Effect of Salinity Stress on Phenylpropanoid Genes Expression and Related Gene Expression in Wheat Sprout

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Extraction and cDNA Synthesis
2.3. Determination of Phenylpropanoid Biosynthetic Pathway Gene Expression
2.4. Analysis of Phenylpropanoid Content by HPLC
2.5. Statistical Analysis
3. Results and Discussion
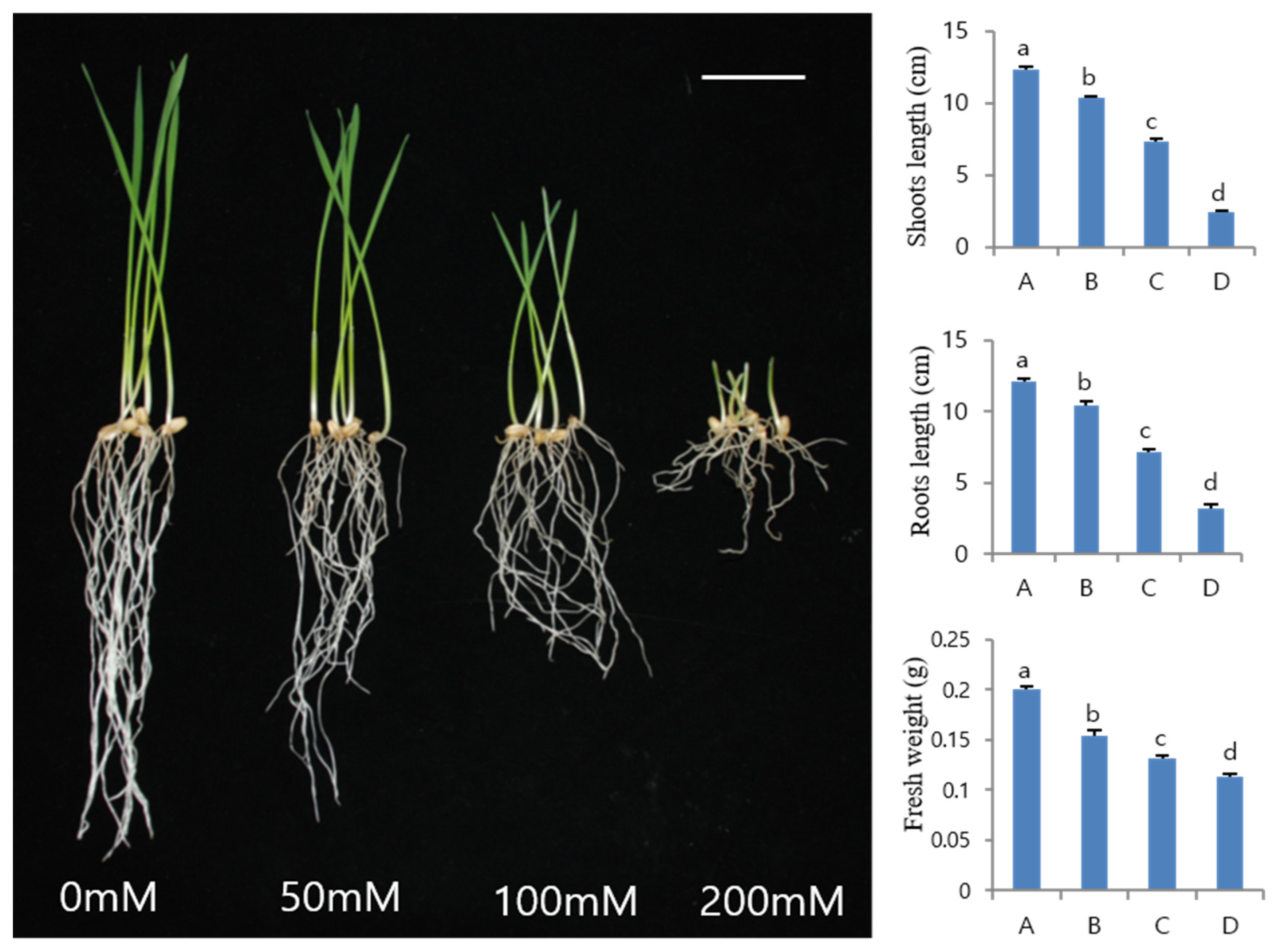
3.1. Effect of NaCl Treatment on the Growth of Wheat Sprouts
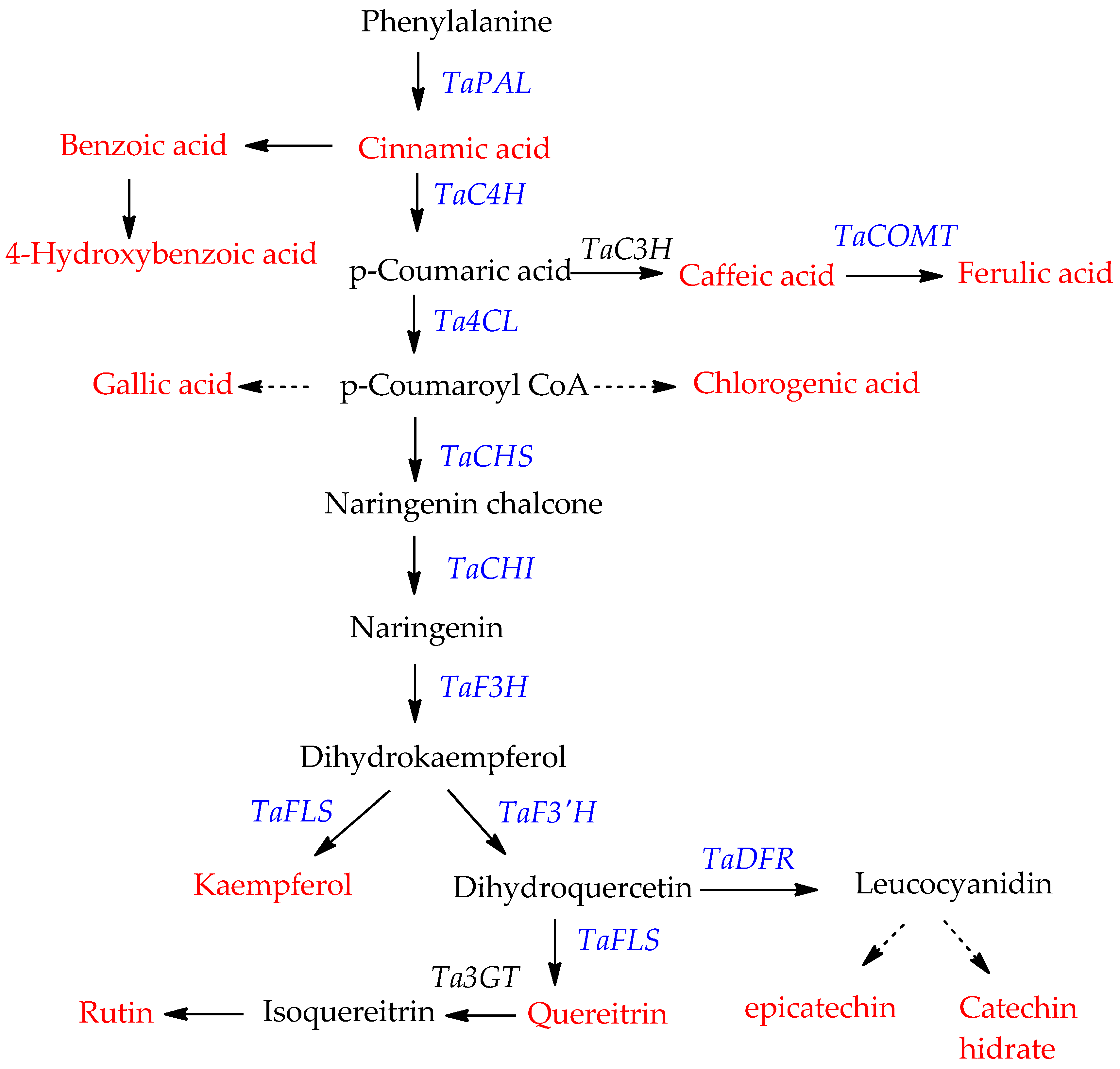
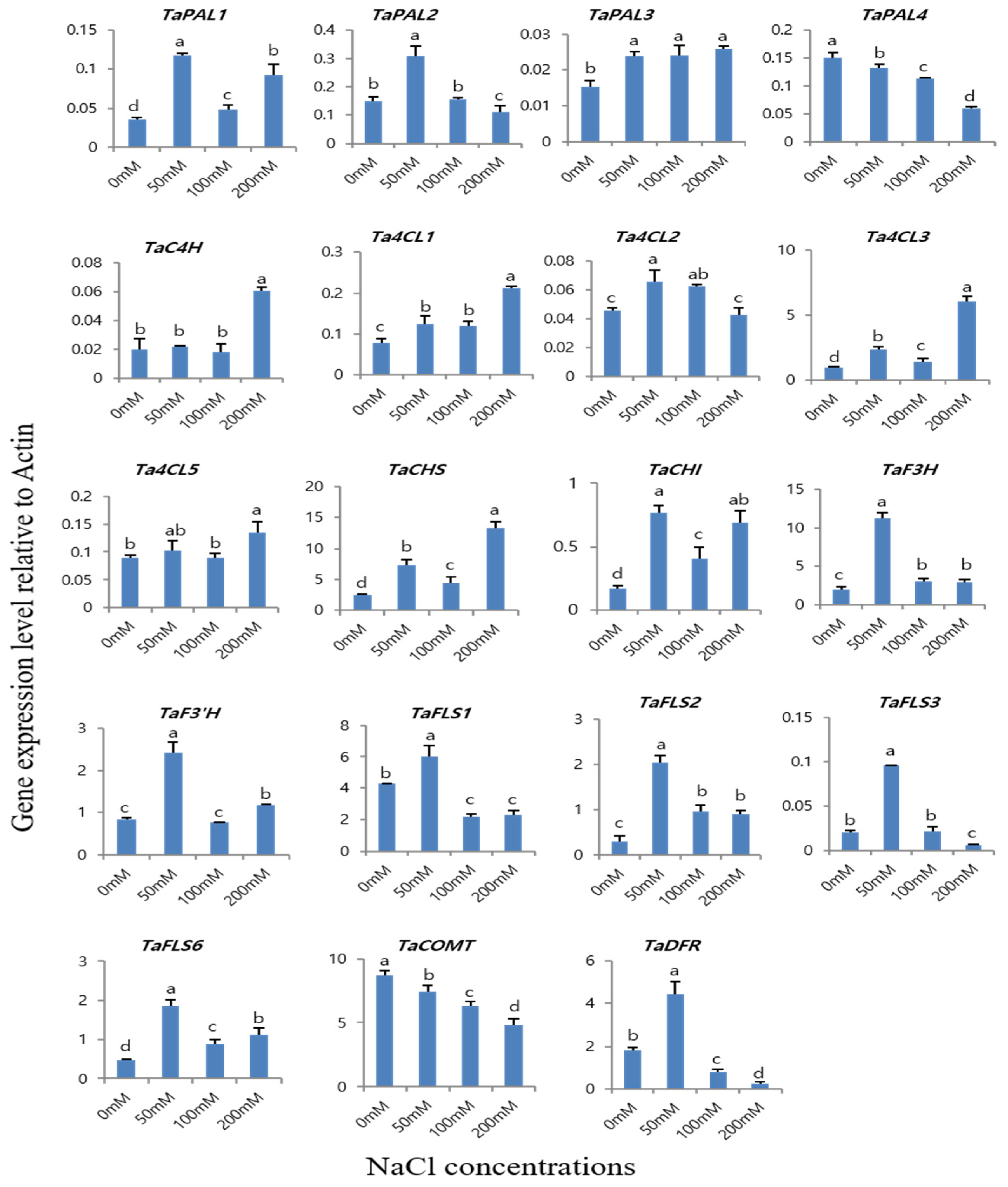
3.2. Effect of NaCl on Genes Expression of Phenylpropanoid Biosynthetic Pathway
3.3. Phenylpropanoid Content under Different NaCl Concentrations
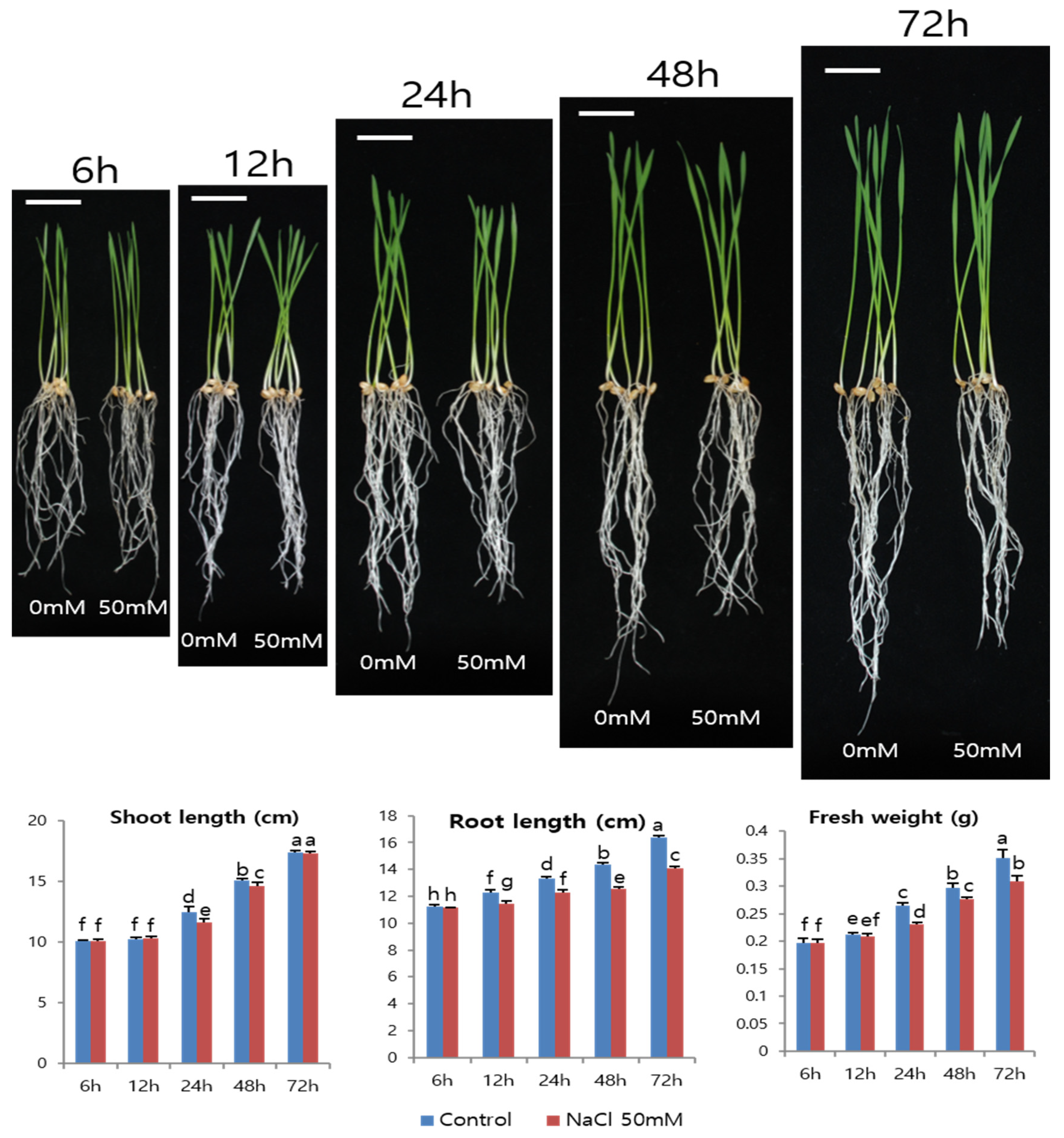
3.4. Wheat Plantlet Growth under 50 mM NaCl Treatment
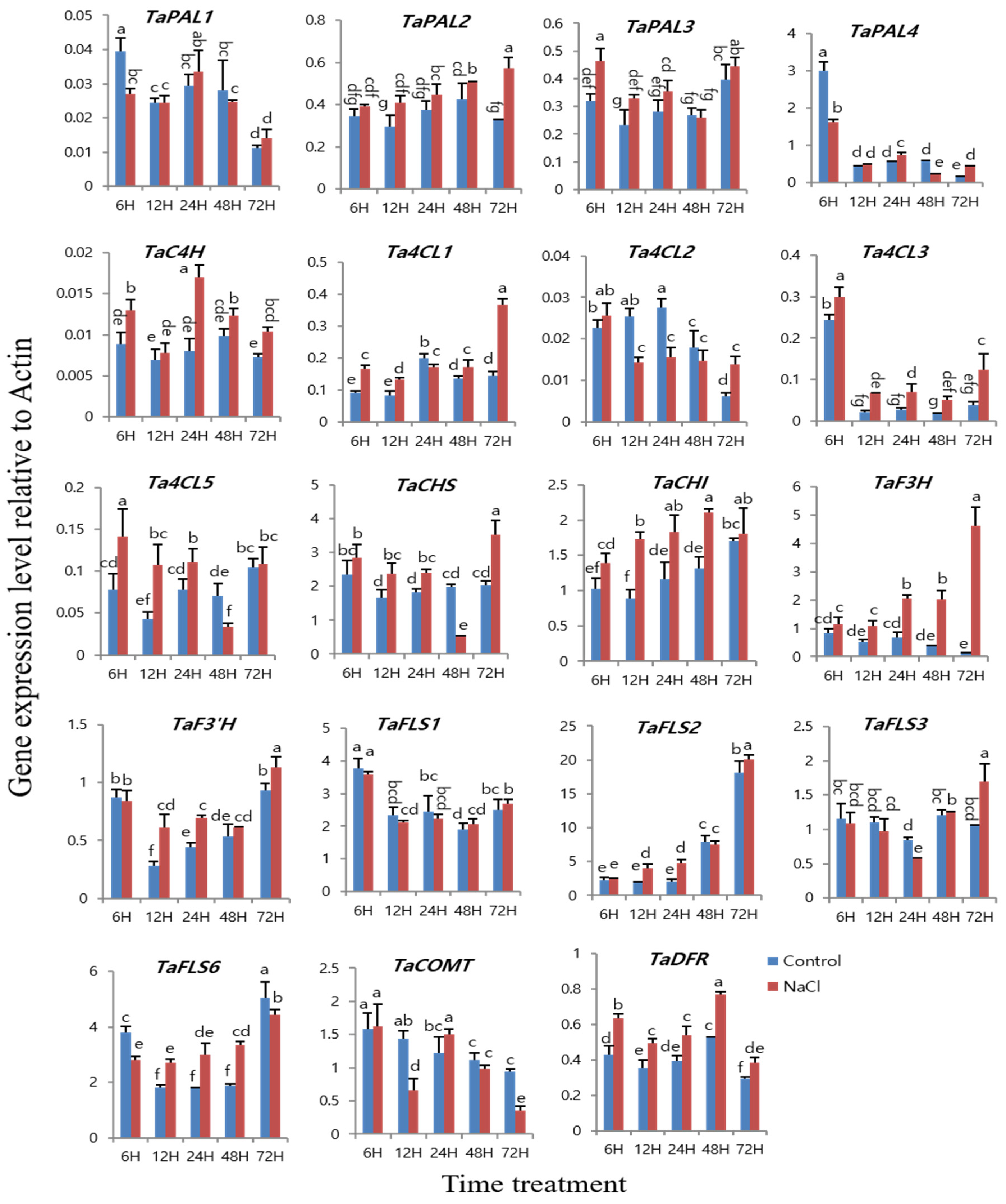
3.5. Expression of Phenylpropanoid Biosynthetic Genes in Six-day-old Wheat Plantlets under 50 mM NaCl Treatment
3.6. Phenylpropanoid Content in Six-day-old Wheat Plantlets after 50 mM NaCl Treatment for 6, 12, 24, 48 and 72 h
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Shannon, M.C. Adaptation of plants to salinity. Adv. Agron. 1997, 60, 75–120. [Google Scholar]
- Gupta, B.; Huang, B.R. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Katerji, N.; van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M.; Karam, F. Salinity and drought, a comparison of their effects on the relationship between yield and evapotranspiration. Agric. Water Manag. 1998, 36, 45–54. [Google Scholar] [CrossRef]
- Hu, Y.C.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Wahid, A.; Ghazanfar, A. Possible involvement of some secondary metabolites in salt tolerance of sugarcane. J. Plant Physiol. 2006, 163, 723–730. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, T.T.; Rowshan, V.; Niazi, A.; Moghadam, A. Effect of salt stress on terpenoid biosynthesis in Salvia mirzayanii: From gene to metabolite. J. Hortic. Sci. Biotech. 2019, 94, 389–399. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterisation of polyphenolic compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Impact of germination on phenolic content and antioxidant activity of 13 edible seed species. Food Chem. 2010, 119, 1485–1490. [Google Scholar] [CrossRef]
- Panda, P.; Appalashetti, M.; Judeh, Z.M.A. Phenylpropanoid Sucrose Esters: Plant-Derived Natural Products as Potential Leads for New Therapeutics. Curr. Med. Chem. 2011, 18, 3234–3251. [Google Scholar] [CrossRef]
- Korkina, L.; Kostyuk, V.; De Luca, C.; Pastore, S. Plant Phenylpropanoids as Emerging Anti-Inflammatory Agents. Mini Rev. Med. Chem. 2011, 11, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Landau, J.M.; Huang, M.T.; Newmark, H.L. Inhibition of carcinogenesis by dietary polyphenolic compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomas-Barberan, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Food Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Eudes, A.; Liang, Y.; Mitra, P.; Loque, D. Lignin bioengineering. Curr. Opin. Biotechnol. 2014, 26, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Naoumkina, M.A.; Zhao, Q.; Gallego-Giraldo, L.; Dai, X.; Zhao, P.X.; Dixon, R.A. Genome-wide analysis of phenylpropanoid defence pathways. Mol. Plant Pathol. 2010, 11, 829–846. [Google Scholar] [CrossRef]
- Liu, J.Y.; Osbourn, A.; Ma, P.D. MYB Transcription Factors as Regulators of Phenylpropanoid Metabolism in Plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [Green Version]
- Noreen, Z.; Ashraf, M. Assessment of variation in antioxidative defense system in salt-treated pea (Pisum sativum) cultivars and its putative use as salinity tolerance markers. J. Plant Physiol. 2009, 166, 1764–1774. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Park, K.J.; Kim, B.K.; Jeong, J.W.; Kim, H.J. Effect of salinity stress on phenolic compounds and carotenoids in buckwheat (Fagopyrum esculentum M.) sprout. Food Chem. 2012, 135, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Fonseca, J.M.; Choi, J.H.; Kubota, C.; Kwon, D.Y. Salt in irrigation water affects the nutritional and visual properties of romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef] [PubMed]
- Hichem, H.; Mounir, D.; Naceur, E. Differential responses of two maize (Zea mays L.) varieties to salt stress: Changes on polyphenols composition of foliage and oxidative damages. Ind. Crop. Prod. 2009, 30, 144–151. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Cuong, D.M.; Kwon, S.J.; Jeon, J.; Park, Y.J.; Park, J.S.; Park, S.U. Identification and Characterization of Phenylpropanoid Biosynthetic Genes and Their Accumulation in Bitter Melon (Momordica charantia). Molecules 2018, 23, 469. [Google Scholar] [CrossRef] [Green Version]
- Koca, H.; Bor, M.; Ozdemir, F.; Turkan, I. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot. 2007, 60, 344–351. [Google Scholar] [CrossRef]
- Saha, P.; Chatterjee, P.; Biswas, A.K. NaCl pretreatment alleviates salt stress by enhancement of antioxidant defense system and osmolyte accumulation in mungbean (Vigna radiata L. Wilczek). Indian J. Exp. Biol. 2010, 48, 593–600. [Google Scholar]
- Agarwal, S.; Pandey, V. Antioxidant enzyme responses to NaCl stress in Cassia angustifolia. Biol. Plant. 2004, 48, 555–560. [Google Scholar] [CrossRef]
- Jamil, M.; Bae, D.L.; Yong, J.K.; Ashraf, M.; Chun, L.S.; Shik, R.E. Effect of salt (NaCl) stress on germination and early seedling growth of four vegetables species. J Cent. Eur. Agric. 2006, 7, 273–282. [Google Scholar]
- Bohnert, H.J.; Jensen, R.G. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol. 1996, 14, 89–97. [Google Scholar] [CrossRef]
- Rossi, L.; Borghi, M.; Francini, A.; Lin, X.; Xie, D.Y.; Sebastiani, L. Salt stress induces differential regulation of the phenylpropanoid pathway in Olea europaea cultivars Frantoio (salt-tolerant) and Leccino (salt-sensitive). J. Plant Physiol. 2016, 204, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Irani, S.; Todd, C.D.; Wei, Y.; Bonham-Smith, P.C. Changes in phenylpropanoid pathway gene expression in roots and leaves of susceptible and resistant Brassica napus lines in response to Plasmodiophora brassicae inoculation. Physiol. Mol. Plant Pathol. 2019, 106, 196–203. [Google Scholar] [CrossRef]
- Ni, J.; Hao, J.; Jiang, Z.; Zhan, X.; Dong, L.; Yang, X.; Sun, Z.; Xu, W.; Wang, Z.; Xu, M. NaCl induces flavonoid biosynthesis through a putative novel pathway in post-harvest ginkgo leaves. Front. Plant Sci. 2017, 8, 920. [Google Scholar] [CrossRef] [Green Version]
- Ben Abdallah, S.; Aung, B.; Amyot, L.; Lalin, I.; Lachaal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant. 2016, 38, 72. [Google Scholar] [CrossRef]
- Navarro, J.M.; Flores, P.; Garrido, C.; Martinez, V. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006, 96, 66–73. [Google Scholar] [CrossRef]
- Lopez-Berenguer, C.; Martinez-Ballesta, M.D.; Moreno, D.A.; Carvajal, M.; Garcia-Viguera, C. Growing hardier crops for better health: Salinity tolerance and the nutritional value of Broccoli. J. Agric. Food Chem. 2009, 57, 572–578. [Google Scholar] [CrossRef]
- Grieve, C.M.; Francois, L.E.; Poss, J.A. Effect of salt stress during early seedling growth on phenology and yield of spring wheat. Cereal Res. Commun. 2001, 29, 167–174. [Google Scholar] [CrossRef]
- Pandey, M.; Penna, S. Time course of physiological, biochemical, and gene expression changes under short-term salt stress in Brassica juncea L. Crop. J. 2017, 5, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Tang, N.; Huang, L.; Zhao, Y.; Tang, X.; Wang, K. Effects of salt stress on plant growth, antioxidant capacity, glandular trichome density, and volatile exudates of Schizonepeta tenuifolia Briq. Int. J. Mol. Sci. 2018, 19, 252. [Google Scholar] [CrossRef] [Green Version]
- Yun, Y.; Jung, H.J.; Rahim, M.A.; Park, N.K.; Kuk, Y. Molecular analysis of genes related to phenylpropanoid and ascorbate biosynthesis in salt and UV-B treated pak choi grown under LEDs. Botany 2019, 97, 513–519. [Google Scholar] [CrossRef]
- Kim, N.S.; Kwon, S.J.; Cuong, D.M.; Jeon, J.; Park, J.S.; Park, S.U. Accumulation of phenylpropanoids in tartary buckwheat (Fagopyrum tataricum) under salt stress. Agronomy 2019, 9, 739. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ID | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|
| TaPAL1 | CJ628388.1 | GCTCTTTGAGGCCAATGTTCTT | GCTTCTATCTGTCCAGGGTGGT |
| TaPAL2 | CJ707202.1 | CTACATGGCACTCGCAAAGAAG | GTTGATCTCACGCTCGATTGAC |
| TaPAL3 | CJ805150.1 | TACATGAAGCAGGCAAAGAAGC | TTGTCGTTGACGGAGTTGATCT |
| TaPAL4 | BQ752712.1 | GTGAACTCTCTGGGCCTTGTGT | TCACGGAGGTCTTGATGTTCTC |
| TaC4H | HX132432.1 | CAACCACCGCAACCTCAT | GGTGAAGATGTCGAAGACGA |
| Ta4CL1 | GH727954.1 | GCCGCTGTTCCACATCTA | GGTCGCTCTTGGCGATCT |
| Ta4CL2 | CJ951387.1 | GAGGCCACAAAGAACACCAT | TTTGATTTCGGCGTGTGTAA |
| Ta4CL3 | CK163034.1 | AGTTCGCCTTCTCCTTCCTC | CGTACTCCAGCACCTTGTCC |
| Ta4CL5 | GH724596.1 | CAAGGGCGTCATGCTCA | AGCGAGTAGATGTGGAACAGC |
| TaCHS | HX091886.1 | CGTGGACGAAGTGATGAAGA | TTAGGTGTTCGCTGTTGGTG |
| TaCHI | CD890742.1 | ACAAGGTGACGGAGAACTGC | GAGTGGGTGAAGAGGATGGA |
| TaF3H | CJ659795.1 | CTGGAGAAGGTGATGGCTGT | CAGATAGTCCCGCCAGTTGT |
| TaF3′H | HX255892.1 | AAGGAGAGGCGCAATAGGAT | GTGATGGGGAAGCTACAAGC |
| TaFLS1 | BE423889.1 | TGAAGGATTTGGCTACTGTGG | GAGAAGACGCGGATGTCGT |
| TaFSL2 | BQ244276.1 | ATCCAAACTGACACGCATGA | AGTTCCCGGCCAAGTACAAG |
| TaFLS3 | HX107493.1 | CTACGGCTTCTTCCAGATCG | CAGATAGTCCCGCCAGTTGT |
| TaFLS6 | CD454732.1 | GTACCAGCATCCGTCCTTGA | GTGGTGCTCCTCCAGAAGAT |
| TaDFR | CJ714375.1 | TACGACCAGGACAACTGGAG | GGGATGATGCTGATGAAGTC |
| TaCOMT1 | CJ858964.1 | CCATCAAGGGCATCAACTTC | CAAGGCGTCGTAGCAGTTCT |
| TaActin | GQ339780 | CGTGTTGGATTCTGGTGATG | CGAGCTTCTCCTTGATGTCC |
| Phenylpropanoid Compound | 0 mM NaCl | 50 mM NaCl | 100 mM NaCl | 200 mM NaCl |
|---|---|---|---|---|
| 4-hydroxybenzoic acid | ND | 22.59 ± 0.55 a | 9.81 ± 0.06 b | ND |
| Catechin hydrate | 65.41 ± 4.26 a | 2.31 ± 0.03 b | 2.65 ± 0.09 b | ND |
| Chlorogenic acid | ND | ND | ND | 44.59 ± 0.49 a |
| Caffeic acid | 8.61 ± 0.16 c | 10.11 ± 0.34 b | 11.62 ± 0.14 a | 6.99 ± 0.09 c |
| Epicatechin | 617.15 ± 26.58 c | 2708.96 ± 227.11 a | 1323.13 ± 29.72 b | 1218.85 ± 29.45 b |
| Ferulic acid | 30.63 ± 1.91 c | 53.75 ± 1.6 a | 36.3 ± 2.53 b | 28.37 ± 1.31 c |
| Benzoic acid | 1604.61 ± 65.0 a | 510.14 ± 12.26 c | 739.53 ± 6.28 b | 191.62 ± 0.51 d |
| Rutin | 3.99 ± 0.61 a | 3.76 ± 0.37 a | 4.01 ± 0.8 a | 3.33 ± 0.47 a |
| Trans-cinnamic acid | 0.57 ± 0.27 b | 1.12 ± 0.15 a | 0.64 ± 0.2 b | 0.52 ± 0.19 b |
| Quercetin | 35.7 ± 4.93 a | 23.37 ± 3.49 b | 5.07 ± 1.64 c | 6.78 ± 3.65 c |
| Kaempferol | 7.06 ± 0.63 a | 6.77 ± 0.88 a | 5.01 ± 0.54 b | 5.19 ± 0.72 b |
| Total phenylpropanoid | 2373.74 ± 45.06 b | 3342.88 ± 241.55 a | 2137.77 ± 27.87 c | 1506.23 ± 25.61 d |
| Time (h) | Condition | Gallic Acid | 4-Hydroxybenzoic Acid | Caffeic Acid | Epicatechin | Ferulic Acid |
|---|---|---|---|---|---|---|
| 6 | Control | 8.11 ± 0.09 h | 33.99 ± 1.7 g | 10.61 ± 0.86 f | 6716.99 ± 123.18 d | 37.99 ± 3.25 f |
| NaCl 50mM | 8.91 ± 0.26 gh | 46.31 ± 1.15 f | 19.91 ± 0.68 cd | 7228.10 ± 89.74 c | 72.63 ± 2.61 d | |
| 12 | Control | 8.7 ± 0.34 gh | 58.51 ± 2.44 e | 20.8 ± 1.00 c | 6875.85 ± 78.56 d | 68.83 ± 4.26 d |
| NaCl 50mM | 10.53 ± 0.32 de | 59.82 ± 1.48 e | 22.74 ± 0.60 b | 8480.07 ± 140.49 b | 110.72 ± 8.16 d | |
| 24 | Control | 10.27 ± 0.083 ef | 60.55 ± 6.08 e | 18.59 ± 1.52 d | 5925.99 ± 169.33 e | 48.76 ± 3.89 e |
| NaCl 50mM | 9.41 ± 0.34 fg | 66.08 ± 1.99 d | 13.25 ± 0.34 e | 7180.61 ± 164.84 c | 75.77 ± 7.19 d | |
| 48 | Control | 11.29 ± 0.64 cd | 98.49 ± 4.43 b | 14.51 ± 1.39 e | 7224.31 ± 41.13 c | 49.77 ± 1.63 e |
| NaCl 50mM | 14.43 ± 0.39 b | 105.27 ± 1.55 a | 24.91 ± 0.32 a | 9158.68 ± 56.56 a | 100.58 ± 2.38 c | |
| 72 | Control | 16.2 ± 1.37 a | 90.39 ± 0.48 c | 22.35 ± 0.27 b | 1756.89 ± 97.70 g | 179.24 ± 8.93 a |
| NaCl 50mM | 11.94 ± 0.25 c | 93.14 ± 1.84 c | 26.08 ± 0.64 a | 4184.02 ± 113.26 f | 52.77 ± 3.46 e | |
| Time (h) | Condition | Benzoic Acid | Rutin | Trans-Cinnamic Acid | Kaempferol | Total Phenylpropanoids |
| 6 | Control | 847.63 ± 11.04 f | 129.15 ± 6.52 f | 1.68 ± 0.13 f | 5.75 ± 0.94 f | 7791.91 ± 118.28 f |
| NaCl 50mM | 878.01 ± 2.24 f | 142.55 ± 2.28 f | 1.85 ± 0.46 ef | 5.09 ± 0.11 f | 8403.41 ± 85.22 e | |
| 12 | Control | 1032.99 ± 27.20 de | 157.21 ± 6.29 e | 2.21 ± 0.02 de | 6.09 ± 0.94 ef | 8231.19 ± 81.91 e |
| NaCl 50mM | 998.95 ± 28.71 e | 156.61 ± 4.67 e | 2.32 ± 0.07 d | 6.54 ± 0.83 f | 9848.29 ± 164.88 b | |
| 24 | Control | 1016.11 ± 25.62 de | 232.47 ± 7.32 d | 4.97 ± 0.025 c | 5.85 ± 0.59 f | 7323.56 ± 208.51 g |
| NaCl 50mM | 1044.75 ± 24.02 d | 224.57 ± 14.10 d | 5.05 ± 0.23 c | 7.99 ± 0.34 c | 8627.49 ± 156.74 d | |
| 48 | Control | 1449.77 ± 33.71 a | 366.73 ± 5.79 b | 6.91 ± 0.19 b | 7.38 ± 0.98 de | 9229.18 ± 70.52 c |
| NaCl 50mM | 1261.05 ± 14.24 b | 351.68 ± 3.49 c | 6.87 ± 0.24 b | 8.7 ± 0.46 c | 11032.17 ± 42.17 a | |
| 72 | Control | 305.84 ± 6.68 g | 416.41 ± 11.08 a | 6.95 ± 0.25 b | 16.25 ± 1.29 a | 2810.52 ± 73.55 i |
| NaCl 50mM | 1148.33 ± 19.58 c | 425.98 ± 8.42 a | 9.33 ± 0.27 a | 10.53 ± 0.07 b | 5962.11 ± 142.09 h |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuong, D.M.; Kwon, S.-J.; Nguyen, B.V.; Chun, S.W.; Kim, J.K.; Park, S.U. Effect of Salinity Stress on Phenylpropanoid Genes Expression and Related Gene Expression in Wheat Sprout. Agronomy 2020, 10, 390. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030390
Cuong DM, Kwon S-J, Nguyen BV, Chun SW, Kim JK, Park SU. Effect of Salinity Stress on Phenylpropanoid Genes Expression and Related Gene Expression in Wheat Sprout. Agronomy. 2020; 10(3):390. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030390
Chicago/Turabian StyleCuong, Do Manh, Soon-Jae Kwon, Bao Van Nguyen, Se Won Chun, Jae Kwang Kim, and Sang Un Park. 2020. "Effect of Salinity Stress on Phenylpropanoid Genes Expression and Related Gene Expression in Wheat Sprout" Agronomy 10, no. 3: 390. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030390