Study of C, N, P and K Release from Residues of Newly Proposed Cover Crops in a Spanish Olive Grove

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
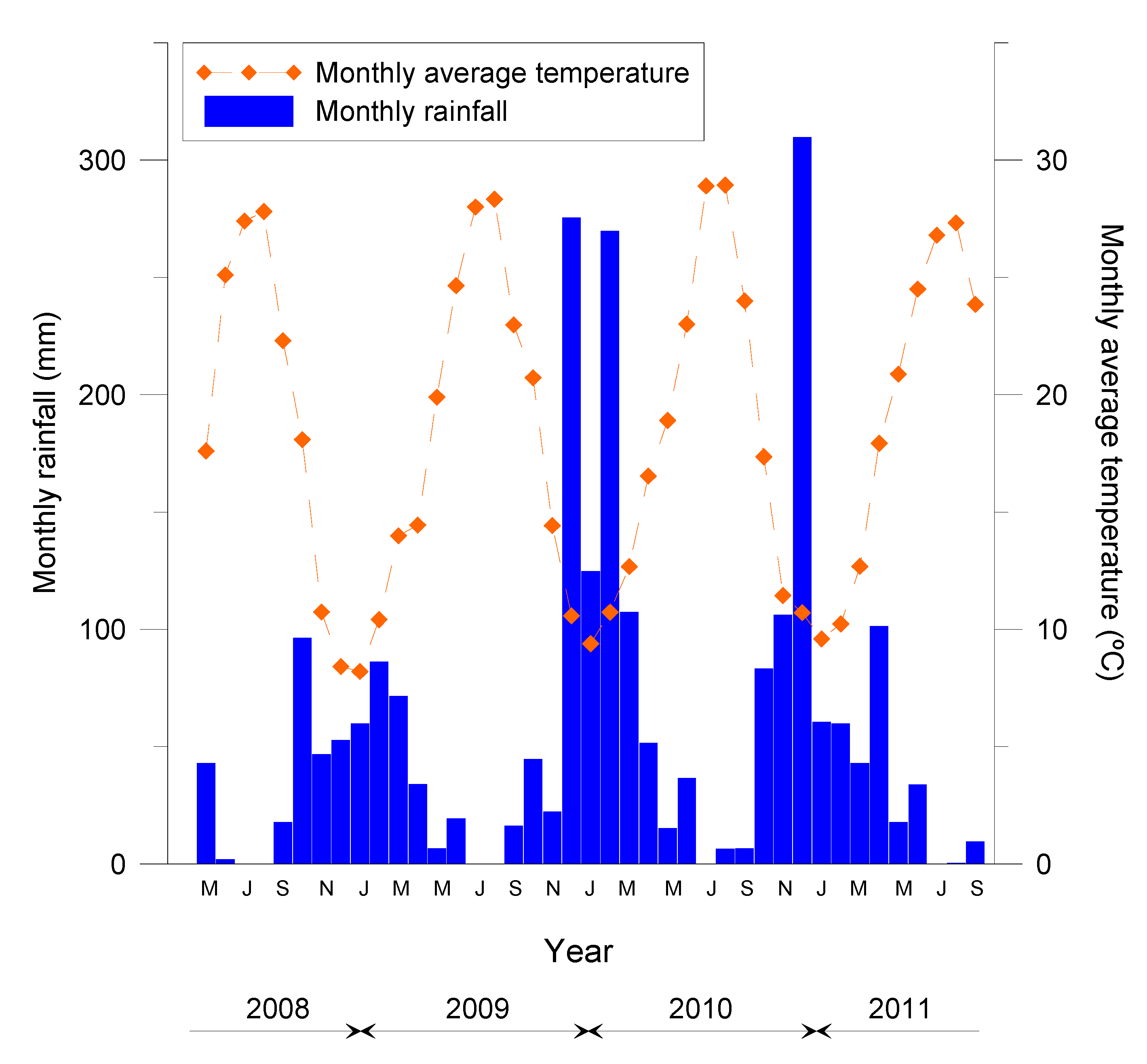
2.1. Site and Experimental Design
2.2. Sampling Scheme
2.3. Analysis of Samples
2.4. Data Analysis
2.4.1. C, N, P and K Release from Residues
2.4.2. C, N and P Modelling
2.4.3. Statistical Analysis and Software
3. Results and Discussion
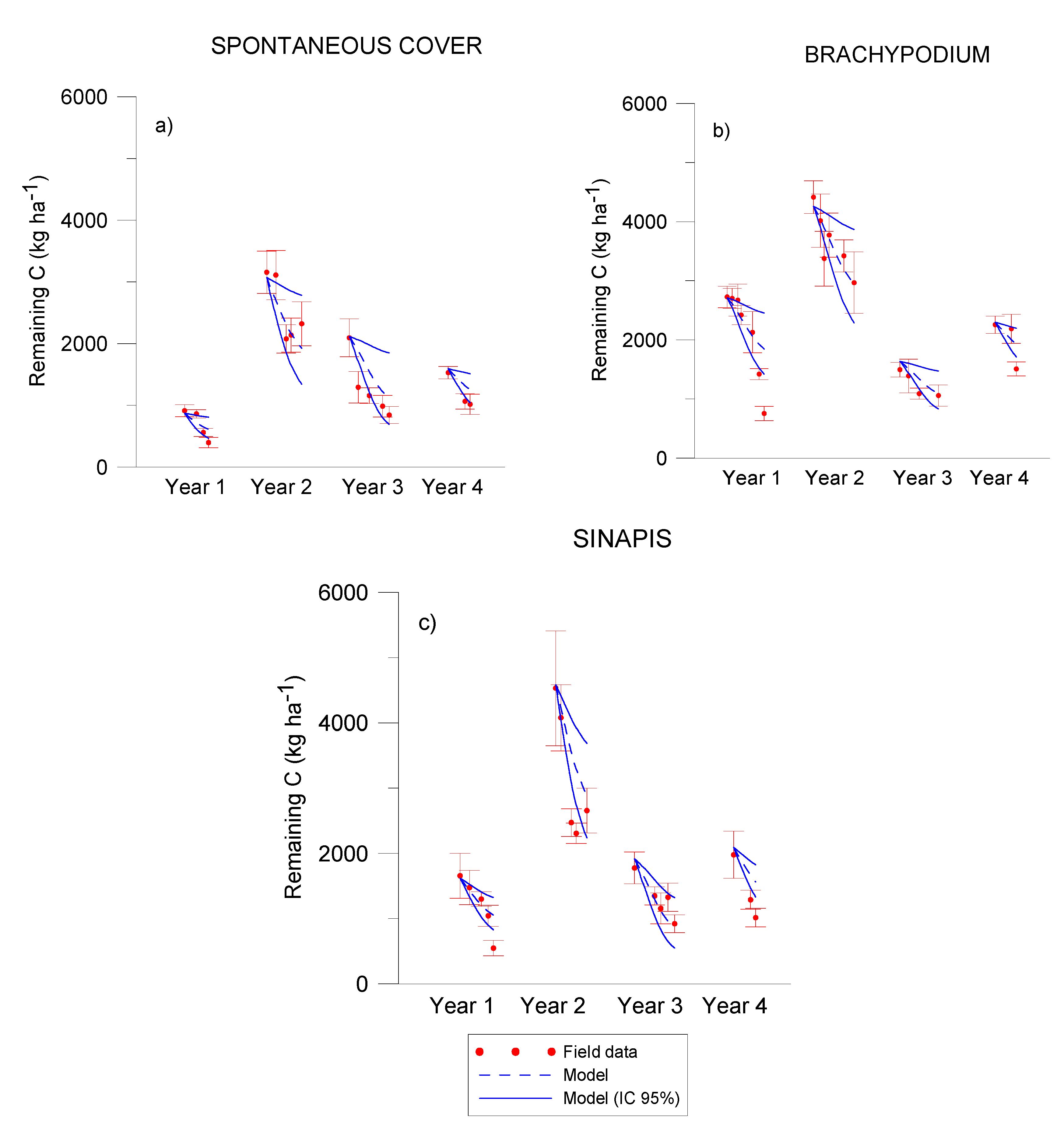
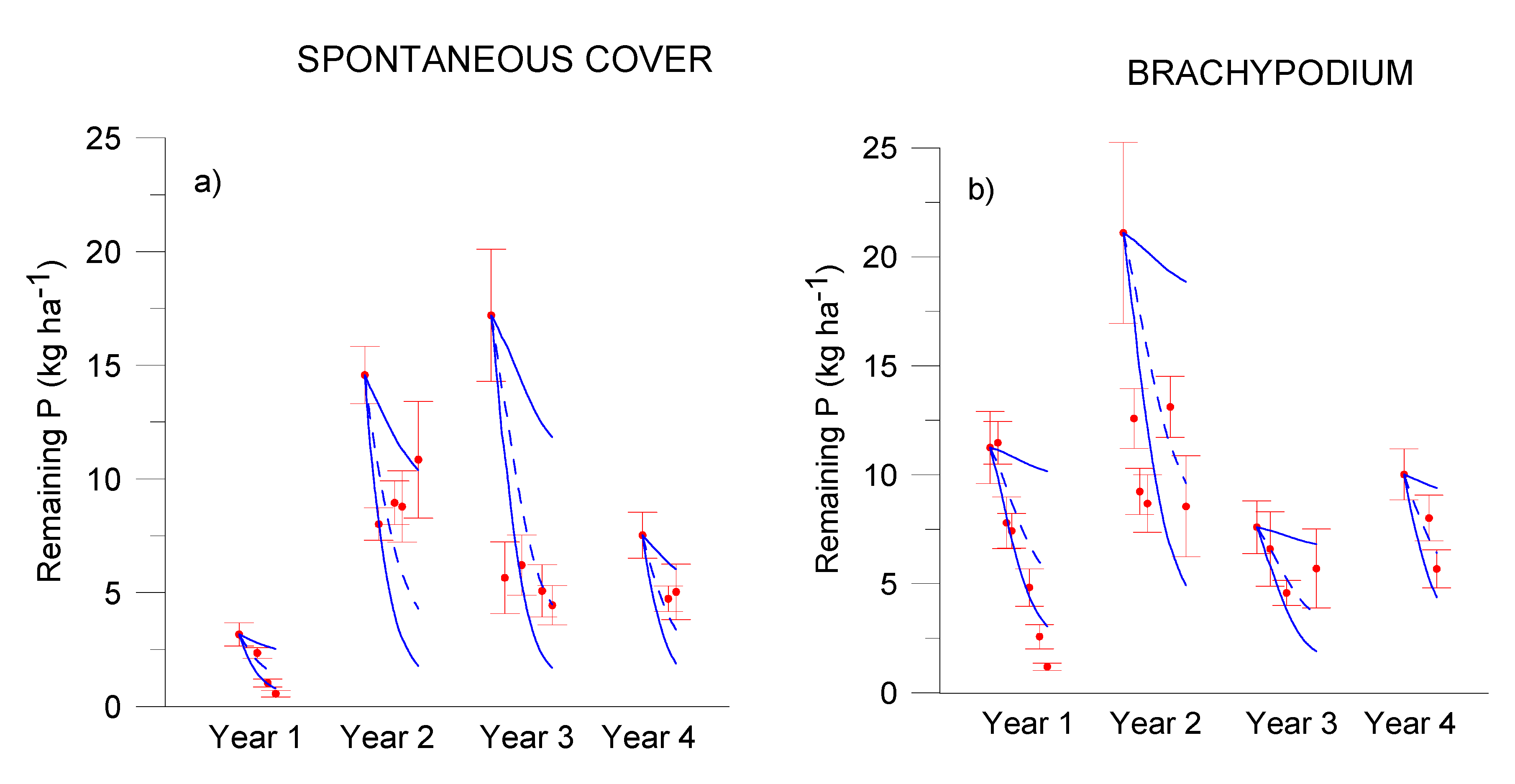
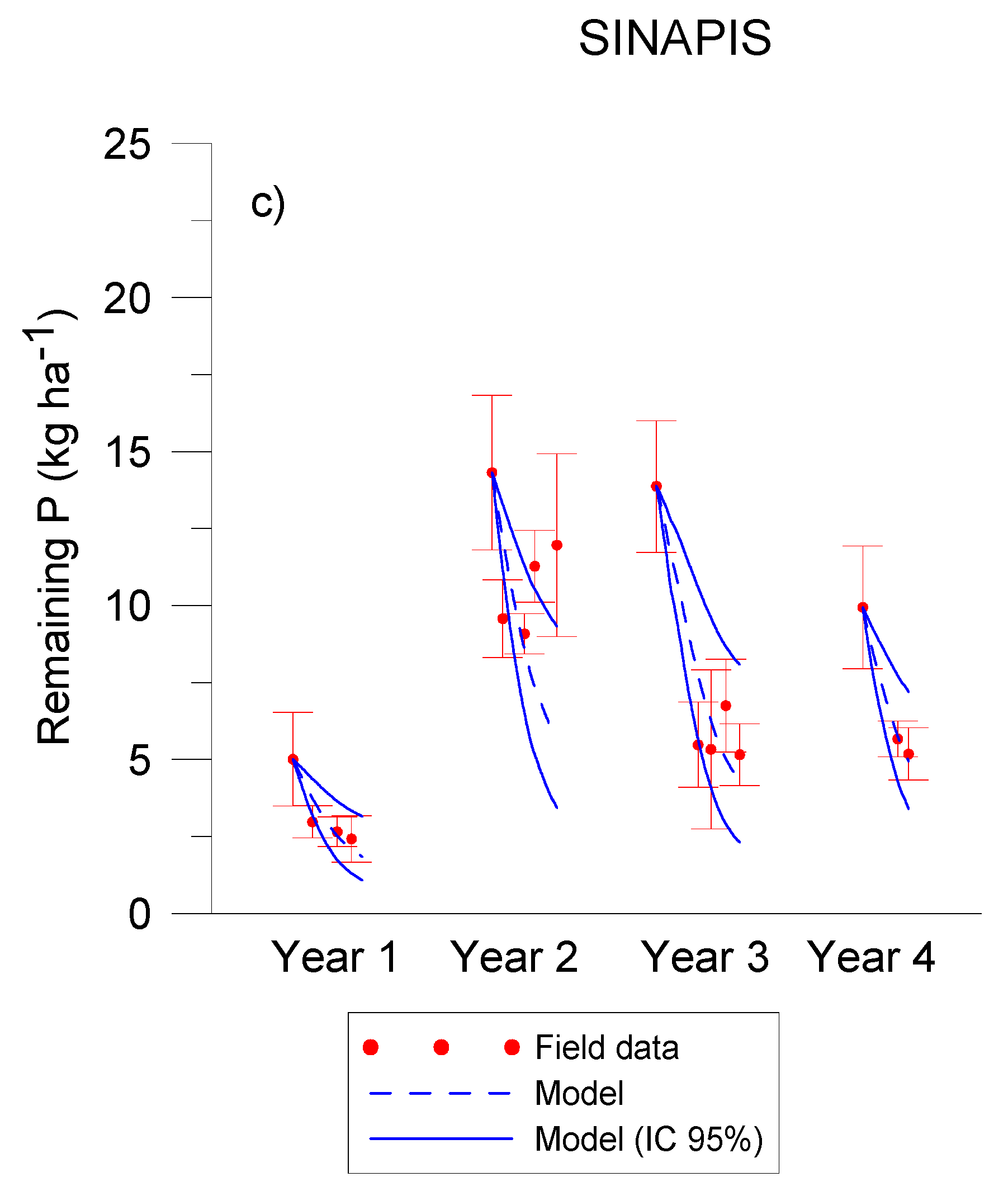
3.1. Residue Release of C, N, P, K
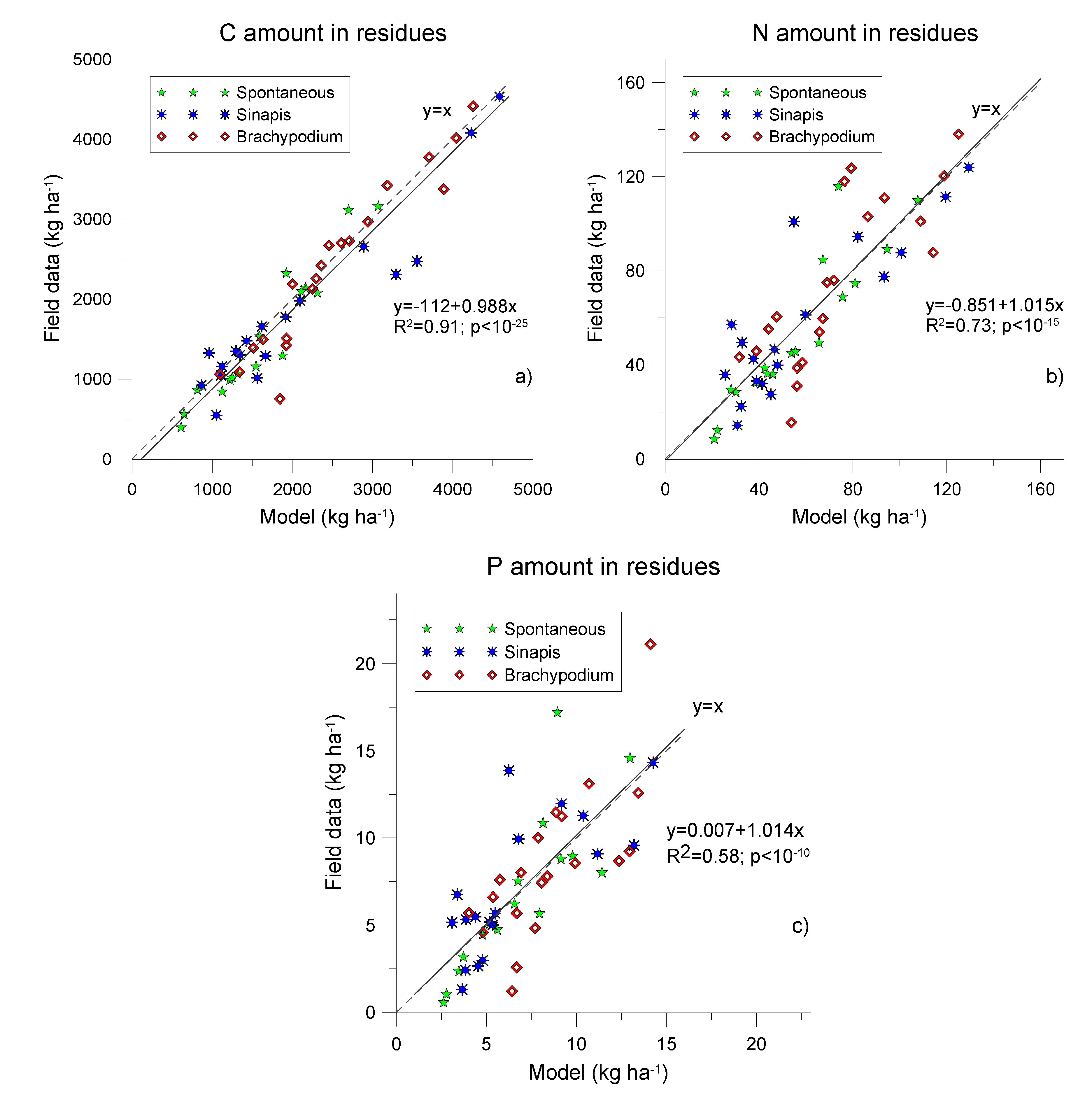
3.2. C, N and P Modelling
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO Agricultural Statistics. Available online: http://faostat.fao.org (accessed on 29 December 2019).
- Mairech, H.; López-Bernal, Á.; Moriondo, M.; Dibari, C.; Regni, L.; Proietti, P.; Villalobos, F.J.; Testi, L. Is new olive farming sustainable? A spatial comparison of productive and environmental performances between traditional and new olive orchards with the model OliveCan. Agric. Syst. 2020, 181, 102816. [Google Scholar] [CrossRef]
- MAPA. Encuesta Sobre Superficies Y Rendimientos de Cultivos (ESYRCE): Resultados 2019. 2019, p. 176. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/boletin2019_tcm30-536911.pdf (accessed on 27 May 2020).
- Cerdà, A.; Lavee, H.; Romero-Díaz, A.; Hooke, J.; Montanarella, L. Soil erosion and degradation in Mediterranean-type ecosystems. Land Degrad. Dev. 2010, 21, 71–74. [Google Scholar] [CrossRef]
- Espejo-Pérez, A.J.; Rodríguez-Lizana, A.; Ordóñez, R.; Giráldez, J.V. Soil loss and runoff reduction in olive-tree dry-farming with cover crops. Soil Sci. Soc. Am. J. 2013, 77, 2140–2148. [Google Scholar] [CrossRef] [Green Version]
- Ordóñez-Fernández, R.; Rodríguez-Lizana, A.; Espejo-Pérez, A.J.; González-Fernández, P.; Saavedra, M.M. Soil and available phosphorus losses in ecological olive groves. Eur. J. Agron. 2007, 27, 144–153. [Google Scholar] [CrossRef]
- Miranda-Fuentes, A.; Rodríguez-Lizana, A.; Gil, E.; Agüera-Vega, J.; Gil-Ribes, J.A. Influence of liquid-volume and airflow rates on spray application quality and homogeneity in super-intensive olive tree canopies. Sci. Total Environ. 2015, 537, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Parras-Alcántara, L.; Lozano-García, B.; Keesstra, S.; Cerdá, A.; Brevik, E.C. Long-term effects of soil management on ecosystem services and soil loss estimation in olive grove top soils. Sci. Total Environ. 2016, 571, 498–506. [Google Scholar] [CrossRef]
- Lal, R. Restoring soil quality to mitigate soil degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef] [Green Version]
- Kassam, A.; Friedrich, T.; Derpsch, R.; Lahmar, R.; Mrabet, R.; Basch, G.; González-Sánchez, E.J.; Serraj, R. Conservation agriculture in the dry Mediterranean climate. Field Crop. Res. 2012, 132, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Carbonell-Bojollo, R.; Ordóñez-Fernández, R.; Rodríguez-Lizana, A. Influence of olive mill waste application on the role of soil as a carbon source or sink. Clim. Chang. 2010, 102, 625–640. [Google Scholar] [CrossRef]
- Rodríguez-Lizana, A.; Ordóñez, R.; Espejo-Pérez, A.J.; González, P. Plant cover and control of diffuse pollution from P in olive groves. Water Air Soil Pollut. 2007, 181, 17–34. [Google Scholar] [CrossRef]
- Durán-Zuazo, V.H.; Rodríguez-Pleguezuelo, C.R. Soil-erosion and runoff prevention by plant covers. A review. Agron. Sustain. Dev. 2008, 28, 65–86. [Google Scholar] [CrossRef] [Green Version]
- Keesstra, S.D.; Rodrigo-Comino, J.; Novara, A.; Giménez-Morera, A.; Pulido, M.; Di Prima, S.; Cerdà, A. Straw mulch as a sustainable solution to decrease runoff and erosion in glyphosate-treated clementine plantations in Eastern Spain. An assessment using rainfall simulation experiments. Catena 2019, 174, 95–103. [Google Scholar] [CrossRef]
- Saavedra, M.; Pastor, M. Sistemas de Cultivo en Olivar: Manejo de Malas Hierbas y Herbicidas; Editorial Agrícola Española: Madrid, Spain, 2002. [Google Scholar]
- MAGRAMA. Encuesta Sobre Superficies Y Rendimientos de Cultivo (ESYRCE): Análisis de las Técnicas de Mantenimiento de Los Suelos Y de Los Métodos de Siembra en España. 2019. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/cubiertas2019_tcm30-526244.pdf (accessed on 28 May 2020).
- Repullo, M.A.; Carbonell, R.; Hidalgo, J.; Rodríguez-Lizana, A.; Ordóñez, R. Using olive pruning residues to cover soil and improve fertility. Soil Tillage Res. 2012, 124, 36–46. [Google Scholar] [CrossRef]
- Rodríguez-Lizana, A.; Pereira, M.J.; Ribeiro, M.C.; Soares, A.; Márquez-García, F.; Ramos, A.; Gil-Ribes, J. Assessing local uncertainty of soil protection in an olive grove area with pruning residues cover: A geostatistical cosimulation approach. Land Degrad. Dev. 2017, 28, 2086–2097. [Google Scholar] [CrossRef]
- Ordóñez-Fernández, R.; Repullo-Ruibérriz de Torres, M.A.; Román-Vázquez, J.; González-Fernández, P.; Carbonell-Bojollo, R. Macronutrients released during the decomposition of pruning residues used as plant cover and their effect on soil fertility. J. Agric. Sci. 2015, 153, 615–630. [Google Scholar] [CrossRef]
- Rodríguez-Lizana, A.; Espejo-Pérez, A.J.; González-Fernández, P.; Ordóñez-Fernández, R. Pruning residues as an alternative to traditional tillage to reduce erosion and pollutant dispersion in olive groves. Water Air Soil Pollut. 2008, 193, 165–173. [Google Scholar] [CrossRef]
- Tribouillois, H.; Cohan, J.P.; Justes, E. Cover crop mixtures including legume produce ecosystem services of nitrate capture and green manuring: Assessment combining experimentation and modelling. Plant Soil 2016, 401, 347–364. [Google Scholar] [CrossRef]
- Alcántara, C.; Sánchez, S.; Pujadas, A.; Saavedra, M. Brassica species as winter cover crops in sustainable agricultural systems in southern spain. J. Sustain. Agric. 2009, 33, 619–635. [Google Scholar] [CrossRef]
- García-Díaz, A.; Bienes, R.; Sastre, B.; Novara, A.; Gristina, L.; Cerdà, A. Nitrogen losses in vineyards under different types of soil groundcover. A field runoff simulator approach in central Spain. Agric. Ecosyst. Environ. 2017, 236, 256–267. [Google Scholar] [CrossRef]
- Alcántara, C.; Pujadas, A.; Saavedra, M. Management of Sinapis alba subsp. mairei winter cover crop residues for summer weed control in southern Spain. Crop Prot. 2011, 30, 1239–1244. [Google Scholar] [CrossRef]
- Jurado-Bello, J.; De Haro, A.; Hidalgo, J.; Hidalgo, J.C.; Vega, V.; Bejarano-Alcázar, J. Evaluación de la eficacia de enmiendas basadas en especies crucíferas implantadas como cubiertas vegetales para el control de la verticilosis del olivo en condiciones de campo. In Proceedings of the XVI Scientific-Technical Symposium of Olive Oil Expoliva, Jaén, Spain, 8–11 May 2013. [Google Scholar]
- De Torres, M.A.R.R.; Ordóñez-Fernández, R.; Giráldez, J.V.; Márquez-García, J.; Laguna, A.; Carbonell-Bojollo, R. Efficiency of four different seeded plants and native vegetation as cover crops in the control of soil and carbon losses by water erosion in olive orchards. Land Degrad. Dev. 2018, 29, 2278–2290. [Google Scholar] [CrossRef]
- Reicosky, D.C.; Archer, D.W. Moldboard plow tillage depth and short-term carbon dioxide release. Soil Tillage Res. 2007, 94, 109–121. [Google Scholar] [CrossRef]
- Dong, S.; Ma, Z.; Wang, L.; Yan, C.; Liu, L.; Gong, Z.; Cui, G. Decomposition and nutrient release characteristics of incorporated soybean and maize straw in northeast China. Ekoloji 2019, 28, 2119–2129. [Google Scholar]
- Wang, Y.; Adnan, A.; Wang, X.; Shi, Y.; Yang, S.; Ding, Q.; Sun, G. Nutrient recycling, wheat straw decomposition, and the potential effect of straw shear strength on soil mechanical properties. Agronomy 2020, 10, 314. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Garcia, J.; Gabriel, J.L.; Alonso-Ayuso, M.; Quemada, M. Quantitative characterization of five cover crop species. J. Agric. Sci. 2015, 153, 1174–1185. [Google Scholar] [CrossRef] [Green Version]
- Weinert, T.L.; Pan, W.L.; Moneymaker, M.R.; Santo, G.S.; Stevens, R.G. Nitrogen recycling by nonleguminous winter cover crops to reduce leaching in potato rotations. Agron. J. 2002, 94, 365–372. [Google Scholar] [CrossRef]
- Gómez-Muñoz, B.; Hatch, D.J.; Bol, R.; García-Ruiz, R. Nutrient dynamics during decomposition of the residues from a sown legume or ruderal plant cover in an olive oil orchard. Agric. Ecosyst. Environ. 2014, 184, 115–123. [Google Scholar] [CrossRef]
- Ruffo, M.L.; Bollero, G.A. Modeling rye and hairy vetch residue decomposition as a function of degree-days and decomposition-days. Agron. J. 2003, 95, 900–907. [Google Scholar] [CrossRef]
- Tian, G.; Kang, B.; Brussaard, L. Biological effect of plant residues with contrasting chemical compositions under humid tropical conditions-decomposition and nutrient release. Soil Biol. Biochem. 1992, 24, 1051–1060. [Google Scholar] [CrossRef]
- Ágoston-Szabó, E.; Schöll, K.; Kiss, A.; Dinka, M. Mesh size and site effects on leaf litter decomposition in a side arm of the River Danube on the Gemenc floodplain (Danube-Dráva National Park, Hungary). Hydrobiologia 2016, 774, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Bokhorst, S.; Wardle, D.A. Microclimate within litter bags of different mesh size: Implications for the “arthropod effect” on litter decomposition. Soil Biol. Biochem. 2013, 58, 147–152. [Google Scholar] [CrossRef]
- Piccini, C.; Marchetti, A.; Francaviglia, R. Estimation of soil organic matter by geostatistical methods: Use of auxiliary information in agricultural and environmental assessment. Ecol. Indic. 2014, 36, 301–314. [Google Scholar] [CrossRef]
- Wadoux, A.M.J.C.; Marchant, B.P.; Lark, R.M. Efficient sampling for geostatistical surveys. Eur. J. Soil Sci. 2019, 70, 975–989. [Google Scholar] [CrossRef]
- Douglas, C.; Rickman, R. Estimating crop residue decomposition from air temperature, initial nitrogen content, and residue placement. Soil Sci. Soc. Am. J. 1992, 56, 272–278. [Google Scholar] [CrossRef]
- Rickman, R.; Douglas, C.; Albrecht, S.; Berc, J. Tillage, crop rotation, and organic amendment effect on changes in soil organic matter. Environ. Pollut. 2002, 116, 405–411. [Google Scholar] [CrossRef]
- Liang, Y.; Gollany, H.T.; Rickman, R.W.; Albrecht, S.L.; Follett, R.F.; Wilhelm, W.W.; Novak, J.M.; Douglas, C.L. Simulating soil organic matter with CQESTR (v. 2.0): Model description and validation against long-term experiments across North America. Ecol. Modell. 2009, 220, 568–581. [Google Scholar] [CrossRef]
- Rodríguez-Lizana, A.; De Torres, M.A.R.R.; Carbonell-Bojollo, R.; Alcántara, C.; Ordóñez-Fernández, R. Brachypodium distachyon, Sinapis alba, and controlled spontaneous vegetation as groundcovers: Soil protection and modeling decomposition. Agric. Ecosyst. Environ. 2018, 265, 62–72. [Google Scholar] [CrossRef]
- Rodríguez-Lizana, A.; Carbonell, R.; González, P.; Ordóñez, R. N, P and K released by the field decomposition of residues of a pea-wheat-sunflower rotation. Nutr. Cycl. Agroecosystems 2010, 87, 199–208. [Google Scholar] [CrossRef]
- Laloy, E.; Bielders, C.L. Modelling intercrop management impact on runoff and erosion in a continuous maize cropping system: Part I. model description, global sensitivity analysis and bayesian estimation of parameter identifiability. Eur. J. Soil Sci. 2009, 60, 1005–1021. [Google Scholar] [CrossRef]
- Gollany, H.T.; Nash, P.R.; Johnson, J.M.F.; Barbour, N.W. Predicted annual biomass input to maintain soil organic carbon under contrasting management. Agron. J. 2019, 111, 11. [Google Scholar] [CrossRef]
- Gollany, H.T.; Elnaggar, A.A. Simulating soil organic carbon changes across toposequences under dryland agriculture using CQESTR. Ecol. Modell. 2017, 355, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Quebrajo, L.; Pérez-Ruiz, M.; Rodríguez-Lizana, A.; Agüera, J. An approach to precise nitrogen management using hand-held crop sensor measurements and winter wheat yield mapping in a mediterranean environment. Sensors 2015, 15, 5504–5517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: WAshington, DC, USA, 2014. [Google Scholar]
- Chochois, V.; Voge, J.P.; Rebetzke, G.J.; Watt, M. Variation in adult plant phenotypes and partitioning among seed and stem-borne roots across Brachypodium distachyon accessions to exploit in breeding cereals for well-watered and drought environments. Plant Physiol. 2015, 168, 953–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajzler, M.; Klimesová, J.; Streda, T.; Vejrazka, K.; Marcek, V.; Cholastová, T. Root system production and aboveground biomass production of chosen cover crops. Eng. Technol. 2012, 69, 713–718. [Google Scholar]
- Lohmann, M.; Scheu, S.; Müller, C. Decomposers and root feeders interactively affect plant defence in Sinapis alba. Oecologia 2009, 160, 289–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Peña, M.; González-Moro, M.B.; Marino, D. Providing carbon skeletons to sustain amide synthesis in roots underlines the suitability of Brachypodium distachyon for the study of ammonium stress in cereals. AoB Plants 2019, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, K.; Iglewicz, B. Bivariate extensions of the boxplot. Technometrics 1992, 34, 307–320. [Google Scholar] [CrossRef]
- Wilcox, R. Introduction to Robust Estimation and Hypothesis Testing, 2nd ed.; Elsevier Academic Press: Burlington, CO, USA, 2010; ISBN 0-12-751542-9. [Google Scholar]
- Giloni, A.; Padberg, M. Least trimmed squares regression, least median squares regression, and mathematical programming. Math. Comput. Model. 2002, 35, 1043–1060. [Google Scholar] [CrossRef]
- Klemes, V. Operational testing of hydrological simulation models. Hydrol. Sci. J. 1986, 31, 13–24. [Google Scholar] [CrossRef]
- Zema, D.A.; Bingner, R.; Denisi, P.; Govers, G.; Licciardello, F.; Zimbone, S.M. Evaluation of runoff, peak flow and sediment yield events simulated by annagnps in a belgian agricultural watershed. Land Degrad. Dev. 2012, 23, 205–215. [Google Scholar] [CrossRef]
- Nash, J.; Sutcliffe, J. River flow forecasting through conceptual models: Part I. A discussion of principles. J. Hidrol. 1970, 10, 282–290. [Google Scholar] [CrossRef]
- Loague, K.; Green, R.E. Statistical and graphical methods for evaluating solute transport models: Overview and application. J. Contam. Hidrol. 1991, 7, 51–73. [Google Scholar] [CrossRef]
- Van Liew, M.W.; Garbretch, J. Hydrologic simulation of the Little Washita River experimental watersed using SWAT. J. Am. Water Resour. Assoc. 2003, 39, 413–426. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org (accessed on 17 July 2020).
- Ma, S.; He, F.; Tian, D.; Zou, D.; Yan, Z.; Yang, Y.; Zhou, T.; Huang, K.; Shen, H.; Fang, J. Variations and determinants of carbon content in plants: A global synthesis. Biogeosciences 2018, 15, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, L.G.; Cherubin, M.R.; Oliveira, D.M.S.; Cerri, C.E.P.; Cerri, C.C. Decomposition of sugarcane straw: Basis for management decisions for bioenergy production. Biomass Bioenergy 2019, 122, 133–144. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, X.; Wei, T.; Yang, Z.; Jia, Z.; Yang, B.; Han, Q.; Ren, X. Effects of straw incorporation on the soil nutrient contents, enzyme activities, and crop yield in a semiarid region of china. Soil Tillage Res. 2016, 160, 65–72. [Google Scholar] [CrossRef]
- Sui, N.; Zhou, Z.; Yu, C.; Liu, R.; Yang, C.; Zhang, F.; Song, G.; Meng, Y. Yield and potassium use efficiency of cotton with wheat straw incorporation and potassium fertilization on soils with various conditions in the wheat- cotton rotation system. Field Crop. Res. 2015, 172, 132–144. [Google Scholar] [CrossRef]
- Aznar-Sánchez, J.A.; Velasco-Muñoz, J.F.; López-Felices, B.; Del Moral-Torres, F. Barriers and Facilitators for Adopting Sustainable Soil Management Practices in Mediterranean Olive Groves. Agronomy 2020, 10, 506. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lu, J.; Li, X.; Ren, T.; Cong, R.; Zhou, L. Dynamics of potassium release and adsorption on rice straw residue. PLoS ONE 2014, 9, e90440. [Google Scholar] [CrossRef]
- Ordóñez-Fernández, R.; De Torres, M.A.R.R.; Márquez-García, J.; Moreno-García, M.; Carbonell-Bojollo, R.M. Legumes used as cover crops to reduce fertilisation problems improving soil nitrate in an organic orchard. Eur. J. Agron. 2018, 95, 1–13. [Google Scholar] [CrossRef]
- Wang, X.; Yang, H.; Liu, J.; Wu, J.; Chen, W.; Wu, J.; Zhu, L.; Bian, X. Effects of ditch-buried straw return on soil organic carbon and rice yields in a rice-wheat rotation system. Catena 2015, 127, 56–63. [Google Scholar] [CrossRef]
- Goovaerts, P. Geostatistical modelling of uncertainty in soil science. Geoderma 2001, 103, 3–26. [Google Scholar] [CrossRef]
- Keskin, H.; Grunwald, S. Regression kriging as a workhorse in the digital soil mapper’s toolbox. Geoderma 2018, 326, 22–41. [Google Scholar] [CrossRef]
- Cavigelli, M.A.; Nash, P.R.; Gollany, H.T.; Rasmann, C.; Polumsky, R.W.; Le, A.N.; Conklin, A.E. Simulated soil organic carbon changes in maryland are affected by tillage, climate change, and crop yield. J. Environ. Qual. 2018, 47, 588–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordóñez-Fernández, R.; Rodríguez-Lizana, A.; Carbonell, R.; González, P.; Perea, F. Dynamics of residue decomposition in the field in a dryland rotation under Mediterranean climate conditions in southern Spain. Nutr. Cycl. Agroecosystems 2007, 79, 243–253. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth (cm) | Sand (%) | Silt (%) | Clay (%) | OM (%) | CO3 = (%) | N (%) | P (mg kg−1) | K (mg kg−1) | CEC (molc kg−1) | pH (H2O) | pH (CaCl2) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–10 | 6.0 | 43.5 | 50.5 | 0.85 | 29.9 | 0.04 | 6.5 | 326.2 | 0.24 | 8.1 | 7.7 |
| 10–20 | 9.8 | 39.4 | 50.8 | 0.72 | 28.5 | 0.03 | 13.6 | 369.5 | 0.22 | 8.2 | 7.7 |
| 20–40 | 8.4 | 41.7 | 49.9 | 0.65 | 31.8 | 0.02 | 9.9 | 271.8 | 0.23 | 8.3 | 7.7 |
| 40–60 | 8.8 | 41.8 | 49.4 | 0.58 | 33.1 | 0.02 | 11.0 | 209.7 | 0.22 | 8.4 | 7.7 |
| Year | Date | Operations | ||
|---|---|---|---|---|
| Brachypodium | Sinapis | Spontaneous Soil Cover | ||
| 1 | 22/10/07 | Disc harrow Sowing Drag tine harrow | Disc harrow Sowing Drag tine harrow | Disc harrow |
| 31/03/08 | Mowing | Mowing | Mowing | |
| 07/05/08 | Mowing | Mowing | Mowing | |
| 2 | 25/11/08 | † | Disc harrow Sowing Drag tine harrow | † |
| 03/04/09 | Mowing | Mowing | Mowing | |
| 07/05/09 | Mowing | Mowing | Mowing | |
| 3 | 30/11/09 | † | Disc harrow Sowing Drag tine harrow | † |
| 24/03/10 | Mowing | Mowing | Mowing | |
| 03/05/10 | Mowing | Mowing | Mowing | |
| 4 | 04/11/10 | † | Disc harrow Sowing Drag tine harrow | † |
| 22/03/11 | Mowing | Mowing | Mowing | |
| 10/05/11 | Mowing | Mowing | Mowing | |
| Species | Year | Dates | ||||||
|---|---|---|---|---|---|---|---|---|
| Brachypodium | 1 | 13/05/08 | 03/06/08 | 27/06/08 | 11/07/08 | 28/08/08 | 25/09/08 | 16/10/08 |
| 2 | 11/05/09 | 09/06/09 | 25/06/09 | 16/07/09 | 16/09/09 | 29/10/09 | ||
| 3 | 10/05/10 | 15/06/10 | 30/07/10 | 22/09/10 | 19/10/10 | |||
| 4 | 15/06/11 | 22/08/11 | 13/09/11 | |||||
| Sinapis | 1 | 03/06/08 | 27/06/08 | 11/07/08 | 28/08/08 | 25/09/08 | 16/10/08 | |
| 2 | 25/06/09 | 16/07/09 | 27/08/09 | 16/09/09 | 29/10/09 | |||
| 3 | 10/05/10 | 15/06/10 | 30/07/10 | 24/08/10 | 22/09/10 | 19/10/10 | ||
| 4 | 15/06/11 | 22/08/11 | 13/09/11 | |||||
| Spontaneous | 1 | 11/07/08 | 28/08/08 | 25/09/08 | 16/10/08 | |||
| 2 | 16/07/09 | 27/08/09 | 16/09/09 | 29/10/09 | ||||
| 3 | 10/05/10 | 15/06/10 | 30/07/10 | 22/09/10 | 19/10/10 | |||
| 4 | 15/06/11 | 22/08/11 | 13/09/11 | |||||
| Cover Crop | Element (%) | Year | ||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | Average | ||
| Brachypodium | C | 40.8 ± 1.1 | 41.7 ± 0.8 | 37.4 ± 3.0 | 39.7 ± 1.1 | 39.6 ± 0.4 |
| N | 1.9 ± 0.3 | 1.3 ± 0.1 | 1.6 ± 0.5 | 1.1 ± 0.3 | 1.5 ± 0.1 | |
| P | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.0 | 0.2 ± 0.0 | |
| K | 1.6 ± 0.5 | 1.6 ± 0.8 | 0.9 ± 0.9 | 0.6 ± 0.2 | 1.2 ± 0.1 | |
| Sinapis | C | 41.5 ± 1.2 | 41.8 ± 1.9 | 38.4 ± 1.4 | 38.5 ± 1.7 | 40.1 ± 0.3 |
| N | 1.2 ± 0.3 | 1.3 ± 0.2 | 2.2 ± 0.4 | 1.2 ± 0.2 | 1.5 ± 0.1 | |
| P | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.3 ± 0.1 | 0.2 ± 0.0 | 0.2 ± 0.0 | |
| K | 0.5 ± 0.5 | 0.6 ± 0.2 | 2.9 ± 1.0 | 0.5 ± 0.3 | 1.2 ± 0.2 | |
| Spontaneous | C | 41.3 ± 1.1 | 40.7 ± 1.4 | 38.4 ± 1.5 | 38.4 ± 1.5 | 39.9 ± 0.3 |
| N | 1.4 ± 0.4 | 1.5 ± 0.4 | 2.2 ± 0.4 | 1.2 ± 0.6 | 1.6 ± 0.1 | |
| P | 0.1 ± 0.0 | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.0 | |
| K | 1.9 ± 1.0 | 1.1 ± 0.5 | 2.7 ± 1.1 | 0.3 ± 0.1 | 1.5 ± 0.2 | |
| Cover Crop | Element † | Year | ||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | Average | ||
| Brachypodium | C (shoots) | 72.4 | 32.8 | 29.2 | 33.2 | 42.2 |
| C (roots) | 58.5 | 62.4 | 60.6 | 42.7 | 57.0 | |
| N | 87.5 | 25.3 | 28.4 | 35.2 | 47.5 | |
| P | 89.3 | 59.5 | 25.0 | 44.3 | 57.7 | |
| K | 96.9 | 70.6 | 75.8 | 73.2 | 80.8 | |
| Sinapis | C (shoots) | 67.0 | 41.4 | 48.2 | 48.7 | 48.3 |
| C (roots) | 53.6 | 51.0 | 59.2 | 41.5 | 51.0 | |
| N | 51.9 | 23.7 | 64.4 | 55.1 | 45.8 | |
| P | 51.6 | 16.4 | 62.8 | 47.9 | 42.7 | |
| K | 72.4 | 67.4 | 95.7 | 72.0 | 85.0 | |
| Spontaneous | C (shoots) | 56.5 | 26.4 | 59.7 | 33.5 | 40.5 |
| N | 70.1 | 23.0 | 71.9 | 20.7 | 46.0 | |
| P | 82.3 | 25.5 | 74.1 | 35.1 | 50.8 | |
| K | 94.6 | 83.5 | 96.7 | 56.9 | 90.4 | |
| Cover crop | Amount Released (kg ha−1) | Year | |||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | Sum † | Global †† | ||
| Brachypodium | C (shoots) | 1974 | 1444.8 | 435.9 | 747.9 | 4602 | 10886 |
| C (roots) | 2354.5 | 3923.5 | 1482.1 | 1461.9 | 9222 | 16175 | |
| N ††† | 108.2 | 35.0 | 17.2 | 21.0 | 181 | 382 | |
| P | 10.0 | 12.6 | 1.9 | 4.3 | 29 | 50 | |
| K | 100.0 | 90.4 | 16.8 | 23.7 | 231 | 286 | |
| Sinapis | C (shoots) | 1110.2 | 1875.5 | 856.2 | 964.5 | 4806 | 9945 |
| C (roots) | 285.6 | 751.4 | 371.3 | 283.9 | 1692 | 3316 | |
| N | 24.2 | 29.4 | 65.1 | 33.8 | 152 | 333 | |
| P | 2.5 | 2.4 | 8.7 | 4.8 | 18 | 43 | |
| K | 10.8 | 37.1 | 131.2 | 16.4 | 195 | 230 | |
| Spontaneous | C (shoots) | 516.2 | 834.9 | 1251.3 | 512.8 | 3115 | 7694 |
| N | 19.9 | 25.2 | 83.2 | 9.5 | 138 | 300 | |
| P | 2.6 | 3.7 | 12.7 | 2.5 | 21 | 42 | |
| K | 42.8 | 73.6 | 131.5 | 7.7 | 256 | 283 | |
| Cover Crop | Parameter | |
|---|---|---|
| α | β | |
| Brachypodium | −39.02 | 0.4128 |
| Sinapis | −62.28 | 0.4238 |
| Spontaneous | 1.77 | 0.3948 |
| Species | Element | ||||
|---|---|---|---|---|---|
| C | N | P | |||
| Brachypodium distachyon | Coefficient | E | 0.69 | 0.38 | 0.28 |
| CRM | −0.02 | −0.12 | −0.14 | ||
| Observed values (kg ha−1) | Mean | 2433 | 76 | 8.8 | |
| SD | 1167 | 41 | 5.5 | ||
| Simulated values (kg ha−1) | Mean | 2493 | 85 | 9.9 | |
| SD | 898 | 31 | 4.7 | ||
| Correlation | rxy ✡ | 0.84 | 0.67 | 0.62 | |
| Sinapis alba | Coefficient | E | 0.64 | 0.54 | 0.37 |
| CRM | −0.12 | 0.08 | 0.08 | ||
| Observed values (kg ha−1) | Mean | 1815 | 60 | 7.2 | |
| SD | 1155 | 40 | 5.1 | ||
| Simulated values (kg ha−1) | Mean | 2039 | 55 | 6.7 | |
| SD | 1111 | 29 | 3.7 | ||
| Correlation | rxy ✡ | 0.83 | 0.75 | 0.63 | |
| Spontaneous soil cover | Coefficient | E | 0.72 | 0.50 | 0.44 |
| CRM | −0.10 | −0.04 | −0.01 | ||
| Observed values (kg ha−1) | Mean | 1503 | 54 | 6.8 | |
| SD | 900 | 39 | 5.1 | ||
| Simulated values (kg ha−1) | Mean | 1647 | 57 | 6.7 | |
| SD | 716 | 31 | 4.6 | ||
| Correlation | rxy ✡ | 0.83 | 0.72 | 0.70 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Lizana, A.; Repullo-Ruibérriz de Torres, M.Á.; Carbonell-Bojollo, R.; Moreno-García, M.; Ordóñez-Fernández, R. Study of C, N, P and K Release from Residues of Newly Proposed Cover Crops in a Spanish Olive Grove. Agronomy 2020, 10, 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10071041
Rodríguez-Lizana A, Repullo-Ruibérriz de Torres MÁ, Carbonell-Bojollo R, Moreno-García M, Ordóñez-Fernández R. Study of C, N, P and K Release from Residues of Newly Proposed Cover Crops in a Spanish Olive Grove. Agronomy. 2020; 10(7):1041. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10071041
Chicago/Turabian StyleRodríguez-Lizana, Antonio, Miguel Ángel Repullo-Ruibérriz de Torres, Rosa Carbonell-Bojollo, Manuel Moreno-García, and Rafaela Ordóñez-Fernández. 2020. "Study of C, N, P and K Release from Residues of Newly Proposed Cover Crops in a Spanish Olive Grove" Agronomy 10, no. 7: 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10071041