Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material and Design
2.2. Measurements
2.2.1. Growth and Yield Traits
2.2.2. Biochemical Measurements
2.2.3. Enzyme Activity
2.2.4. Macronutrient and Pb Analysis
2.3. Statistical Analysis
3. Results
3.1. Growth and Yield Traits
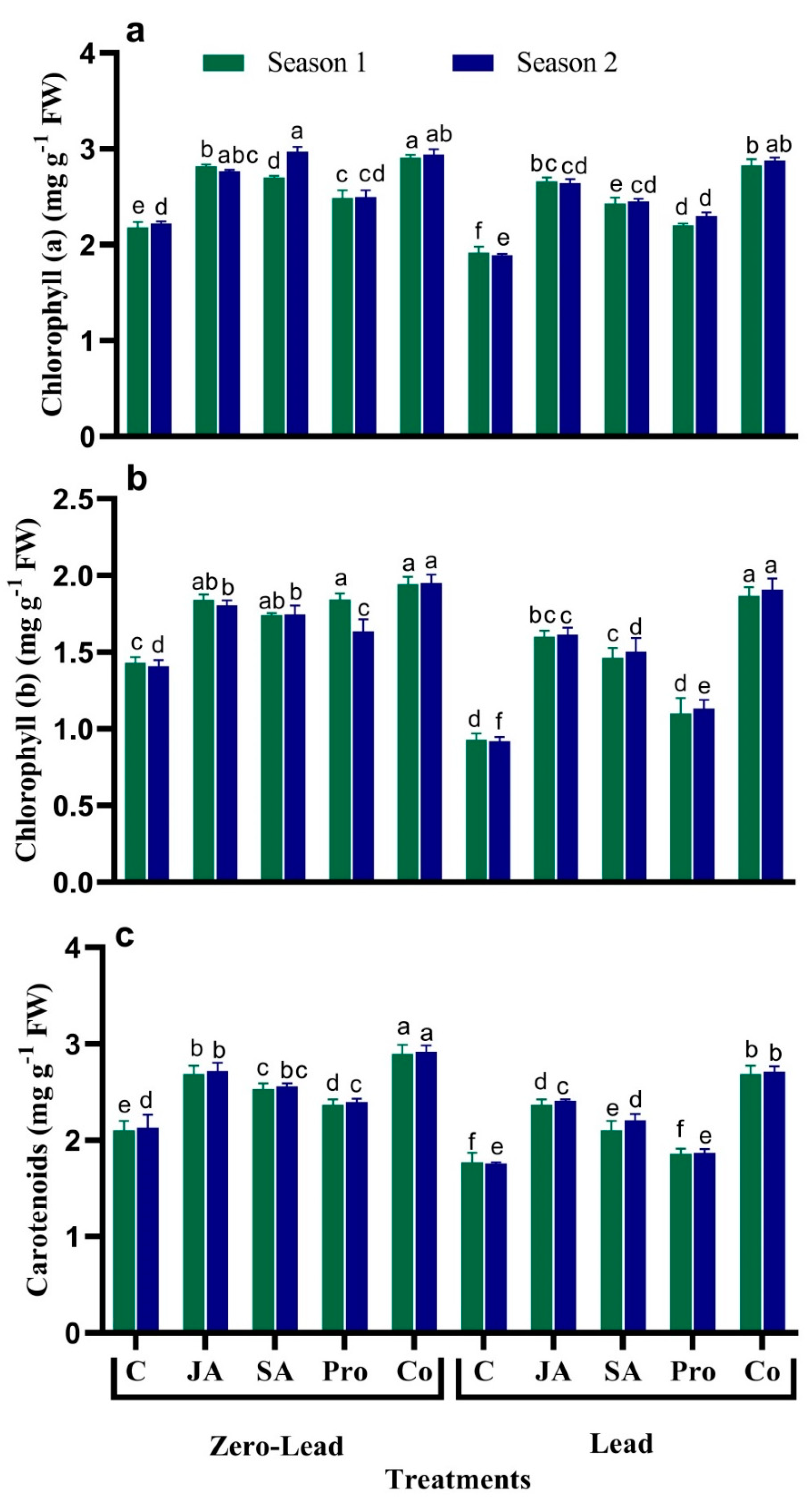
3.2. Chlorophyll Content
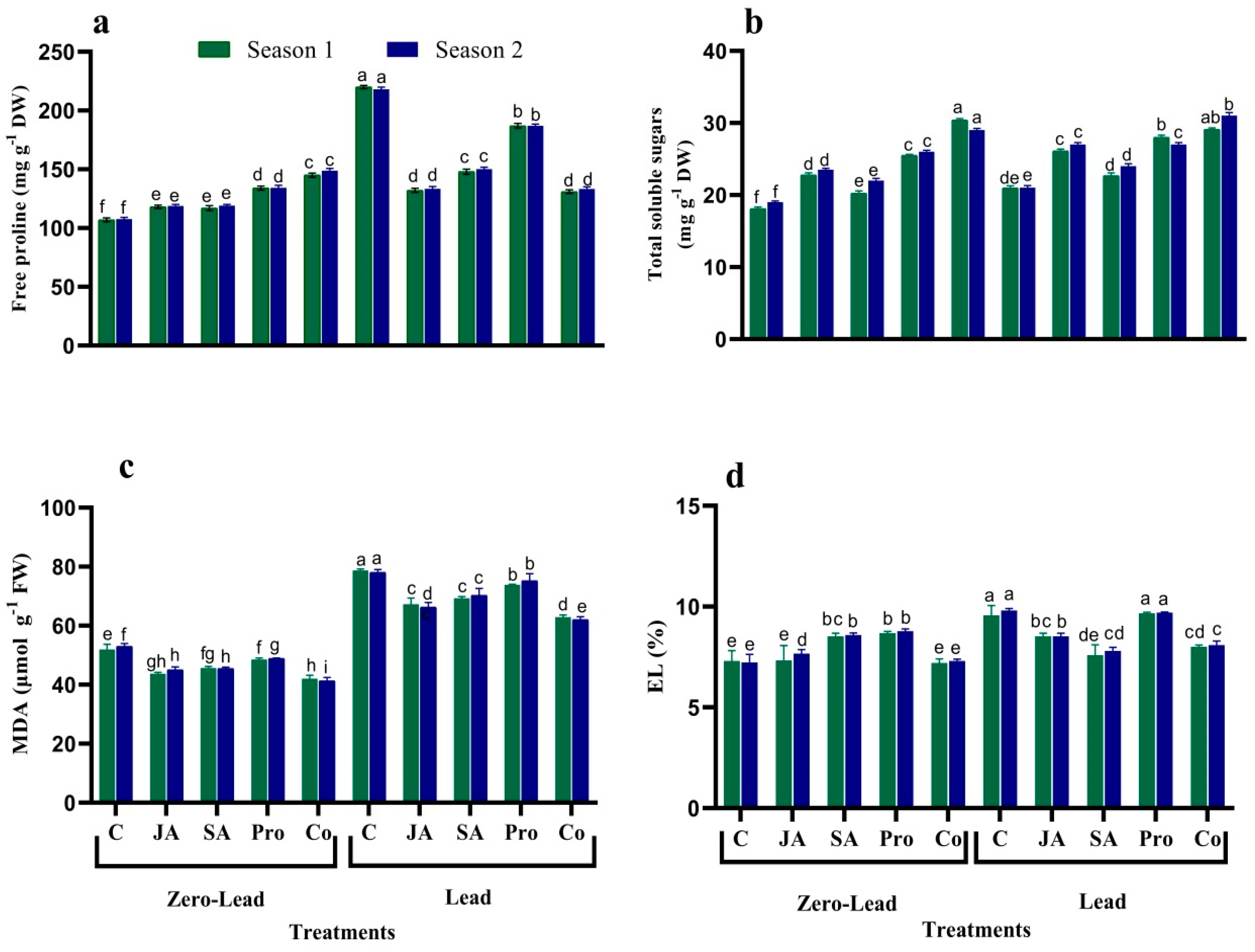
3.3. Osmolyte Content
3.4. Lipid Peroxidation and EL
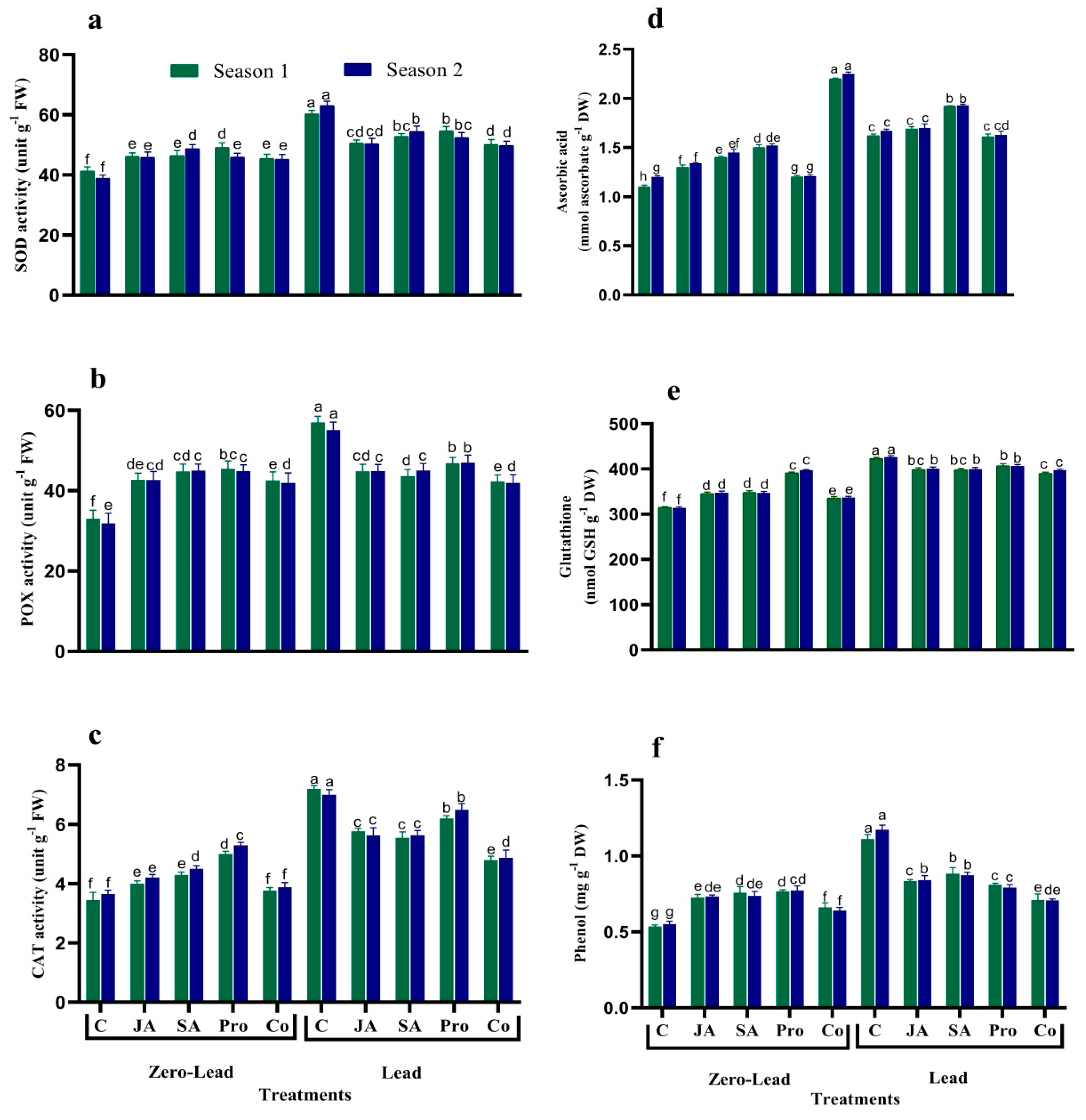
3.5. Antioxidant Enzymes
3.6. GSH, AsA, and Phenols
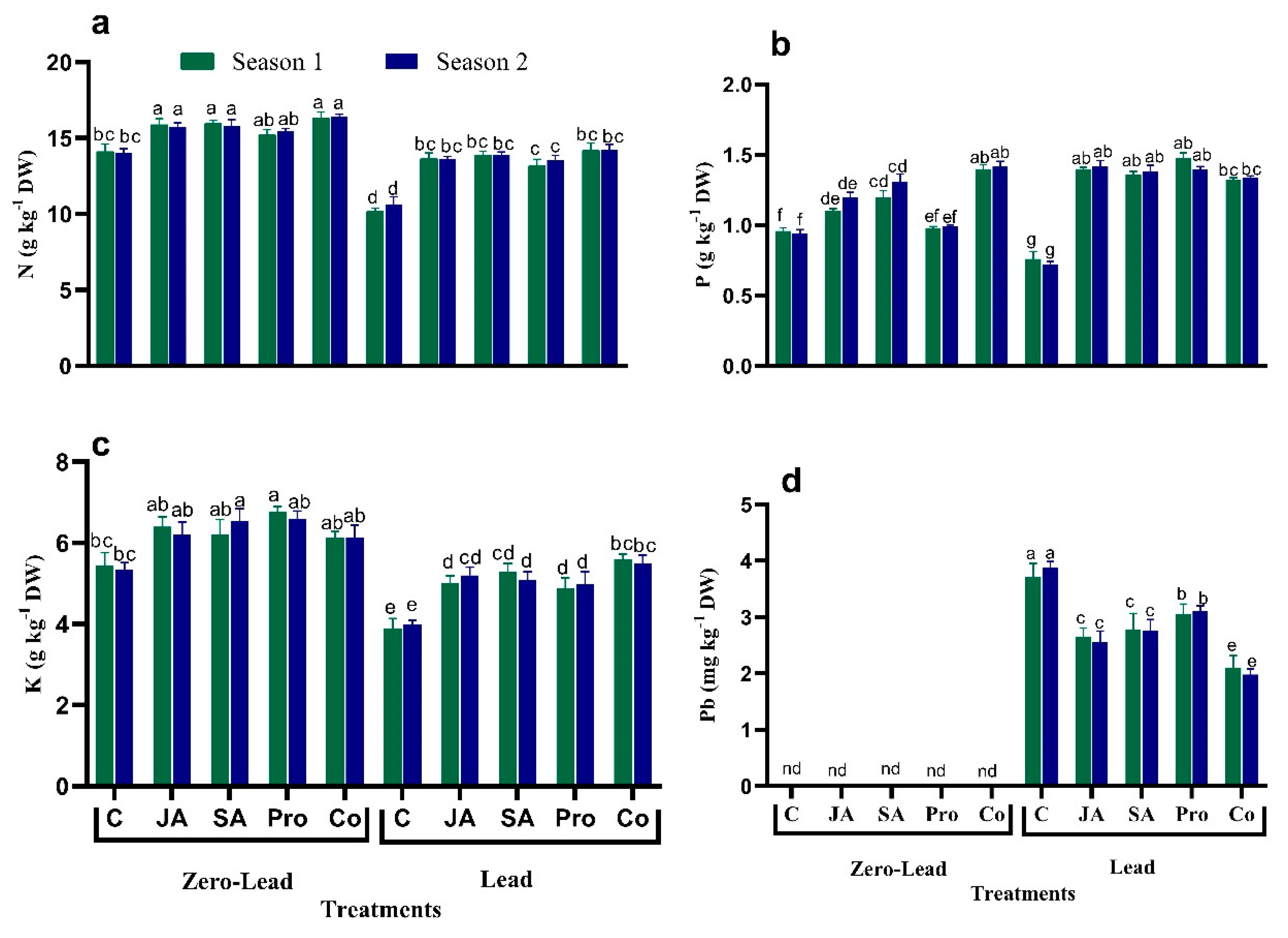
3.7. Macronutrients Concentration and Pb
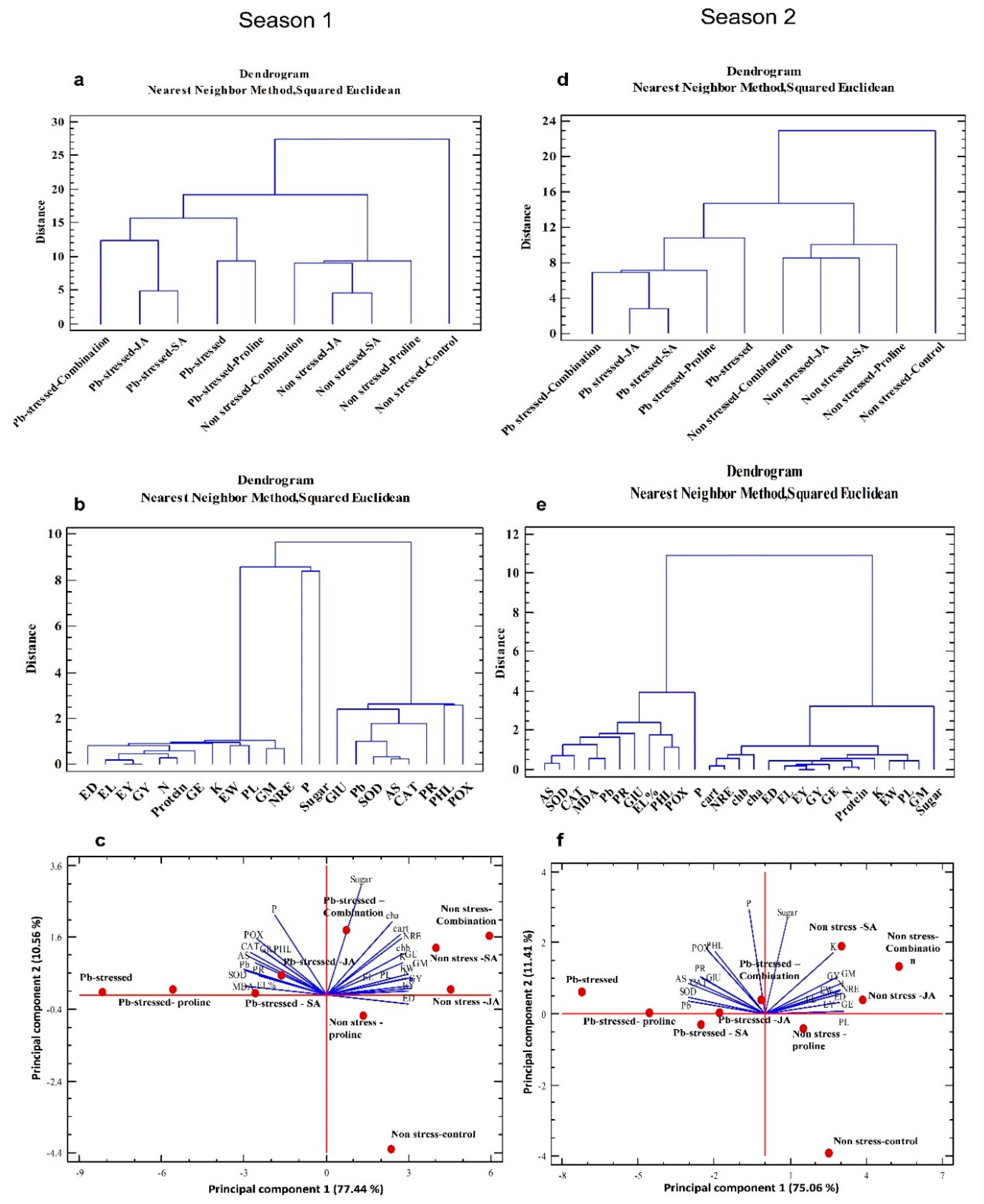
3.8. Multivariate Statistical Approach
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ashraf, M.Y.; Roohi, M.; Iqbal, Z.; Ashraf, M.; Öztürk, M.; Gücel, S. Cadmium (Cd) and lead (Pb) induced changes in growth, some biochemical attributes, and mineral accumulation in two cultivars of mung bean [Vigna radiata (L.) Wilczek]. Commun. Soil Sci. Plant Anal. 2016, 47, 405–413. [Google Scholar]
- Saleem, M.H.; Ali, S.; Seleiman, M.F.; Rizwan, M.; Rehman, M.; Aisha Akram, N.; Liu, L.; Alotaibi, M.; Al-Ashkar, I.; Mubushar, M. Assessing the Correlations between Different Traits in Copper-Sensitive and Copper-Resistant Varieties of Jute (Corchorus capsularis L.). Plants (Basel) 2019, 8, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seleiman, M.F.; Ali, S.; Refay, Y.; Rizwan, M.; Alhammad, B.A.; El-Hendawy, S.E. Chromium resistant microbes and melatonin reduced Cr uptake and toxicity, improved physio-biochemical traits and yield of wheat in contaminated soil. Chemosphere 2020, 126239. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Santanen, A.; Mäkelä, P.S. Recycling sludge on cropland as fertilizer–Advantages and risks. Resour. Conserv. Recycl. 2020, 155, 104647. [Google Scholar] [CrossRef]
- Mohamed, H.I. Molecular and biochemical studies on the effect of gamma rays on lead toxicity in cowpea (Vigna sinensis) plants. Biol. Trace Elem. Res. 2011, 144, 1205–1218. [Google Scholar] [CrossRef]
- Malar, S.; Vikram, S.S.; Favas, P.J.; Perumal, V. Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)]. Bot. Stud. 2014, 55, 54. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Sakran, A.M.; Basalah, M.O.; Ali, H.M. Effect of calcium and potassium on antioxidant system of Vicia faba L. under cadmium stress. J. Int. J. Mol. Sci. 2012, 13, 6604–6619. [Google Scholar] [CrossRef] [Green Version]
- Sharaf, A.; Farghal, I.I.; Sofy, M.R. Role of gibberellic acid in abolishing the detrimental effects of Cd and Pb on broad bean and lupin plants. Res. J. Agric. Biol. Sci. 2009, 5, 668–673. [Google Scholar]
- El-Beltagi, H.S.; Mohamed, H.I. Alleviation of cadmium toxicity in Pisum sativum L. seedlings by calcium chloride. Not. Bot. Horti Agrobot. Cluj. Napoca 2013, 41, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P.S. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in Boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Akladious, S.A.; Mohamed, H.I. Physiological role of exogenous nitric oxide in improving performance, yield and some biochemical aspects of sunflower plant under zinc stress. Acta Biol. Hung. 2017, 68, 101–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, H.M.; Abdel-Salam, A.; Abdel-Salam, M.A.; Seleiman, M.F. Phytoremediation of metal and metalloids from contaminated soil. In Plants Under Metal and Metalloid Stress; Springer Nature Singapore Pte Ltd.: Singapore, 2018; pp. 249–262. [Google Scholar]
- Mohamed, H.I.; Latif, H.H.; Hanafy, R.S. Influence of nitric oxide application on some biochemical aspects, endogenous hormones, minerals and phenolic compounds of Vicia faba plant grown under arsenic stress. Gesunde Pflanzen 2016, 68, 99–107. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Bano, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.A.; Khan, F.; Chen, Y.; Wu, C. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: Consequences for changing environment. Environ. Sci. Pollut. Res. Int. 2015, 22, 4907–4921. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant. Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Aslam, M.; Saeed, M.S.; Sattar, S.; Sajad, S.; Sajjad, M.; Adnan, M.; Iqbal, M.; Sharif, M.T. Specific role of proline against heavy metals toxicity in plants. Int. J. Pure Appl. Biosci. 2017, 5, 27–34. [Google Scholar] [CrossRef]
- Wasternack, C. Action of jasmonates in plant stress responses and development—Applied aspects. Biotechnol. Adv. 2014, 32, 31–39. [Google Scholar] [CrossRef]
- Maksymiec, W.; Krupa, Z. Jasmonic acid and heavy metals in Arabidopsis plants-a similar physiological response to both stressors? J. Plant Physiol. 2002, 159, 509–515. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Latif, H.H. Improvement of drought tolerance of soybean plants by using methyl jasmonate. Physiol. Mol. Bio Plants 2017, 23, 545–556. [Google Scholar] [CrossRef]
- Ashry, N.A.; Ghonaim, M.M.; Mohamed, H.I.; Mogazy, A.M. Physiological and molecular genetic studies on two elicitors for improving the tolerance of six Egyptian soybean cultivars to cotton leaf worm. Plant. Physiol. Biochem. 2018, 130, 224–234. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. USA 1995, 92, 4114–4119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poonam, S.; Kaur, H.; Geetika, S. Effect of jasmonic acid on photosynthetic pigments and stress markers in Cajanus cajan (L.) Millsp. seedlings under copper stress. Am. J. Plant. Sci. 2013, 4, 817. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, I.; Ahmadi, N.; Ghanati, F.; Sadeghi, M. Effects of salicylic acid applied pre-or post-transport on post-harvest characteristics and antioxidant enzyme activity of gladiolus cut flower spikes. N. Z. J. Crop. Hortic. Sci. 2015, 43, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ding, Y.; Wang, F.; Ye, Y.; Zhu, C. Role of salicylic acid in resistance to cadmium stress in plants. Plant. Cell Rep. 2016, 35, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.H.; Seleiman, M.F. Response of barley quality traits, yield and antioxidant enzymes to water-stress and chemical inducers. Intern. J. Plant Prod. 2017, 11, 477–490. [Google Scholar]
- Hay, R.; Gilbert, R. Variation in the harvest index of tropical maize: Evaluation of recent evidence from Mexico and Malawi. Ann. Appl. Biol. 2001, 138, 103–109. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Kheir, A.M. Saline soil properties, quality and productivity of wheat grown with bagasse ash and thiourea in different climatic zones. Chemosphere 2018, 193, 538–546. [Google Scholar] [CrossRef]
- Pimentel, D.; Patzek, T.W. Ethanol production using corn, switchgrass, and wood; biodiesel production using soybean and sunflower. Nat. Resour. Res. 2005, 14, 65–76. [Google Scholar] [CrossRef]
- Bekrić, V.; Radosavljević, M. Savremeni pristupi upotrebe kukuruza. Časopis za Procesnu Tehniku i Energetiku u Poljoprivredi 2008, 12, 93–96. [Google Scholar]
- Seleiman, M.F.; Santanen, A.; Stoddard, F.L.; Mäkelä, P. Feedstock quality and growth of bioenergy crops fertilized with sewage sludge. Chemosphere 2012, 89, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Santanen, A.; Jaakkola, S.; Ekholm, P.; Hartikainen, H.; Stoddard, F.L.; Mäkelä, P.S. Biomass yield and quality of bioenergy crops grown with synthetic and organic fertilizers. Biomass Bioenergy 2013, 59, 477–485. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Kleemola, J.; Stoddard, F.L.; Mäkelä, P.S. Improved sustainability of feedstock production with sludge and interacting mycorrhiza. Chemosphere 2013, 91, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization of the United Nations Statistics Division. Available online: http://faostat.fao.org/site/567/DesktopDefault.aspx (accessed on 23 February 2020).
- Ouda, S.; Mohamed, S.G.; Khalil, F.A. Modeling the effect of different stress conditions on maize productivity using yield-stress model. Int. J. Nat. Eng. Sci. 2008, 2, 57–62. [Google Scholar]
- Arshad, M.; Lowery, B.; Grossman, B. Physical tests for monitoring soil quality. In Methods for Assessing Soil Quality; Doran, J.W., Jones, A.J., Eds.; Special Publication 49; Soil Science Society of America: Madison, WI, USA, 1997; pp. 123–141. [Google Scholar]
- Thomas, G.W. Soil pH and soil acidity. In Methods of Soil Analysis: Part 3; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1996; pp. 475–490. [Google Scholar]
- Gupta, P.K. Soil, Plant, Water and Fertilizer Analysis; Agrobios: Jodhpur, India, 2009. [Google Scholar]
- Vernon, L.; Seely, G. The Chlorophylls; Academic Press: New York, NY, USA, 1966. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 350–382. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Sullivan, C.Y. Selection for drought and heat tolerance in grain sorghum. In Stress Physiology in Crop Plants; Mussell, H., Staples, R.C., Eds.; John Wiley & Sons: New York, NY, USA, 1979; pp. 263–281. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.; Einerich, D.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Griffith, O.W.J. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Mukherjee, S.; Choudhuri, M. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Galicia, L.N.; Rosales, E.; Rojas, A.P.N. Laboratory Protocols: Maize Nutrition Quality and Plant Tissue Analysis Laboratory; CIMMYT: Mexico, DF, Mexico, 2009. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Bergmeyer, H.U.; Gawehn, K. Methods of Enzymatic Analysis; Elsevier Academic Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Chen, Y.; Cao, X.; Lu, Y.; Wang, X. Effects of rare earth metal ions and their EDTA complexes on antioxidant enzymes of fish liver. Bull. Environ. Contam. Toxicol. 2000, 65, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, W. Official Methods of Analysis of the Association of Official Agricultural Chemists, 8th ed.; Association of Official Agricultural Chemists: Washington, DC, USA, 1956; p. 347. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall of India Pvt. Ltd.: New Delhi, India, 1967; pp. 144–197, 326–338. [Google Scholar]
- Page, A.I.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Barceló, J.; Poschenrieder, C. Plant water relations as affected by heavy metal stress: A review. J. Plant Nutr. 1990, 13, 1–37. [Google Scholar] [CrossRef]
- Piotrowska, A.; Bajguz, A.; Godlewska-Żyłkiewicz, B.; Czerpak, R.; Kamińska, M. Jasmonic acid as modulator of lead toxicity in aquatic plant Wolffia arrhiza (Lemnaceae). Environ. Exp. Bot. 2009, 66, 507–513. [Google Scholar] [CrossRef]
- Shakirova, F.; Bezrukova, M.; Yuldashev, R.; Fatkhutdinova, R.; Murzabaev, A. Involvement of lectin in the salicylic acid-induced wheat tolerance to cadmium and the role of endogenous ABA in the regulation of its level. Dokl Biol. Sci. 2013, 488, 49–51. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Ahmad, A. Proline enhances antioxidative enzyme activity, photosynthesis and yield of Cicer arietinum L. exposed to cadmium stress. Acta Bot. Croat 2013, 72, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Alyemeni, M.N.; Hayat, Q.; Wijaya, L.; Hayat, S. Effect of salicylic acid on the growth, photosynthetic efficiency and enzyme activities of leguminous plant under cadmium stress. Not. Bot. Horti Agrobot. Cluj. Napoca 2014, 42, 440–445. [Google Scholar] [CrossRef] [Green Version]
- Kwok, D.; Shetty, K. Effects of proline and proline analogs on total phenolic and rosmarinic acid levels in shoot clones of thyme (Thymus vulgaris L.). J. Food Biochem. 1998, 22, 37–51. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q. Role of Proline and Salicylic Acid in Overcoming the Stress of Cadmium: Chickpea (Cicer Arietinum); LAP Lambert Academic Publishing: Saarbrücken, Germany, 2011. [Google Scholar]
- Ahmad, M.S.A.; Hussain, M.; Ijaz, S.; Alvi, A.K. Photosynthetic performance of two mung bean (Vigna radiata) cultivars under lead and copper stress. Int. J. Agric. Biol. 2008, 10, 167–172. [Google Scholar]
- Cenkci, S.; Ciğerci, İ.H.; Yıldız, M.; Özay, C.; Bozdağ, A.; Terzi, H.J.E. Lead contamination reduces chlorophyll biosynthesis and genomic template stability in Brassica rapa L. Environ. Exp. Bot. 2010, 67, 467–473. [Google Scholar] [CrossRef]
- Bali, S.; Kaur, P.; Kohli, S.K.; Ohri, P.; Thukral, A.K.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Jasmonic acid induced changes in physio-biochemical attributes and ascorbate-glutathione pathway in Lycopersicon esculentum under lead stress at different growth stages. Sci. Total Environ. 2018, 645, 1344–1360. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.A.D.; Siddiqui, M.H.; Al-Khaishany, M.Y.; Ali, H.M.; Al-Amri, A.; AlRabiah, H.K. Exogenous application of salicylic acid improves tolerance of wheat plants to lead stress. Adv. Agric. Sci. 2018, 6, 25–35. [Google Scholar]
- Zouari, M.; Ahmed, C.B.; Zorrig, W.; Elloumi, N.; Rabhi, M.; Delmail, D.; Rouina, B.B.; Labrousse, P.; Abdallah, F.B. Exogenous proline mediates alleviation of cadmium stress by promoting photosynthetic activity, water status and antioxidative enzymes activities of young date palm (Phoenix dactylifera L.). Ecotoxicol. Environ. Saf. 2016, 128, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Deshna, D.; Bafna, A. Effect of lead stress on chlorophyll content, malondialdehyde and peroxidase activity in seedlings of mung bean (Vigna radiata). Int. J. Res. Chem. Environ. 2013, 3, 20–25. [Google Scholar]
- Ali, E.; Hussain, N.; Shamsi, I.H.; Jabeen, Z.; Siddiqui, M.H.; Jiang, L.-X. Role of jasmonic acid in improving tolerance of rapeseed (Brassica napus L.) to Cd toxicity. J. Zhejiang Univ. Sci. B 2018, 19, 130–146. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.; Gaur, J. Heavy-metal-induced proline accumulation and its role in ameliorating metal toxicity in Chlorella vulgaris. New Phytol. 1999, 143, 253–259. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Boguszewska, D.; Zagdańska, B. ROS as signaling molecules and enzymes of plant response to unfavorable environmental conditions. In Oxidative Stress–Molecular Mechanisms and Biological Effects; InTech: Rijeka, Croatia, 2012; pp. 341–362. [Google Scholar]
- Hattab, S.; Hattab, S.; Flores-Casseres, M.L.; Boussetta, H.; Doumas, P.; Hernandez, L.E.; Banni, M. Characterisation of lead-induced stress molecular biomarkers in Medicago sativa plants. Environ. Exp. Bot. 2016, 123, 1–12. [Google Scholar] [CrossRef]
- Akladious, S.A.; Mohamed, H.I. Ameliorative effects of calcium nitrate and humic acid on the growth, yield component and biochemical attribute of pepper (Capsicum annuum) plants grown under salt stress. Sci. Hortic. 2018, 236, 244–250. [Google Scholar] [CrossRef]
- Fontenele, N.M.B.; Otoch, M.; Gomes-Rochette, N.F.; Menezes Sobreira, A.C.; Barreto, A.A.G.C.; Oliveira, F.D.B.; Costa, J.H.; Borges, S.; Nascimento, R.F.; Melo, D.F. Effect of lead on physiological and antioxidant responses in two Vigna unguiculata cultivars differing in Pb-accumulation. Chemosphere 2017, 176, 397–404. [Google Scholar] [CrossRef]
- Maksymiec, W.; Wojcik, M.; Krupa, Z. Variation in oxidative stress and photochemical activity in Arabidopsis thaliana leaves subjected to cadmium and excess copper in the presence or absence of jasmonate and ascorbate. Chemosphere 2007, 66, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Kandziora-Ciupa, M.; Ciepał, R.; Nadgórska-Socha, A.; Barczyk, G. Accumulation of heavy metals and antioxidant responses in Pinus sylvestris L. needles in polluted and non-polluted sites. Ecotoxicology 2016, 25, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sinha, S. Accumulation of metals and its effects in Brassica juncea (L.) Czern.(cv. Rohini) grown on various amendments of tannery waste. Ecotoxicol. Environ. Saf. 2005, 62, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Matin, M.; Fardus, J.; Hasanuzzaman, M.; Hossain, M.; Parvin, K. Foliar application of salicylic acid improves growth and yield attributes by upregulating the antioxidant defense system in Brassica campestris plants grown in lead-amended soils. Acta Agrobot 2019, 72. [Google Scholar] [CrossRef]
- Doncheva, S.; Moustakas, M.; Ananieva, K.; Chavdarova, M.; Gesheva, E.; Vassilevska, R.; Mateev, P. Plant response to lead in the presence or absence EDTA in two sunflower genotypes (cultivated H. annuus cv. 1114 and interspecific line H. annuus × H. argophyllus). Environ. Sci. Pollut. Res. Int. 2013, 20, 823–833. [Google Scholar] [CrossRef]
- Bali, S.; Jamwal, V.L.; Kaur, P.; Kohli, S.K.; Ohri, P.; Gandhi, S.G.; Bhardwaj, R.; Al-Huqail, A.A.; Siddiqui, M.H.; Ahmad, P. Role of P-type ATPase metal transporters and plant immunity induced by jasmonic acid against lead (Pb) toxicity in tomato. Ecotoxicol. Environ. Saf. 2019, 174, 283–294. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Jin, X.; Yang, X.; Islam, E.; Liu, D.; Mahmood, Q. Effects of cadmium on ultrastructure and antioxidative defense system in hyperaccumulator and non-hyperaccumulator ecotypes of Sedum alfredii Hance. J. Hazard. Mater. 2008, 156, 387–397. [Google Scholar] [CrossRef]
- Phang, C.; Leung, D.W.; Taylor, H.H.; Burritt, D. The protective effect of sodium nitroprusside (SNP) treatment on Arabidopsis thaliana seedlings exposed to toxic level of Pb is not linked to avoidance of Pb uptake. Ecotoxicol. Environ. Saf. 2011, 74, 1310–1315. [Google Scholar] [CrossRef]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Dietz, K.-J. Salicylic acid alleviates the cadmium toxicity in barley seedlings. J. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q.; Ashraf, M.; Shahbaz, M.; Humera, H. Ameliorating effect of foliar applied proline on nutrient uptake in water stressed maize (Zea mays L.) plants. Pak. J. Bot. 2008, 40, 211–219. [Google Scholar]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of ascorbic acid, glutathione and proline applied as singly or in sequence combination in improving chickpea plant through physiological change and antioxidant defense under different levels of irrigation intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Season 2017 | Season 2018 |
|---|---|---|
| TSS, ppm | 772 | 782 |
| pH | 7.9 | 7.6 |
| EC (dS cm−1) * | 1.75 | 1.78 |
| Cations (mmolc L−1) * | ||
| Na+ | 2.63 | 2.67 |
| K+ | 0.62 | 0.65 |
| Ca++ | 2.17 | 2.15 |
| Mg++ | 1.0 | 1.0 |
| Anion (mmolc L−1) * | ||
| Cl= | 4.3 | 4.1 |
| SO4−2 | 0.97 | 0.99 |
| HCO3− | 1.0 | 1.0 |
| CO3− | nd | nd |
| Soil texture | ||
| Sand | 26.24 | 26.36 |
| Clay | 49.19 | 48.61 |
| Silt | 24.57 | 25.03 |
| Texture | Clay loamy | Clay loamy |
| Pb Treatments | Exogenous Treatments | Plant Height (cm) | Ear Length (cm) | Ear Diameter (mm) | Number of Row Ear−1 | No. of Grains Ear−1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| S 1 | S 2 | S 1 | S 2 | S 1 | S 2 | S 1 | S 2 | S 1 | S 2 | ||
| Zero-Pb | Control | 229.0 d | 229.2 e | 18.5 d | 18.6 d | 44.0 d | 44.3 abc | 11.6 abc | 12.3 abc | 444 c | 454 c |
| JA | 244.7 b | 245.5 b | 24.2 b | 24.4 b | 46.1 b | 46.3 a | 13.6 ab | 14.3 a | 493 b | 496 ab | |
| SA | 237.6 c | 237.8 c | 22.1 c | 22.4 c | 45.6 bc | 48.8 a | 13.0 abc | 13.0 abc | 444 c | 446 c | |
| Pro | 231.5 d | 231.4 d | 18.7 d | 19.1 d | 44.9 c | 44.9 ab | 13.0 abc | 13.6 ab | 426 d | 466 bc | |
| Combination | 258.0 a | 248.4 a | 26.2 a | 26.3 a | 47.5 a | 47.6 a | 14.3 a | 14.3 a | 532 a | 534 a | |
| Pb | Control | 150.6 h | 150.8 i | 10.7 h | 10.9 h | 33.3 i | 33.4 d | 10.3 c | 9.6 d | 301 i | 301 f |
| JA | 200.6 e | 200.9 f | 15.0 f | 15.2 f | 38.3 f | 38.5 bcd | 12.3 abc | 13.0 abc | 350 f | 334 ef | |
| SA | 189.5 f | 189.9 g | 14.3 f | 14.6 f | 35.4 g | 35.6 d | 11.6 abc | 11.6 bcd | 341 g | 345 e | |
| Pro | 170.7 g | 170.7 h | 12.2 g | 12.5 g | 34.3 h | 34.5 d | 11.0 bc | 11.0 cd | 321 h | 331 ef | |
| Combination | 230.0 d | 216.9 e | 17.4 e | 17.7 e | 40.2 e | 38.1 cd | 13.6 ab | 13.0 abc | 399 e | 402 d | |
| S.E.M. | 1.87 | 1.24 | 0.20 | 0.17 | 0.17 | 1.35 | 0.56 | 0.52 | 0.88 | 7.90 | |
| Significance | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | |
| Lead (Pb) | ** | ** | ** | ** | ** | ** | * | ** | ** | ** | |
| Acids (A) | ** | ** | ** | ** | ** | * | ** | ** | ** | ** | |
| Interaction (Pb × A) | ** | ** | ** | ** | ** | ns | ns | ns | ** | * | |
| Lead (Pb) | Exogenous Treatments | Ear Weight (g) | 100-Grain Weight (g) | Ear Yield (kg ha−1) | |||
|---|---|---|---|---|---|---|---|
| S 1 | S 2 | S 1 | S 2 | S 1 | S 2 | ||
| Zero-Pb | Control | 170.0 d | 172.0 d | 30.3 c | 28.0 bc | 9851 d | 9886 d |
| JA | 189.3 ab | 190.3 b | 34.0 ab | 34.3 ab | 11,382 b | 11,660 b | |
| SA | 184.0 bc | 188.0 bc | 33.3 ab | 33.6 ab | 10,775 c | 10,923 c | |
| Pro | 178.7 c | 182.3 c | 33.0 b | 33.0 ab | 9851 d | 9966 d | |
| Combination | 193.3 a | 198.0 a | 35.3 a | 35.3 a | 12,203 a | 12,544 a | |
| Pb | Control | 127.3 g | 134.0 g | 20.7 f | 21.3 d | 6490 i | 6523 g |
| JA | 152.3 e | 152.0 e | 28.7 cd | 29.3 abc | 8316 f | 8377 e | |
| SA | 146.0 f | 145.3 f | 27.7 d | 28.3 bc | 7414 g | 7484 f | |
| Pro | 145.0 f | 137.7 g | 23.7 e | 24.3 cd | 7095 h | 7310 f | |
| Combination | 186.3 b | 184.7 bc | 29.7 cd | 30.3 abc | 8597 e | 8694 e | |
| S.E.M. | 1.25 | 1.23 | 0.43 | 1.31 | 56.73 | 80.60 | |
| Significance | ** | ** | ** | ** | ** | ** | |
| Lead (Pb) | ** | ** | ** | ** | ** | ** | |
| Acids (A) | ** | ** | ** | ** | ** | ** | |
| Interaction (Pb × A) | ** | ** | ** | ns | ** | ** | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline. Agronomy 2020, 10, 699. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050699
Sofy MR, Seleiman MF, Alhammad BA, Alharbi BM, Mohamed HI. Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline. Agronomy. 2020; 10(5):699. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050699
Chicago/Turabian StyleSofy, Mahmoud R., Mahmoud F. Seleiman, Bushra A. Alhammad, Basmah M. Alharbi, and Heba I. Mohamed. 2020. "Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline" Agronomy 10, no. 5: 699. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050699