute ingestion of fresh and stored lettuce (Lactuca sativa) on plasma total antioxidant capacity and antioxidant levels in human subjects. Br. J. Nutr. 2002, 88, 615–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninfali, P.; Mea, G.; Giorgini, S.; Rocchi, M.; Bacchiocca, M. Antioxidant capacity of vegetables, spices and dressings relevant to nutrition. Br. J. Nutr. 2005, 93, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, R.; Sultana, S.; Rejaul Islam, M.D.; Monda, C. Effect of aerated and non-aerated compost tea against some fungal phytopathogens. J. Bangladesh Agric. Univ. 2019, 17, 142–147. [Google Scholar] [CrossRef]
- Gil, M.I.; Ferreres, F.; Tomás-Barberá, F.T. Effect of postharvest storage and processing on the antioxidant constituents (flavonoids and vitamin C) of fresh-cut spinach. J. Agric. Food Chem. 1999, 47, 2213–2217. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Iwamoto, T.; Carey, E.E. Antioxidant capacity of leafy vegetables as affected by high tunnel environment, fertilisation and growth stage. J. Sci. Food Agric. 2007, 87, 2692–2699. [Google Scholar] [CrossRef]
- DuPont, M.S.; Mondin, Z.; Williamson, G.; Price, K.R. Effect of variety, processing, and storage on the flavonoid glycoside content and composition of lettuce and endive. J. Agric. Food Chem. 2000, 48, 3957–3964. [Google Scholar] [CrossRef]
- Selma, M.V.; Luna, M.C.; Martínez-Sánchez, A.; Tudela, J.A.; Beltrán, D.; Baixauli, C.; Gil, M.I. Sensory quality, bioactive constituents and microbiological quality of green and red fresh-cut lettuces (Lactuca sativa L.) are influenced by soil and soilless agricultural production systems. Postharvest Biol. Technol. 2012, 63, 16–24. [Google Scholar] [CrossRef]


{kind=link}
| NO3− (mg L−1) | SO42− (mg L−1) | Cl− (mg L−1) | Na+ (mg L−1) | K+ (mg L−1) | Mg2+ (mg L−1) | Ca2+ (mg L−1) | |
|---|---|---|---|---|---|---|---|
| CENS | 13.6 | 143.2 | 170.6 | 109.4 | 26.9 | 30.3 | 77.2 |
| CEMP | 13.4 | 143.5 | 176.1 | 115.6 | 21.9 | 30.0 | 77.0 |
| Fresh Biomass (g Plant−1) | Dry Matter (%) | SLA (m2 kg−1) | Total Root Length (cm) | Root Diameter (mm) | Length of 0 to 0.5 mm Diam. Root (cm) | |
|---|---|---|---|---|---|---|
| Inoculation (A) | ||||||
| NI | 2.16 ± 0.04 b | 3.76 ± 0.07 b | 81.08 ± 0.73 b | 307.61 ± 5.30 b | 0.54 ± 0.01 a | 248.59 ± 4.22 b |
| I | 1.98 ± 0.04 a | 4.19 ± 0.08 a | 75.06 ± 0.82 a | 176.57 ± 2.97 a | 0.64 ± 0.01 b | 137.73 ± 2.77 a |
| Application (B) | ||||||
| C | 2.00 ± 0.05 a | 4.14 ± 0.10 b | 80.27 ± 1.40 b | 232.28 ± 8.09 b | 0.69 ± 0.01 b | 186.19 ± 6.65 b |
| CENS | 2.22 ± 0.05 b | 4.04 ± 0.10 b | 75.07 ± 0.61 a | 289.65 ± 4.31 c | 0.56 ± 0.01 a | 239.43 ± 6.65 c |
| CEMP | 2.00 ± 0.04 a | 3.76 ± 0.08 a | 78.87 ± 0.72ab | 204.36 ± 8.41 a | 0.54 ± 0.01 a | 153.86 ± 3.54 a |
| A × B | ||||||
| NI × C | 2.05 ± 0.07 | 3.60 ± 0.13 a | 90.36 ± 1.26 d | 311.96 ± 7.08 d | 0.66 ± 0.01 d | 256.73 ± 3.61 c |
| NI × CENS | 2.32 ± 0.07 | 3.62 ± 0.10 a | 80.34 ± 0.70 b | 368.81 ± 7.85 e | 0.46 ± 0.02 a | 300.65 ± 6.32 d |
| NI × CEMP | 2.12 ± 0.06 | 3.45 ± 0.10 a | 85.18 ± 0.71 c | 242.06 ± 3.94 c | 0.53 ± 0.01 b | 188.40 ± 5.37 b |
| I × C | 1.96 ± 0.07 | 4.68 ± 0.14 c | 72.55 ± 0.56 a | 152.59 ± 3.64 a | 0.72 ± 0.02 e | 115.65 ± 2.68 a |
| I × CENS | 2.12 ± 0.08 | 4.46 ± 0.15 c | 70.19 ± 1.76 a | 211.49 ± 4.98 b | 0.64 ± 0.01 d | 178.21 ± 4.37 b |
| I × CEMP | 1.88 ± 0.05 | 4.07 ± 0.12 b | 69.79 ± 0.34 a | 166.65 ± 3.76 a | 0.59 ± 0.01 c | 119.31 ± 2.21 a |
| Significance | ||||||
| Inoculation (A) | ** | *** | *** | *** | *** | *** |
| Application (B) | ** | *** | *** | *** | *** | *** |
| A × B | n.s. | ** | *** | *** | *** | *** |
| Fresh Biomass (g Plant−1) | % Dry Matter | SLA (m2 kg−1) | Total Root Length (cm) | Root Diameter (mm) | Length of 0 to 0.5 mm Diam. Root (cm) | |
|---|---|---|---|---|---|---|
| Inoculation (A) | ||||||
| NI | 1.49 ± 0.02 | 4.43 ± 0.07 | 76.40 ± 0.51 | 164.24 ± 2.74 b | 0.29 ± 0.01 a | 138.79 ± 1.92 b |
| I | 1.45 ± 0.03 | 4.41 ± 0.08 | 76.20 ± 0.66 | 148.62 ± 2.13 a | 0.32 ± 0.01 b | 132.01 ± 1.85 a |
| Application (B) | ||||||
| C | 1.49 ± 0.03 b | 4.30 ± 0.10 | 79.05 ± 1.66 b | 156.07 ± 2.70 b | 0.28 ± 0.01 a | 136.40 ± 2.31 b |
| CENS | 1.52 ± 0.03 b | 4.45 ± 0.10 | 74.94 ± 1.79 a | 176.13 ± 3.21 c | 0.31 ± 0.01 b | 148.58 ± 2.04 c |
| CEMP | 1.39 ± 0.03 a | 4.49 ± 0.08 | 74.91 ± 1.07 a | 138.60 ± 2.37 a | 0.32 ± 0.01 b | 121.22 ± 1.97 a |
| A × B | ||||||
| NI × C | 1.52 ± 0.04 | 4.26 ± 0.13 | 79.40 ± 0.82 b | 164.66 ± 4.01 c | 0.24 ± 0.01 a | 143.58 ± 3.30 |
| NI × CENS | 1.55 ± 0.05 | 4.39 ± 0.13 | 76.31 ± 0.99 ab | 190.63 ± 4.51 d | 0.32 ± 0.01 bc | 149.20 ± 2.85 |
| NI × CEMP | 1.40 ± 0.04 | 4.62 ± 0.13 | 76.33 ± 0.53 b | 140.44 ± 3.48 ab | 0.30 ± 0.01 b | 123.60 ± 2.97 |
| I × C | 1.47 ± 0.05 | 4.34 ± 0.15 | 78.71 ± 2.76 b | 147.48 ± 3.30 b | 0.31 ± 0.01 bc | 129.23 ± 2.45 |
| I × CENS | 1.50 ± 0.05 | 4.51 ± 0.15 | 73.57 ± 1.54 a | 161.62 ± 3.83 c | 0.33 ± 0.01 c | 147.97 ± 2.74 |
| I × CEMP | 1.39 ± 0.04 | 4.37 ± 0.11 | 73.48 ± 0.68 a | 136.77 ± 3.27 a | 0.33 ± 0.01 c | 118.83 ± 2.79 |
| Significance | ||||||
| Inoculation (A) | n.s. | n.s. | n.s. | *** | *** | ** |
| Application (B) | ** | n.s. | ** | *** | ** | *** |
| A × B | n.s. | n.s. | ** | *** | *** | n.s. |
| Nitrate (mg kg−1 FW) | Total Phenolics (mg GA kg−1 FW) | Total Flavonoids (mg Rutin kg−1 FW) | Antioxidant Capacity (mg DPPHreduced kg−1 FW) | Mesophilic Microorganisms (Log CFU g−1) | Psychrophilic Microorganisms (Log CFU g−1) | |
|---|---|---|---|---|---|---|
| Application (A) | ||||||
| C | 1379.76 ± 74.91 b | 1044.02 ± 151.38 a | 1912.56 ± 162.07 a | 105.32 ± 9.57 a | 3.57 ± 0.24 | 3.74 ± 0.38 b |
| CENS | 1290.69 ± 71.72 b | 1168.74 ± 137.57 b | 2691.93 ± 92.71 b | 120.84 ± 7.49 b | 3.48 ± 0.25 | 3.59 ± 0.64 a |
| CEMP | 1079.91 ± 61.10 a | 1280.77 ± 131.99 c | 2917.70 ± 80.25 b | 141.93 ± 12.74 c | 3.59 ± 0.27 | 3.99 ± 0.37 c |
| Storage (B) | ||||||
| 0 days | 1491.93 ± 39.58 b | 1727.93 ± 41.73 b | 2872.28 ± 77.43 b | 159.37 ± 6.39 b | 2.74 ± 0.05 a | 2.23 ± 0.13 a |
| 7 days | 1007.27 ± 38.22 a | 604.09 ± 25.58 a | 2142.51 ± 126.60 a | 86.02 ± 12.88 a | 4.35 ± 0.09 b | 5.32 ± 0.07 b |
| A × B | ||||||
| C × 0 days | 1636.46 ± 49.72 | 1636.45 ± 98.01 | 2550.72 ± 98.89 bc | 141.22 ± 7.71 c | 2.88 ± 0.06 | 2.41 ± 0.02 b |
| CENS × 0 days | 1582.35 ± 58.61 | 1717.41 ± 67.29 | 3009.85 ± 88.43 de | 148.76 ± 6.47 c | 2.67 ± 0.12 | 1.47 ± 0.003 a |
| CEMP × 0 days | 1300.96 ± 43.94 | 1820.92 ± 26.62 | 3056.26 ± 149.35 e | 188.14 ± 11.73 d | 2.68 ± 0.03 | 2.76 ± 0.02 c |
| C × 7 days | 1123.06 ± 70.01 | 451.60 ± 6.63 | 1274.39 ± 31.76 a | 69.42 ± 2.84 a | 4.25 ± 0.26 | 5.03 ± 0.005 d |
| CENS × 7 days | 1043.04 ± 55.61 | 620.06 ± 25.37 | 2374.01 ± 58.67 b | 92.93 ± 1.46 b | 4.29 ± 0.07 | 5.71 ± 0.03 f |
| CEMP × 7 days | 855.70 ± 39.27 | 740.62 ± 20.03 | 2779.13 ± 16.20 cd | 95.73 ± 4.34 b | 4.51 ± 0.06 | 5.21 ± 0.04 e |
| Significance | ||||||
| Application (A) | *** | *** | *** | *** | n.s. | ** |
| Storage (B) | *** | *** | *** | *** | *** | *** |
| A × B | n.s. | n.s. | *** | * | n.s. | *** |
| Nitrate (mg kg−1 FW) | Total Phenolics (mg GA kg−1 FW) | Total Flavonoids (mg Rutin kg−1 FW) | Antioxidant Capacity (mg DPPHreduced kg−1 FW) | Mesophilic Microorganisms (Log CFU g−1) | Psychrophilic Microorganisms (Log CFU g−1) | |
|---|---|---|---|---|---|---|
| Application (A) | ||||||
| C | 2015.92 ± 135.74 c | 597.96 ± 32.85 a | 1316.82 ± 26.75 a | 90.95 ± 2.83 a | 3.56 ± 0.35 b | 3.22 ± 0.52 a |
| CENS | 1812.72 ± 130.42 b | 841.05 ± 78.45 b | 1496.79 ± 22.36 b | 117.31 ± 3.20 b | 3.24 ± 0.13 a | 2.96 ± 0.33 a |
| CEMP | 1463.84 ± 106.93 a | 1005.97 ± 98.33 c | 1594.37 ± 13.94 c | 138.27 ± 2.98 c | 3.32 ± 0.18 ab | 4.08 ± 0.49 b |
| Storage (B) | ||||||
| 0 days | 2331.93 ± 90.41 b | 1093.75 ± 56.79 b | 1514.17 ± 26.52 b | 126.26 ± 4.08 b | 2.71 ± 0.08 a | 2.08 ± 0.10 a |
| 7 days | 1196.39 ± 26.52 a | 536.23 ± 18.02 a | 1424.48 ± 27.99 a | 104.75 ± 3.88 a | 4.04 ± 0.14 b | 4.76 ± 0.27 b |
| A × B | ||||||
| C × 0 days | 2807.25 ± 103.84 e | 727.74 ± 11.56 c | 1372.11 ± 20.45 | 100.43 ± 2.83 | 2.39 ± 0.02 a | 1.85 ± 0.11 |
| CENS × 0 days | 2337.72 ± 37.09 d | 1156.81 ± 17.69 d | 1535.90 ± 40.69 | 130.28 ± 0.71 | 2.94 ± 0.14 b | 1.94 ± 0.20 |
| CEMP × 0 days | 1850.82 ± 101.76 c | 1396.71 ± 47.84 e | 1634.51 ± 17.01 | 148.10 ± 3.06 | 2.80 ± 0.09 b | 2.45 ± 0.02 |
| C × 7 days | 1224.58 ± 29.40 ab | 468.17 ± 15.56 a | 1261.53 ± 43.12 | 81.46 ± 1.85 | 4.72 ± 0.14 d | 4.59 ± 0.64 |
| CENS × 7 days | 1287.71 ± 44.81 b | 525.29 ± 30.28 a | 1457.68 ± 9.31 | 104.36 ± 1.11 | 3.55 ± 0.12 c | 3.99 ± 0.13 |
| CEMP × 7 days | 1076.87 ± 28.29 a | 615.22 ± 25.15 b | 1554.24 ± 11.56 | 128.44 ± 2.07 | 3.85 ± 0.16 c | 5.71 ± 0.03 |
| Significance | ||||||
| Application (A) | *** | *** | *** | *** | * | ** |
| Storage (B) | *** | *** | *** | *** | *** | *** |
| A × B | *** | *** | n.s. | n.s. | *** | n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giménez, A.; Fernández, J.A.; Pascual, J.A.; Ros, M.; Egea-Gilabert, C. Application of Directly Brewed Compost Extract Improves Yield and Quality in Baby Leaf Lettuce Grown Hydroponically. Agronomy 2020, 10, 370. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030370
Giménez A, Fernández JA, Pascual JA, Ros M, Egea-Gilabert C. Application of Directly Brewed Compost Extract Improves Yield and Quality in Baby Leaf Lettuce Grown Hydroponically. Agronomy. 2020; 10(3):370. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030370
Chicago/Turabian StyleGiménez, Almudena, Juan A. Fernández, José A. Pascual, Margarita Ros, and Catalina Egea-Gilabert. 2020. "Application of Directly Brewed Compost Extract Improves Yield and Quality in Baby Leaf Lettuce Grown Hydroponically" Agronomy 10, no. 3: 370. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030370